The complete mitochondrial genome of Fusicolla acetilerea (Nectriaceae, Hypocreales)
Yehyeon Cha, Young-Hyun You, Seung-Yeol Lee, Hee-Young Jung, Changmu Kim, Seung-Yoon Oh

TL;DR
This paper presents the full mitochondrial genome of the fungus Fusicolla acetilerea, offering a valuable resource for genetic studies in this and related species.
Contribution
The study provides the first complete mitochondrial genome for Fusicolla acetilerea, enhancing genomic resources for this under-researched fungus.
Findings
The mitochondrial genome is 29,182 bp long with a GC content of 28.7%.
It includes all standard oxidative phosphorylation genes, two rRNA genes, 25 tRNA genes, and an intronic rps3 gene.
Phylogenetic analysis places F. acetilerea within Nectriaceae, closely related to Nectria cinnabarina.
Abstract
Fusicolla acetilerea (Nectriaceae, Ascomycota) is an emerging saprotrophic/plant-associated fungus whose genomic resources remain scarce. Here, we report the complete mitochondrial genome of F. acetilerea strain NIBRFGC000505922, assembled from Illumina NovaSeq reads. The circular-mapping mitogenome is 29,182 bp in length with a GC content of 28.7%. It encodes the canonical set of oxidative phosphorylation genes, two rRNA genes, 25 tRNA genes, and an intronic rps3 gene. A phylogenetic analysis based on 14 core protein-coding genes places F. acetilerea within Nectriaceae, forming a well-supported clade with Nectria cinnabarina. This mitogenome provides a reference for various genetic studies in Fusicolla and related taxa.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
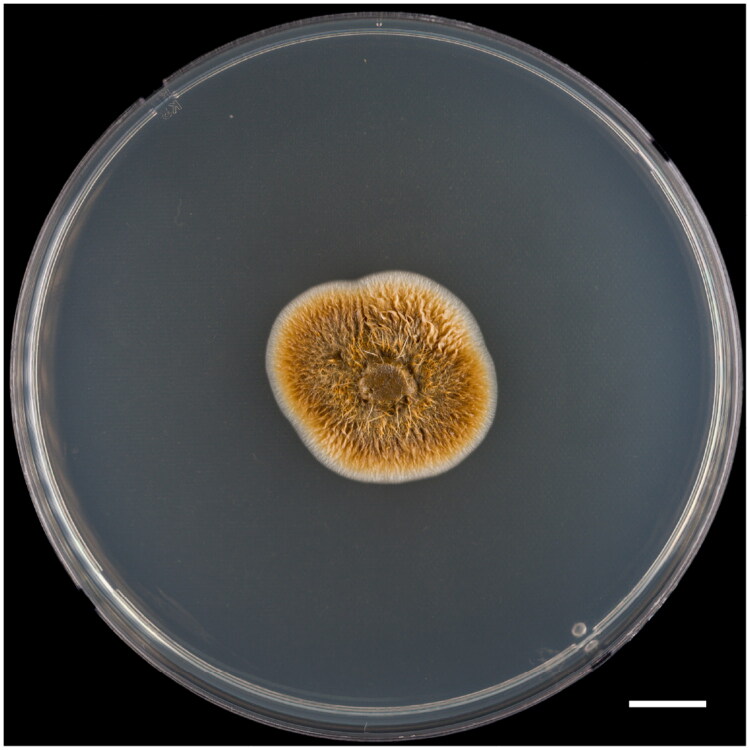 Figure 1
Figure 1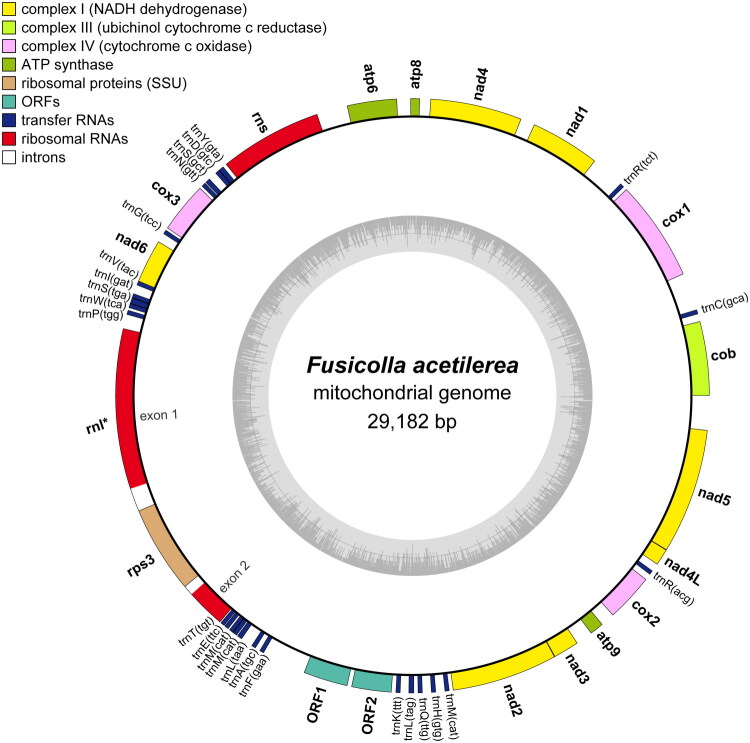 Figure 2
Figure 2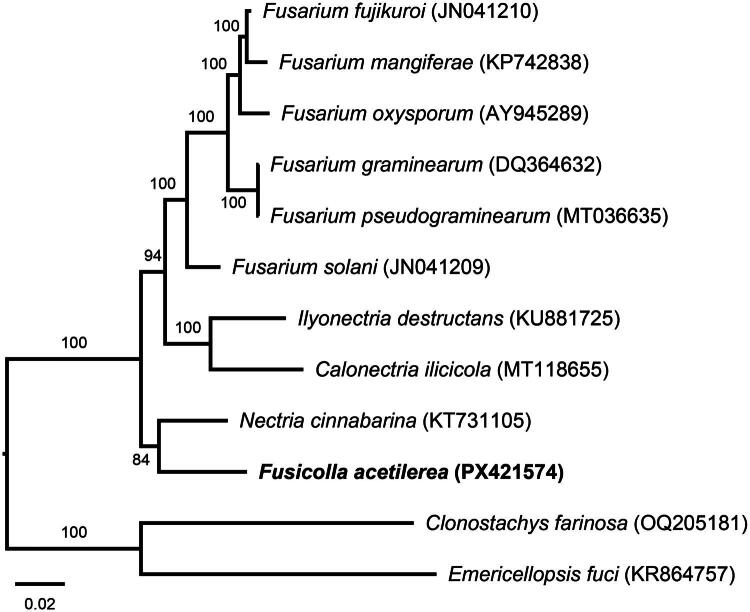 Figure 3
Figure 3- —National Institute of Biological Resources (NIBR)10.13039/501100005880
- —Ministry of Environment (MOE) of the Republic of Korea10.13039/501100003562
- —Korea Basic Science Institute (National Research Facilities and Equipment Center) Grant10.13039/501100003716
- —Ministry of Education10.13039/100009122
- —RISE (Regional Innovation System & Education) Program10.13039/100024717
