Metabolic potentiation of antibiotic killing by L-arginine in drug-resistant Edwardsiella tarda
Bei-bei Yan, Na Li, Yang Zhou, Li-li Kang, Xue-sa Dong, Xiao Xu, Li An, Qing-lei Meng, Xi-rong Wang, Ling Yang, Xiao-ying Li, Chao Wang

TL;DR
L-arginine significantly boosts the effectiveness of chloramphenicol against drug-resistant Edwardsiella tarda by disrupting key metabolic pathways.
Contribution
This study is the first to uncover how L-arginine potentiates antibiotics via dual metabolic disruption in drug-resistant bacteria.
Findings
L-arginine enhances chloramphenicol's bactericidal activity by over 100-fold against Edwardsiella tarda.
L-arginine disrupts TCA cycle, redox balance, and phenylalanine metabolism to impair bacterial resistance mechanisms.
Abstract
The role of metabolic state reprogramming in modulating antibiotic susceptibility has attracted growing interest as a promising strategy to combat antimicrobial resistance. Our study revealed that L-arginine potentiates chloramphenicol’s bactericidal activity by at least two orders of magnitude against multidrug-resistant Edwardsiella tarda via the coordinated modulation of three interconnected metabolic pathways: the tricarboxylic acid cycle disruption, redox homeostasis alteration, and phenylalanine metabolic suppression. Mechanistically, L-arginine-mediated tricarboxylic acid cycle inhibition diminished NADH production and compromised proton motive force, thereby depleting cellular energy supply and impairing drug efflux capacity. Concurrently, L-arginine disturbed the bacterial redox balance, which normally provides antibiotic resistance, by both lowering total antioxidant capacity…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
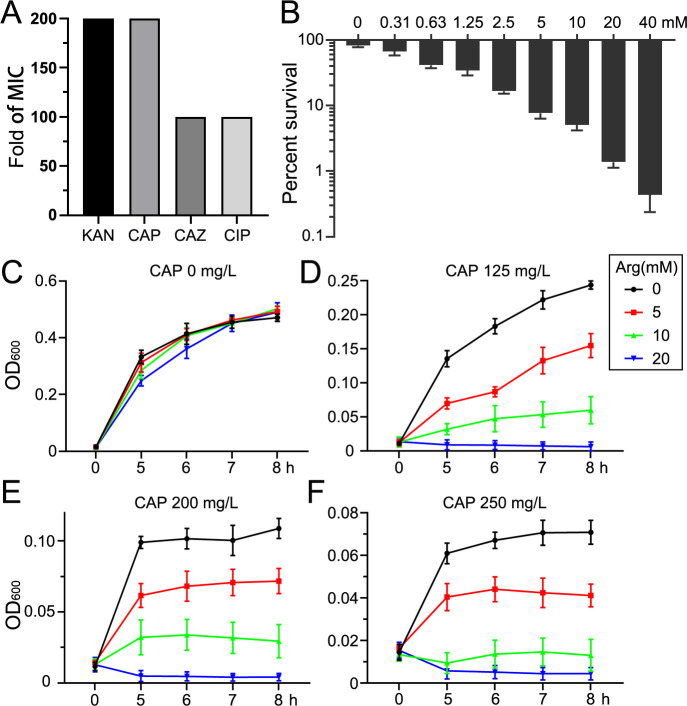 Fig 1
Fig 1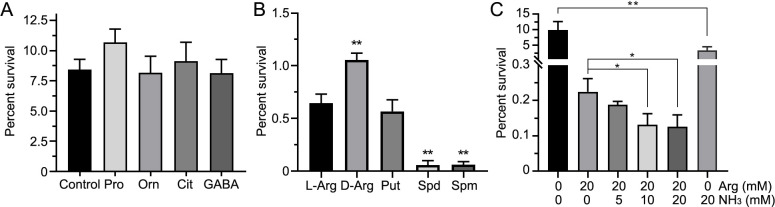 Fig 2
Fig 2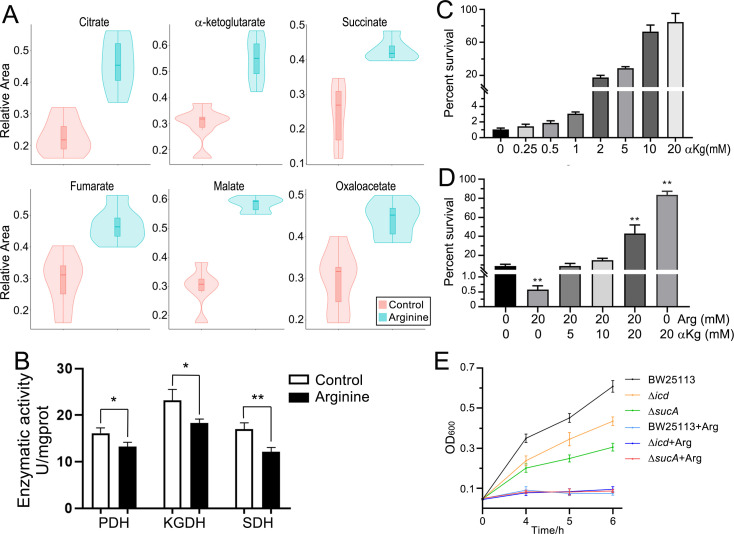 Fig 3
Fig 3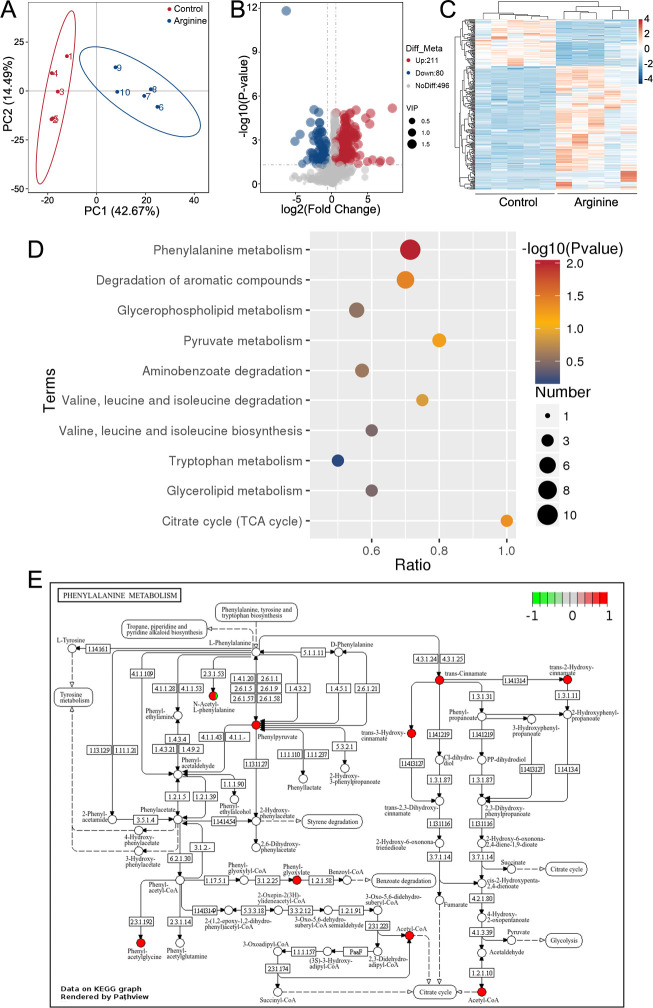 Fig 4
Fig 4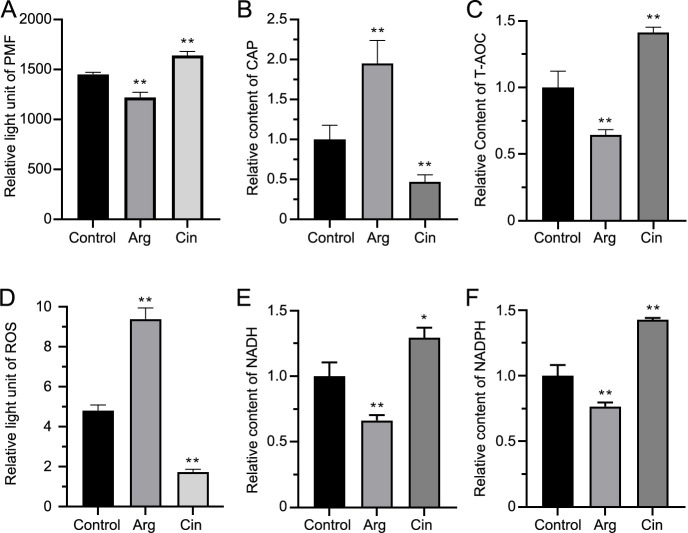 Fig 5
Fig 5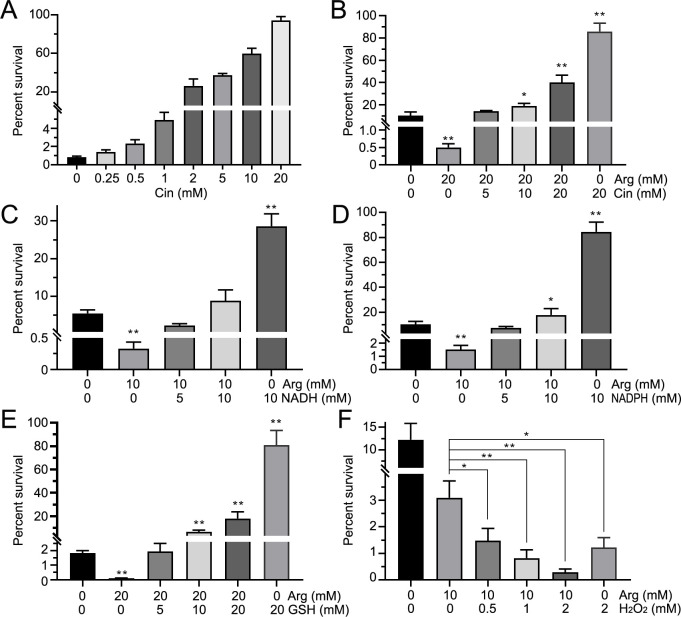 Fig 6
Fig 6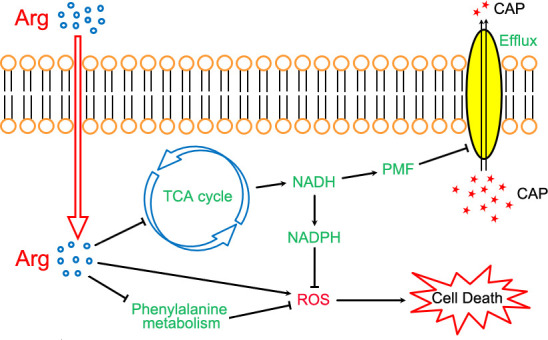 Fig 7
Fig 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial agents and applications · Cancer Research and Treatments · Biopolymer Synthesis and Applications
INTRODUCTION
Edwardsella tarda is an intracellular Gram-negative bacterium with a broad host range, capable of infecting various organisms, including humans, birds, and fish. Such infections result in substantial economic losses globally (1, 2). Antibiotics remain the cornerstone of treatment for bacterial infections. However, the overuse and misuse of antibiotics have led to a worldwide surge in antimicrobial resistance (3). Multidrug-resistant bacteria present a significant challenge, severely compromising the efficacy of infection prevention and treatment. In 2019, bacterial antimicrobial resistance was associated with approximately 5 million deaths (4), a number projected to rise dramatically by 2050 without effective interventions (5). Bacterial antimicrobial resistance, recognized as a critical global health threat, requires a comprehensive understanding of the molecular mechanisms underlying resistance to develop effective strategies against antibiotic-resistant pathogens.