- —Ministry of Education10.13039/100009122
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Genomics and Phylogenetic Studies · Enzyme-mediated dye degradation
Introduction
The genus Fusicolla Bonord. 1851 is primarily saprotrophic and has been detected on diverse organic substrates, including decaying twigs (Zeng and Zhuang 2023), soil (Liu et al. 2022), wild-boar bones (Lechat and Rossman 2017), and even water (Jeon et al. 2020). Beyond its ecological roles, multiple studies have emphasized the significance of Fusicolla species on human health (Zhong et al. 2021), fermentation (Zhu and Huang 2021), and agriculture (Wang et al. 2020). Given these contexts, research on the diversity and functional traits of Fusicolla is of both academical and practical importance.
Fusicolla acetilerea (Tubaki, C. Booth & T. Harada) Gräfenhan & Seifert 2011 (Gräfenhan et al. 2011) is notable species within the genus, characterized by large macroconidia with typically three (occasionally four) transverse septa, formed on phialidic conidiophores, with conidial dimensions of (30–)35–40 × 3.5(–4.0) µm (Tubaki et al. 1976). F. acetilerea has been reported from soil, decayed wood, and in association with ambrosia beetles (Biedermann et al. 2013; Ding et al. 2015). Notably, F. acetilerea strain NIBRFGC000505922 (=KNUF-20-PBU01), a isolate from soil in South Korea, shows the ability of polycaprolactone (PCL) and polylactic acid (PLA) degradation, which suggests potential application on plastic-waste treatment (Lee et al. 2021). In addition, this species can utilize triacrylonitrile and 4-N-trimethylamino-1-butanol as the sole carbon and nitrogen source (Asano et al. 1981; Fujimitsu et al. 2016).
While nuclear genes (acl1, ITS, LSU, rpb2, and tub2) have been actively used to ecological and phylogenetic studies (Lee et al. 2021; Zeng and Zhuang 2023), none of studies use mitochondrial genome for Fusicolla species. In this study, we assembled the mitochondrial genome of F. acetilerea strain NIBRFGC000505922 for the first time in this genus and analyzed its phylogenetic relationships with other Nectriaceae species.