In addition to traditional antibiotic resistance mechanisms, such as reduced membrane permeability, increased efflux pump activity, enzymatic degradation, and drug target modifications (6), metabolic modulation has emerged as an additional factor in bacterial resistance (7, 8). Accumulating evidence indicates that metabolic perturbations play a critical role in antibiotic-induced bacterial death (9, 10). Notably, exogenous metabolite supplementation has emerged as a promising approach to resensitize resistant pathogens through targeted metabolic reprogramming. For instance, in multidrug-resistant Escherichia coli, exogenous glutamine enhances membrane permeability and ampicillin uptake, ultimately exceeding the capacity of efflux systems and reducing bacterial survival (8). Termed metabolic reprogramming, this strategy aims to restore the efficacy of existing antibiotics and develop next-generation therapeutics by enhancing their bactericidal effects against resistant pathogens (11–14).
The tricarboxylic acid (TCA) cycle plays a pivotal role in metabolic modulation, serving as a central hub that integrates carbohydrate, amino acid, and lipid metabolism while generating NADH to fuel ATP synthesis. On one hand, the upregulation of the TCA cycle enhances the proton motive force (PMF), facilitating the intracellular uptake of aminoglycosides and thereby improving antibiotic bactericidal efficiency (8, 15–17). On the other hand, the TCA cycle impacts redox balance through reactive oxygen species (ROS) generation from its key product, NADH, during electron transport (18). The effects of different metabolites on the bactericidal activity and mechanisms of antibiotics vary considerably. For example, fumarate enhances the bactericidal activity of aminoglycosides against antibiotic-resistant P. aeruginosa, while glycolytic metabolites facilitate the eradication of persistent E. coli and Staphylococcus aureus (16, 19). However, certain metabolites within the TCA cycle can also promote antibiotic resistance, such as conferring resistance to chloramphenicol (CAP) in E. tarda (20, 21).
Beyond their primary protein-targeting mechanisms, antibiotics like CAP, tetracyclines, and quinolones can kill bacteria by stimulating the production of ROS (22–24). The bactericidal efficacy of cefoperazone-sulbactam is further enhanced through metabolic reprogramming of the P cycle, which increases NADH availability for the electron transport chain and consequently stimulates ROS production (25). Conversely, sublethal levels of ROS may promote antibiotic resistance through mechanisms such as induction of multidrug efflux pumps or enhanced mutagenesis resulting from redirected glucose flux into the pentose phosphate pathway (12, 26). This dual role makes it essential to clarify the context-dependent functions of ROS in antibiotic efficacy. To counteract ROS-mediated damage, bacteria rely on the antioxidant glutathione (GSH) to maintain cytoplasmic redox homeostasis. Exogenous pyruvate enhances flux through glycine, serine, and threonine metabolism and cysteine and methionine metabolism by activating cystathionine γ-lyase and glutamic-oxaloacetic transaminase. This metabolic reprogramming disrupts glutathione (GSH) homeostasis, resulting in ROS accumulation that ultimately potentiates gentamicin-mediated killing. Conversely, in antibiotic-resistant strains, glycine downregulation stimulates glutathione reductase activity, facilitating GSH regeneration and subsequent ROS clearance (27, 28). These findings underscore the complexity of the TCA cycle and redox state in modulating bacterial survival and antibiotic susceptibility.
As an essential amino acid, L-arginine (Arg) plays a crucial role in numerous physiological and metabolic processes, including its influence on antibiotic resistance. Arg has been shown to enhance gentamicin-mediated killing of P. aeruginosa and S. aureus in a pH-independent manner; however, this effect is mitigated by carbonylcyanide-3-chlorophenylhydrazone (CCCP), a proton ionophore that disrupts the PMF. Conversely, Arg’s ability to potentiate gentamicin killing of E. coli is pH-dependent (29). Additionally, Arg serves as a precursor for nitric oxide (NO) synthesis, which not only modulates ROS but also diminishes the alanine-enhanced bactericidal effect of gentamicin against Vibrio alginolyticus (30). Despite these insights, the mechanisms underlying Arg’s impact on the bactericidal efficacy of antibiotics remain incompletely understood, particularly in the context of CAP and E. tarda.
In this study, we elucidated how the Arg-related pathway enhances the bactericidal activity of CAP against multidrug-resistant E. tarda. We further demonstrated that this effect is intricately linked to the TCA cycle, phenylalanine metabolism, and the regulation of ROS. These findings provide new insights into the bactericidal mechanisms of antibiotics and the regulatory pathways influencing ROS, offering a broader understanding of their role in combating antibiotic resistance.
MATERIALS AND METHODS
Bacterial strains and culture conditions
E. tarda ATCC 15947 was obtained from the China Center of Industrial Culture Collection. The wild-type strains of BW25113 and isogenic deletion mutants of isocitrate dehydrogenase (Δicd) and α-ketoglutarate dehydrogenase (ΔsucA) were obtained from the KEIO collection (31). The strain was cultured in tryptic soy broth (TSB) medium or M9 minimal medium (supplemented with 1 mM MgSO_4_ and 0.1 mM CaCl_2_) at 37°C. Exogenous metabolites and antibiotics used in this study, including proline (Pro), ornithine (Orn), citrulline (Cit), gamma-aminobutyric acid (GABA), L-arginine (Arg), D-arginine (D-Arg), putrescine (Put), spermidine (Spd), spermine (Spm), trans-cinnamate (Cin), kanamycin (KAN), chloramphenicol (CAP), ceftazidime (CAZ), and ciprofloxacin (CIP), were purchased from Sigma-Aldrich Corporation.
Antibiotic bactericidal assays
Bacterial cells were grown overnight in TSB medium at 37°C and harvested by centrifugation at 5,000 × g for 5 min. The collected cells were washed three times with sterile saline and resuspended in M9 medium to a final concentration of 10^6^ CFU/mL. Exogenous metabolites and/or CAP were added to the medium, and the cultures were incubated at 37°C for 5 h. Finally, 10 μL of the culture was serially diluted and plated onto TSA plates. The plates were incubated at 37°C for 18 h, after which bacterial colonies were counted, and the CFU/mL was determined.
Metabolomic profiling
The sample preparation and metabolomics data acquisition were conducted following the protocol described (21). In brief, bacterial cells were incubated in M9 medium with or without exogenous Arg at 37°C for 5 h. The metabolic reactions were terminated by adding cold methanol, and the cells were washed twice with cold sterile saline. The harvested cells were rapidly frozen in liquid nitrogen and stored at −80°C until analysis. Cell pellets were resuspended in 0.1 mL of prechilled methanol (−20°C) and incubated for 60 min. After extraction, samples were centrifuged at 14,000 × g for 15 min at 4°C. The resulting supernatants were concentrated to dryness using a centrifugal evaporator. Dried metabolites were reconstituted in 80% (vol/vol) methanol for subsequent LC-MS/MS analysis. Chromatographic separation was performed on a Vanquish UHPLC system (Thermo Fisher Scientific) coupled to a Q Exactive HF-X hybrid quadrupole-Orbitrap mass spectrometer (Thermo Fisher Scientific). Instrumental parameters were set according to previously established methods (21).
Metabolomic data analysis
Raw mass spectrometry data were processed using Compound Discoverer software (version 3.0, Thermo Fisher Scientific). Following background ion removal and normalization using quality control (QC) samples, the identified and quantified peak data were subjected to subsequent statistical analysis. Differential metabolites were identified based on a threshold of P-value < 0.05 and |log2 (fold change)| > 1.5. Metabolic pathway annotation and analysis were conducted using the KEGG database (http://www.genome.jp/kegg/) and Pathview software (32, 33).
Measurement of membrane potential
Membrane potential was assessed using commercial kits following the protocol described (34). Briefly, bacterial cells were resuspended in M9 medium to an optical density at 600 nm (OD₆₀₀) of 0.2 and incubated at 37°C for 4 h with or without the addition of metabolites. For staining, 198 μL of the bacterial suspension was mixed with 2 μL of 10 mM DiBAC_4_(3) and incubated in the dark for 1 h. Finally, fluorescence was measured using a fluorescence plate reader (Biotek, Synergy HT, Vermont, USA) with excitation and emission wavelengths of 490 nm and 516 nm, respectively.
Assessment of redox state
ROS levels were measured following the protocol described (35). Bacterial cells were resuspended in M9 medium to OD_600_ of 0.2 and incubated at 37°C for 4 h with or without the addition of metabolites. After incubation, cells were collected, washed with PBS, and divided into two portions. One portion was analyzed for ROS. The samples were incubated with 20 µM DCFH-DA (Sigma, United States) at 37°C for 1 h in the dark. Fluorescence was measured using a microplate reader (CLARIO Star Plus, Germany) with excitation and emission wavelengths of 485 and 515 nm, respectively. The other portion was analyzed for total antioxidant capacity (T-AOC) using a ferric-reducing antioxidant power assay kit (Solarbio Science & Technology, Beijing, China). Absorbance was measured at 593 nm using a spectrophotometer.
Measurement of enzyme activity
Bacteria were grown in M9 minimal medium supplemented with or without test metabolites for 5 h at 37°C. Cells were harvested by centrifugation (5,000 × g, 5 min, 4°C) and subjected to ultrasonic lysis with ice-bath cooling. The resulting lysates were clarified by centrifugation (12,000 × g, 10 min, 4°C) to remove cellular debris. Enzyme activities, including pyruvate dehydrogenase (PDH), α-ketoglutarate dehydrogenase (KGDH), succinate dehydrogenase (SDH), NADH, and NADPH were quantified using standardized commercial assay kits (Beyotime Biotechnology, China) according to manufacturer protocols. Enzyme activities (PDH, KGDH, SDH at 566 nm; NADH and NADPH at 450 nm) were measured spectrophotometrically using a microplate reader (Tecan Infinite E Plex). All experimental values were normalized to the untreated control group. Experiments were repeated in at least three independent biological replicates.
Chloramphenicol quantification
Bacterial cultures were grown in the presence or absence of target metabolites using the method described above. Following harvest and ultrasonic lysis, the samples were subjected to chromatographic separation using an ExionLC liquid chromatography system (AB Sciex) coupled to a QTRAP 4500 mass spectrometer (AB Sciex). Chromatographic separation was performed on an Eclipse Plus C18 column (2.1 × 150 mm, 3.5 μm) maintained at 40°C. An injection volume of 10 μL was used with a mobile phase consisting of (A) water and (B) methanol, delivered at a flow rate of 0.30 mL/min. The gradient elution program was as follows: 0–5 min, 90% A to 5% A (10% B to 95% B); 5–7 min, 5% A (95% B); 7–9 min, 5% A to 90% A (95% B to 10% B). Mass spectrometric detection was conducted using an electrospray ionization source in negative ion mode with the following parameters: ion spray voltage, −4,500 V; source temperature, 550°C; collision gas, nitrogen; curtain gas, nitrogen at 241 kPa. Relative chloramphenicol levels were determined based on the corresponding peak areas.