Materials and methods
The F. acetilerea strain NIBRFGC000505922 was isolated from the rhizosphere soil associated with pine tree in Boeun, South Korea (36.488767 N, 127.7174 E) and cultured on potato dextrose agar (PDA) at 25 °C (Figure 1). The strain was identified in the previous study (Lee et al. 2021) and is deposited at the National Institute of Biological Resources (NIBR), Incheon, South Korea (https://species.nibr.go.kr/nibrbiobank, contact: [email protected]) under the strain number NIBRFGC000505922. Genomic DNA was extracted using the Wizard^®^ HMW DNA Extraction Kit (Promega, Madison, WI). DNA quality was evaluated using a Qubit 4.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA) and TapeStation D1000 ScreenTape system (Agilent, Santa Clara, CA). Sequencing was performed using the Illumina NovaSeqX platform at PHYZEN (Seongnam, South Korea).
Colony morphology of Fusicolla acetilerea strain NIBRFGC000505922 on potato dextrose agar. Scale bar = 1 cm. Photograph by Dr. Jung-Jae Woo.
Raw sequencing reads were quality-filtered using FASTP (Chen et al. 2018) and assembled de novo with NOVOPlasty v3.6 (Dierckxsens et al. 2017). Coverage analysis revealed an average depth of 2772× (Figure S1), following method from the previous study (Ni et al. 2023). Annotation of mitochondrial genome was performed using MFannot (Lang et al. 2023) and GeSeq (Tillich et al. 2017), with tRNA genes predicted by tRNAscan-SE v2.0 (Lowe and Chan 2016) and manually curated in Geneious Prime v.2025.2.2 (Dotmatics, Boston, MA). A circular genome map was visualized using OGDRAW v 1.3.1 (Greiner et al. 2019). For phylogenetic analysis, 14 conserved protein-coding genes (PCGs) (atp6, atp8, atp9, cob, cox1–3, nad1–6, and nad4L) were aligned using MUSCLE v.5.3 (Edgar 2022) and poorly aligned positions were removed with Gblock v. 0.91b (Castresana 2000). Then, each alignment was tested for selecting the substitution model using ModelTest-NG v0.1.7 (Darriba et al. 2020) and all alignments were concatenated to single alignment. For all markers, MTZOA was chosen as best-fit model, except for cox1 (LG), nad5 (DCMUT), and nad6 (JTT). Clonostachys farinosa (Henn.) Rossman 2014 (OQ205181) (Oh 2024) and Emericellopsis fuci (Summerb., Zuccaro & W. Gams) L.W. Hou, L. Cai & Crous 2023 (KR864757) (Konovalova and Logacheva 2016) belonged to Bionectriaceae in Hypocreales were used as outgroups. Maximum-likelihood phylogenetic analysis was conducted using RAxML v. 8.2.12 (Stamatakis 2014) with 1000 bootstrap replicates in raxmlGUI v2.0 (Edler et al. 2021).
Results
The complete mitochondrial genome of F. acetilerea was assembled into a circular molecule of 29,182 bp in length, with an overall GC content of 28.7% (Figure 2). The mitogenome encodes a total of 14 standard PCGs involved in oxidative phosphorylation, including three subunits of cytochrome c oxidase (cox1, cox2, and cox3), apocytochrome b (cob), seven subunits of NADH dehydrogenase (nad1, nad2, nad3, nad4, nad4L, nad5, and nad6), and three ATP synthase subunits (atp6, atp8, and atp9). Additionally, the genome contains two ribosomal RNA genes (rnl and rns) and 25 transfer RNA genes (tRNAs) corresponding to all 20 standard amino acids, with some tRNAs exhibiting redundancy (Arg, Leu, Met, and Ser). Intron was found in rnl region as a group IA, harboring rps3 gene. In addition, two free-standing open reading frames (ORF1 and ORF2) encoding hypothetical proteins were annotated in the mitochondrial genome. Phylogenetic analysis based on concatenated amino acid sequences of the 14 PCGs placed F. acetilerea within the family Nectriaceae, forming a sister relationship with Nectria cinnabarina (Tode) Fr. 1849, with strong bootstrap support (84%) (Figure 3). The mitogenome sequence has been deposited in GenBank under accession number PX421574.
Mitogenome feature maps of Fusicolla acetilerea strain NIBRFGC000505922. Genes are depicted on the outer circle and color-coded according to functional categories. GC content is shown in the inner circle. Genes containing introns are indicated with an asterisk ().*
Maximum-likelihood (ML) phylogenetic tree of nine Nectriaceae species inferred from concatenated amino acid sequences of 14 conserved mitochondrial genes (atp6, atp8, atp9, cob, cox1–3, nad1–6, and nad4L). Calonectria ilicicola (MT118655.1; Gai et al. 2020), Clonostachys farinosa (OQ205181.2; Oh 2024), Emericellopsis fuci (KR864757.1; Konovalova and Logacheva 2016), Fusarium fujikuroi (JN041210.1; Al-Reedy et al. 2012), Fusarium graminearum (DQ364632.1; Al-Reedy et al. 2012), Fusarium mangiferae (KP742838.1; unpublished), Fusarium oxysporum (AY945289.1; Pantou et al. 2008), Fusarium pseudograminearum (MT036635.1; Kulik et al. 2020), Fusarium solani (JN041209.1; Al-Reedy et al. 2012), Ilyonectria destructans (KU881725.1; Okorski and Majchrzak 2007), and Nectria cinnabarina (KT731105.1; Wang et al. 2016) were included as a reference. Bootstrap support values (1000 replicates) are indicated on the branch. The scale bar shows the substitution number per site.