RESULTS
Arg enhances the bactericidal activity of CAP against multidrug-resistant E. tarda
The E. tarda strain used in this study exhibited multidrug resistance to kanamycin (KAN), CAP, ceftazidime (CAZ), and ciprofloxacin (CIP) (Fig. 1A). The survival rate of E. tarda decreased progressively with increasing concentrations of Arg. Under CAP treatment, survival dropped from 81.9% in the absence of Arg to 0.437% with 40 mM Arg, representing a 187-fold reduction compared to the control (Fig. 1B). The line chart further highlights the enhanced bacterial growth inhibition achieved with Arg under varying CAP concentrations (Fig. 1C through F). Notably, Arg alone did not exhibit any inhibitory effect on bacterial growth in TSB medium. However, the combination of Arg and CAP, particularly at an Arg concentration of 20 mM, completely suppressed bacterial growth.
L-arginine enhances the bactericidal activity of chloramphenicol against multidrug-resistant E. tarda. (A) Fold increase in the MIC of multidrug-resistant E. tarda compared to the sensitive strain. (B) Effect of varying concentrations of Arg on bacterial survival. The y-axis represents the survival percentage of E. tarda with or without Arg under chloramphenicol pressure, while the x-axis indicates the concentration of Arg in M9 medium. The concentration of CAP is 0.8 mg/mL. (C–F) Impact of different concentrations of Arg and CAP on the growth of E. tarda in TSB medium. KAN, Kanamycin; CAP, Chloramphenicol; CAZ, Ceftazidime; CIP, Ciprofloxacin.
Metabolites associated with the Arg pathway increase sensitivity to CAP
The Arg-related metabolic pathway encompasses both Arg biosynthesis and catabolism. Among the tested metabolites, proline, ornithine, citrulline, and gamma-aminobutyric acid showed no significant impact on the sensitivity of E. tarda to CAP (Fig. 2A). In contrast, L-arginine, D-arginine, putrescine, spermidine, and spermine enhanced CAP’s bactericidal activity (Fig. 2B). Notably, spermidine, and spermine exhibited the most pronounced effects, increasing bactericidal efficiency by 11.3-fold and 17.7-fold, respectively, compared to Arg. Furthermore, ammonia improved sensitivity to CAP, reducing the survival rate from 9.90% to 3.32% at 20 mM (2.98-fold decrease) (Fig. 2C). However, this enhancement was less significant than that observed with Arg, which reduced survival to 0.224% at 20 mM (44.1-fold decrease). The potentiation effect of Arg on CAP activity exhibited partial dependence on environmental pH. Alkaline conditions generated by NH₃ (20 mM) showed limited but statistically significant (P < 0.05) potentiation of Arg’s antibiotic activity, further reducing survival from 0.224% (20 mM Arg) to 0.126% (20 mM Arg + 20 mM NH₃), representing a 1.79-fold increase in potency (Fig. 2C). Consequently, this pH-mediated modulation represented a secondary effect compared to Arg’s primary potentiation, as evidenced by the 44-fold efficacy enhancement with 20 mM Arg alone compared to the 2.98-fold reduction achieved by 20 mM NH₃ alone.
Metabolites related to the arginine pathway enhance sensitivity to chloramphenicol. (A–C) The y-axis represents the survival percentage of E. tarda with or without metabolites under chloramphenicol pressure. The control group contains chloramphenicol without added metabolites, while experimental groups include chloramphenicol (1.0 mg/mL) along with the indicated metabolites (10 mM). Metabolites (Pro, Orn, Cit, GABA, L-Arg, D-Arg, Put, Spd, Spm, and NH₃) are shown on the x-axis. Statistical significance is indicated (/**, P < 0.05/0.01 compared with control). Pro, proline; Orn, ornithine; Cit, citrulline; GABA, gamma-aminobutyric acid; L-Arg, L-arginine; D-Arg, D-arginine; Put, putrescine, Spd, spermidine; Spm, spermine.*
Arginine inhibits the TCA cycle
Given that TCA cycle activation has been demonstrated to enhance CAP resistance in E. tarda (20), we investigated whether Arg alters antibiotic efficacy via modulation of this metabolic pathway. Targeted LC–MS/MS analysis was performed using a triple quadrupole mass spectrometer to quantify changes in TCA cycle metabolite levels upon exogenous Arg supplementation. Untargeted profiling revealed marked increases in six key TCA cycle metabolites: citrate, α-ketoglutarate, succinate, fumarate, malate, and oxaloacetate (Fig. 3A). Enzymatic activity assays revealed that Arg suppressed the activities of PDH, KGDH, and SDH (Fig. 3B). α-Ketoglutarate (αKg), key metabolites in the TCA cycle, was found to increase the resistance of E. tarda to CAP, the percent survival from 0.834% to 94.0% (Fig. 3C). Even a small amount of αKg was sufficient to counteract the effect of Arg in enhancing the bactericidal efficiency of antibiotics. The presence of equal concentrations (20 mM) of αKg with Arg conferred CAP resistance in E. tarda. (Fig. 3D). Compared to the wild-type BW25113 strain, deletion mutants of isocitrate dehydrogenase (Δicd) and α-ketoglutarate dehydrogenase (ΔsucA) exhibited heightened susceptibility to CAP, with OD_600_ values decreasing from 0.609 (wild-type) to 0.435 (Δicd) and 0.306 (ΔsucA) after 6 h of treatment (Fig. 3E). Notably, Arg supplementation potentiated CAP’s inhibitory effects in all three strains.
The mutual influence between the TCA Cycle and Arg. (A) Exogenous addition of arginine leads to an increase in the content of metabolites in the TCA cycle. The y-axis represents the relative abundance of metabolites compared to the control group. (B) Enzymatic activity in the TCA cycle is suppressed by arginine. (C) α-Ketoglutarate reduces the sensitivity of bacteria to chloramphenicol (1.2 mg/mL). (D) α-Ketoglutarate reverses the bactericidal effect of L-arginine. (E) The influence of arginine on the antibiotic sensitivity of gene-deleted strains. The concentration of CAP in LB media is 0.2 mg/mL. (/**, P < 0.05/0.01 compared with control) αKg, α-ketoglutarate; Arg, L-arginine. PDH, pyruvate dehydrogenase; KGDH, α-ketoglutarate dehydrogenase; SDH, succinate dehydrogenase.*
Metabolic profiling following exogenous Arg treatment
The suppressed TCA cycle alone does not fully account for the bactericidal enhancement by Arg. We, therefore, employed metabolomics to investigate its impact on other metabolic pathways. The metabolites in the exogenous Arg group and the control group were analyzed using LC-MS/MS. Principal component analysis revealed clear separation between the two groups (Fig. 4A). A total of 787 metabolites were identified, among which 291 metabolites exhibited significant differences in the L-Arg group compared to the control group. Of these, 211 were upregulated, while 80 were downregulated (Fig. 4B). The heatmap showed distinct abundance patterns of metabolites between the two groups (Fig. 4C). KEGG enrichment analysis identified three altered pathways (P < 0.05): phenylalanine metabolism, degradation of aromatic compounds, and the TCA cycle (Fig. 4D). In the TCA cycle, three metabolites showed significant changes: acetyl-CoA and citrate were upregulated, while thiamin pyrophosphate was downregulated. Within the phenylalanine metabolism pathway, eight metabolites were upregulated, including phenylpyruvate, trans-cinnamate, phenylglyoxylate, phenylacetylglycine, trans-3-hydroxycinnamate, trans-2-hydroxycinnamate, N-acetyl-L-phenylalanine, and acetyl-CoA (Fig. 4E).
Metabolomic analysis of differential metabolites. (A) Principal component analysis of the control group and the exogenous L-arginine group. Each dot represents a biological replicate. (B) Volcano plot illustrating the differential metabolites between the control and exogenous arginine groups. The x-axis represents the fold change of metabolites, while the y-axis indicates statistical significance. Blue, red, and gray dots represent downregulated, upregulated, and non-significant metabolites, respectively. (C) Heatmap of altered metabolites in the presence or absence of L-arginine. Blue and red colors denote low and high metabolite abundance, respectively. (D) KEGG enrichment pathways. The P-value (−log10) of each pathway is represented by color, ranging from red (high significance) to blue (low significance). The diameter of the circles corresponds to the number of altered metabolites in each pathway. The x-axis indicates the ratio of altered metabolites in each pathway. (E) Phenylalanine metabolism pathway significantly enriched in the exogenous Arg group compared with the control group. Red dots represent metabolites with increased abundance.
Modulation of CAP antibacterial efficacy and cellular redox status by Arg and Cin
trans-Cinnamate (Cin), identified as a differential metabolite, serves as a critical intermediate in phenylalanine metabolism. Our biochemical analyses revealed opposing effects between Arg and Cin on bacterial physiology. PMF measurements demonstrated that Arg-mediated depolarization of membrane potential facilitated intracellular CAP accumulation (Fig. 5A and B). Exogenous Arg supplementation attenuated cellular antioxidant capacity, as evidenced by total antioxidant capacity (T-AOC) (Fig. 5C), along with elevated ROS accumulation (Fig. 5D) and decreased NADH/NADPH levels (Fig. 5E and F). In striking contrast, Cin exhibited diametrically opposite effects: it enhanced antioxidant enzyme activities while maintaining membrane potential, consequently limiting antibiotic uptake.
The influence of Arg and Cin on the PMF and redox state. Effects of L-arginine and trans-cinnamate on PMF (A), intracellular CAP (B), T-AOC (C), ROS (D), NADH (E), and NADPH (F). The y-axis represents the relative enzyme activity or content of PMF and intracellular chloramphenicol, under the influence of L-arginine (10 mM) and trans-cinnamate (10 mM) compared to the control group. (/**, P < 0.05/0.01 compared with control) Arg, L-arginine; Cin, trans-cinnamate; PMF, proton motive force; CAP, chloramphenicol; T-AOC, total antioxidant capacity; ROS, reactive oxygen species.*
Arginine-dependent potentiation of chloramphenicol efficacy is modulated by Cin, oxidizing agents, and reducing agents
Consistent with α-ketoglutarate, Cin was observed to enhance E. tarda resistance to CAP (Fig. 6A). At equimolar concentrations (20 mM Arg + 20 mM Cin), bacterial survival increased, indicating an attenuation of antibiotic sensitivity (Fig. 6B). At concentrations of 5 mM, NADH failed to reverse the antibiotic-potentiating effect of Arg (Fig. 6C). While NADH treatment alone increased bacterial survival from 5.45% to 28.6%, this protective effect was markedly weaker than that observed with either NADPH or GSH alone under equivalent conditions. Simultaneously, the combination of 10 mM NADPH with 10 mM Arg resulted in significantly higher survival rates compared to the control (Fig. 6D). Among the tested redox-related metabolites (NADH, NADPH, and GSH), GSH exhibited the most potent capacity to counteract Arg-mediated potentiation of CAP activity. At 5 mM, GSH completely abolished the bactericidal enhancement induced by 20 mM Arg (Fig. 6E). Conversely, H₂O₂ dramatically enhanced CAP bactericidal efficacy (Fig. 6F). This pro-oxidant effect synergized with Arg, resulting in further potentiation of antibiotic activity beyond either treatment alone.