Discussion and conclusions
The mitogenome of F. acetilerea obtained from this study is the first complete mitochondrial genome characterized for the genus Fusicolla, marking a significant contribution in the genomic study of Nectriaceae fungi. The conserved gene content, comprising 14 PCGs, two rRNA genes, and 25 tRNA genes, align closely with those reported for related Nectriaceae species (Al-Reedy et al. 2012; Gai et al. 2020). In addition, the presence of a group IA intron in the rnl region encoding the rps3 gene is similar to patterns observed in other Nectriaceae mitogenomes, consistent with ribosomal protein synthesis regulation in fungal mitochondria (Kouvelis and Hausner 2022). A distinctive feature of the F. acetilerea is its compact size of mitogenome (29,182 bp), making it the smallest known mitogenome within the Nectriaceae family (30,629–110,525 bp) (Kulik et al. 2020). The compact mitogenome of F. acetilerea may reflect reduced intergenic regions or fewer intronic sequences, though the specific evolutionary factors contributing to this compactness remain unclear. Phylogenetic analysis of mitogenome positioned F. acetilerea as a sister to N. cinnabarina within Nectriaceae, aligning with recent taxonomic revisions in the family (Lombard et al. 2015). This relationship highlights the utility of mitogenome as robust phylogenetic markers for resolving evolutionary relationships within Nectriaceae.
The significance of this study is amplified by the limited availability of mitogenomic data within Nectriaceae. Although multiple Fusarium species have sequenced mitogenomes, most other genera remain unrepresented, except for Calonectria and Nectria with a single species of mitogenome (Wang et al. 2016; Gai et al. 2020). This scarcity hinders comparative analyses and evolutionary studies. As the first Fusicolla mitogenome, this sequence provides a key reference for studying mitochondrial evolution and intron dynamics within Nectriaceae.
Supplementary Material
Supplemental Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al-Reedy RM, Malireddy R, Dillman CB, Kennell JC. 2012. Comparative analysis of Fusarium mitochondrial genomes reveals a highly variable region that encodes an exceptionally large open reading frame. Fungal Genet Biol. 49(1):2–14. 10.1016/j.fgb.2011.11.00822178648 · doi ↗ · pubmed ↗
- 2Asano Y, Ando S, Tani Y, Yamada H, Ueno T. 1981. Fungal degradation of triacrylonitrile. Agric Biol Chem. 45(1):57–62. 10.1080/00021369.1981.10864495 · doi ↗
- 3Biedermann PH, Klepzig KD, Taborsky M, Six DL. 2013. Abundance and dynamics of filamentous fungi in the complex ambrosia gardens of the primitively eusocial beetle Xyleborinus saxesenii Ratzeburg (Coleoptera: Curculionidae, Scolytinae). FEMS Microbiol Ecol. 83(3):711–723. 10.1111/1574-6941.1202623057948 · doi ↗ · pubmed ↗
- 4Castresana J. 2000. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol. 17(4):540–552. 10.1093/oxfordjournals.molbev.a 02633410742046 · doi ↗ · pubmed ↗
- 5Chen S, Zhou Y, Chen Y, Gu J. 2018. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 34(17):i 884–i 890. 10.1093/bioinformatics/bty 56030423086 PMC 6129281 · doi ↗ · pubmed ↗
- 6Darriba D et al. 2020. Model Test-NG: a new and scalable tool for the selection of DNA and protein evolutionary models. Mol Biol Evol. 37(1):291–294. 10.1093/molbev/msz 18931432070 PMC 6984357 · doi ↗ · pubmed ↗
- 7Dierckxsens N, Mardulyn P, Smits G. 2017. NOVO Plasty: de novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 45(4):e 18. 10.1093/nar/gkw 95528204566 PMC 5389512 · doi ↗ · pubmed ↗
- 8Ding S, Hu H, Gu JD. 2015. Fungi colonizing wood sticks of Chinese fir incubated in subtropical urban soil growing with Ficus microcarpa trees. Int J Environ Sci Technol. 12(12):3781–3790. 10.1007/s 13762-015-0802-5 · doi ↗