The impact of cinnamate, oxidizing agents, and reducing agents on arginine-potentiated chloramphenicol bactericidal activity percent survival of bacteria under different concentrations of Cin and Arg (A and B). Metabolites, including NADH (C), NADPH (D), GSH (E), and H2O2 (F), affect the bactericidal effect of L-arginine. The y-axis represents the percent survival of bacteria with or without metabolites under chloramphenicol pressure. The x-axis indicates the concentrations of trans-cinnamate, L-arginine, NADH, NADPH, GSH, and H2O2 (mM). The concentration of CAP is 1.0 mg/mL. (/**, P < 0.05/0.01 compared with control). Cin, trans-cinnamate; Arg, L-arginine; GSH, glutathione.*
DISCUSSION
Research on Arg as an antibiotic adjuvant has primarily centered on aminoglycosides, with studies revealing that its ability to enhance bactericidal activity against pathogens such as P. aeruginosa and S. aureus is modulated by pH and proton PMF (29). Additionally, as a precursor for NO synthesis, Arg was reported to increase V. alginolyticus resistance to gentamicin through NO-mediated mechanisms (30). Arg also improved the bactericidal efficiency of ciprofloxacin and tobramycin against P. aeruginosa although the underlying mechanisms remain unclear (36).
In this study, we observed that Arg enhanced the bactericidal efficacy of CAP against multidrug-resistant E. tarda in both M9 minimal and TSB medium. In the pathway of Arg and proline metabolism, certain metabolites such as proline, ornithine, citrulline, and gamma-aminobutyric acid did not alter CAP’s bactericidal efficiency. In contrast, L-Arg, D-Arg, putrescine, spermidine, and spermine notably increased CAP’s bactericidal activity, with spermidine and spermine exhibiting even greater enhancement than Arg. D-Arg acts as an inhibitor of Arg decarboxylase, blocking the decarboxylation of L-Arg and thereby preventing the synthesis of downstream polyamines, including putrescine, spermidine, and spermine, in the Arg metabolic pathway. The similar bacterial survival rates observed with the exogenous addition of L-Arg or D-Arg suggested that L-Arg did not enhance the bactericidal efficacy of CAP through polyamine synthesis. Furthermore, previous studies have demonstrated that polyamines primarily influence the bactericidal activity of antibiotics through two mechanisms. First, polyamines mitigate ROS-induced damage by directly scavenging ROS, enhancing the activity of antioxidant enzymes such as superoxide dismutase and glutathione peroxidase, promoting the expression of antioxidant genes via pathways such as Nrf2, and producing antioxidants like NADPH and glutathione. These actions collectively enhance cellular antioxidant capacity (37, 38). Second, polyamines contribute to cell membrane homeostasis and protection against oxidative stress (39, 40). For instance, polyamines have been reported to enhance E. coli resistance to fluoroquinolones, aminoglycosides, and cephalosporins by scavenging ROS (41). In contrast to these findings, this study revealed that polyamines increased bacterial sensitivity to CAP. This discrepancy highlighted a distinct mechanism and supports the conclusion that L-Arg did not modulate the bactericidal activity of CAP through polyamine metabolism.
Given previous findings that pH influences antibiotic efficacy (29) and that many Arg metabolic intermediates (e.g., putrescine, spermidine, and spermine) are polyamines with alkaline properties, we investigated whether pH might play a role in Arg’s ability to enhance CAP’s bactericidal activity. Ammonia, a basic compound, was observed to enhance the bactericidal effect of CAP, reducing bacterial survival by 2.98-fold. However, this effect was markedly weaker than the 44.1-fold reduction mediated by Arg. Similarly, ornithine, another basic amino acid with a pKa of ~9.71 (slightly lower than Arg’s pKa of ~10.76), did not enhance CAP’s bactericidal efficiency. These findings indicate that while pH changes induced by basic metabolites like ammonia may contribute to CAP’s bactericidal activity to a certain extent, the effect is relatively limited and cannot fully account for Arg’s potent enhancement. This suggests that pH modulation is not the primary mechanism by which Arg promotes CAP’s bactericidal efficacy.
Non-targeted metabolomics analysis revealed significant perturbations in the TCA cycle following Arg supplementation, particularly showing marked upregulation of entry-point metabolites including acetyl-CoA and citrate. Subsequent targeted metabolite quantification confirmed the coordinated elevation of six TCA cycle intermediates. Intriguingly, we observed downregulation of thiamine pyrophosphate, an essential cofactor for both pyruvate dehydrogenase and αKg dehydrogenase complexes, suggesting functional impairment of the TCA cycle. This metabolic reprogramming appears mechanistically linked to enhanced antibiotic susceptibility, as evidenced by two key findings: (i) αKg was shown to promote CAP resistance while abolishing Arg’s bactericidal enhancement; (ii) the Δicd and ΔsucA deletion mutant (defective in isocitrate dehydrogenase or αKg dehydrogenase) exhibited increased sensitivity to CAP. Collectively, these results suggest that the TCA cycle inhibition by Arg underlies its enhancement of CAP’s bactericidal effect.
Arg was found to similarly enhance CAP sensitivity in both deletion strains (Δicd and ΔsucA) and BW25113, indicating that TCA cycle inhibition is not the sole mechanism underlying Arg’s antibiotic-potentiating effects. Furthermore, untargeted metabolomics revealed that exogenous Arg supplementation reprogrammed phenylalanine metabolism, resulting in the upregulation of all eight detected metabolites. The influence of phenylalanine metabolism on the bactericidal efficacy of antibiotics remains underexplored. Previous studies have shown that phenylalanine exerts varying effects on different antibiotics (42), and Cin has been reported to enhance the bactericidal activity of β-lactam antibiotics, although the underlying mechanisms remain unclear (43). Cin, a key metabolite in the phenylalanine metabolism pathway, demonstrated a concentration-dependent effect on bacterial resistance to CAP, with increasing concentrations of Cin leading to a gradual rise in resistance at lower concentration ranges. Additionally, the ability of Arg to enhance the bactericidal efficiency of CAP was markedly inhibited by Cin. Moreover, we discovered that Cin not only reduces antibiotic uptake into bacterial cells by enhancing the PMF but also mitigates antibiotic-induced oxidative damage by boosting the bacterial antioxidant capacity. PMF provides the energy required for drug efflux systems, which are essential for CAP resistance (44, 45). In a contrasting mechanism to Arg, exogenous Cin increases the PMF in E. tarda, thereby enhancing the energy supply for efflux systems and reducing intracellular antibiotic accumulation. Interestingly, while aminoglycosides rely on PMF to facilitate antibiotic uptake (28), this contrast underscores the unique mechanism by which Arg enhances E. tarda sensitivity to CAP. Collectively, these findings suggested that Arg enhanced the bactericidal efficacy of CAP by interfering with phenylalanine metabolism, particularly through the inhibition of Cin metabolism.
TCA cycle inhibition reduced NADH and ATP production—critical components for maintaining cellular reductants including NADPH and glutathione (42). Glutathione serves as a critical component of the bacterial antioxidant defense system (27, 46, 47). In multidrug-resistant E. tarda, pyruvate reprograms glycine, serine, and threonine metabolism, resulting in coordinated upregulation of glutathione peroxidase activity and concurrent suppression of glutathione reductase. This shift in redox enzyme activity disrupts cellular redox homeostasis, leading to accumulation of intracellular reactive oxygen species (ROS) and ultimately restoring antibiotic susceptibility (28). In our study, both Arg and Cin modulate redox homeostasis by altering levels of ROS, NADH, and NADPH. Furthermore, bactericidal assays with exogenous H₂O₂ and reducing agents confirmed that oxidizing agents enhance Arg-potentiated chloramphenicol efficacy, whereas reducing agents abolish it. These results demonstrate that Arg influences CAP bactericidal efficiency primarily through redox regulation.
Based on our findings, Arg was shown to enhance the sensitivity of E. tarda to CAP through three distinct mechanisms (Fig. 7). First, Arg inhibited the TCA cycle, reducing the production of NADH and PMF. This led to insufficient energy for antibiotic efflux and a deficiency in hydrogen donors required for reductants such as NADPH and glutathione. Second, Arg increased ROS levels while decreasing T-AOC, amplifying oxidative damage caused by CAP. Third, Arg further potentiated the effects of these two metabolic pathways by disrupting phenylalanine metabolism. Elevated levels of cellular reductants (NADH and NADPH) combined with enhanced PMF are known to decrease CAP susceptibility. However, L-arginine effectively counteracts this resistance phenotype through the suppression of the TCA cycle and phenylalanine metabolism.
Proposed mechanism of L-arginine enhancing the bactericidal efficiency of chloramphenicol against E. tarda. Red indicates increased metabolite content, while green represents decreased metabolite content or inhibited pathways. Arg, L-arginine; TCA, tricarboxylic acid; NADH, nicotinamide adenine dinucleotide; NADPH, nicotinamide adenine dinucleotide phosphate; PMF, proton motive force; CAP, chloramphenicol; PhM, phenylalanine metabolism; ROS, reactive oxygen species.
Building upon the concept of metabolome reprogramming (48), our study demonstrated that Arg sensitized multidrug-resistant E. tarda to CAP by modulating redox balance and inhibiting the TCA cycle and phenylalanine metabolism. These findings broaden the understanding of how metabolic modulation influences the bactericidal efficacy of antibiotics and provide a solid theoretical foundation for developing therapeutic strategies to combat antibiotic-resistant bacteria.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kamiyama S, Kuriyama A, Hashimoto T. 2019. Edwardsiella tarda bacteremia, Okayama, Japan, 2005-2016. Emerg Infect Dis 25:1817–1823. doi:10.3201/eid 2510.18051831539320 PMC 6759260 · doi ↗ · pubmed ↗
- 2Bakirova GH, Alharthy A, Corcione S, Aletreby WT, Mady AF, De Rosa FG, Karakitsos D. 2020. Fulminant septic shock due to Edwardsiella tarda infection associated with multiple liver abscesses: a case report and review of the literature. J Med Case Rep 14:144. doi:10.1186/s 13256-020-02469-832900379 PMC 7478901 · doi ↗ · pubmed ↗
- 3Maki G, Zervos M. 2021. Health care-acquired infections in low- and middle-income countries and the role of infection prevention and control. Infect Dis Clin North Am 35:827–839. doi:10.1016/j.idc.2021.04.01434362546 PMC 8331241 · doi ↗ · pubmed ↗
- 4Antimicrobial Resistance Collaborators. 2022. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399:629–655. doi:10.1016/S 0140-6736(21)02653-237500506 · doi ↗ · pubmed ↗
- 5Ho CS, Wong CTH, Aung TT, Lakshminarayanan R, Mehta JS, Rauz S, Mc Nally A, Kintses B, Peacock SJ, de la Fuente-Nunez C, Hancock REW, Ting DSJ. 2025. Antimicrobial resistance: a concise update. Lancet Microbe 6:100947. doi:10.1016/j.lanmic.2024.07.01039305919 · doi ↗ · pubmed ↗
- 6Walsh C. 2000. Molecular mechanisms that confer antibacterial drug resistance. Nature 406:775–781. doi:10.1038/3502121910963607 · doi ↗ · pubmed ↗
- 7Lopatkin AJ, Bening SC, Manson AL, Stokes JM, Kohanski MA, Badran AH, Earl AM, Cheney NJ, Yang JH, Collins JJ. 2021. Clinically relevant mutations in core metabolic genes confer antibiotic resistance. Science 371:eaba 0862. doi:10.1126/science.aba 086233602825 PMC 8285040 · doi ↗ · pubmed ↗
- 8Zhao X-L, Chen Z-G, Yang T-C, Jiang M, Wang J, Cheng Z-X, Yang M-J, Zhu J-X, Zhang T-T, Li H, Peng B, Peng X-X. 2021. Glutamine promotes antibiotic uptake to kill multidrug-resistant uropathogenic bacteria. Sci Transl Med 13:eabj 0716. doi:10.1126/scitranslmed.abj 071634936385 · doi ↗ · pubmed ↗