Cross-feeding interactions between Fusobacterium nucleatum and the glycan forager Segatella oris
Joshua R. Fletcher, Areej Malik, Jacob Driggers, Ryan C. Hunter

TL;DR
This study explores how two bacteria, Fusobacterium nucleatum and Segatella oris, interact in mucosal environments, influencing each other's growth and host responses.
Contribution
The paper reveals bidirectional microbial interactions and their impact on nutrient acquisition, gene expression, and epithelial responses in mucosal diseases.
Findings
Segatella oris enables Fusobacterium nucleatum to access sialic acids through glycan degradation.
Fusobacterium nucleatum suppresses polysaccharide utilization loci in Segatella oris.
Co-colonization modulates epithelial gene expression linked to inflammation and cancer.
Abstract
Fusobacterium nucleatum is a common member of the oral microbiota frequently associated with extra-oral infections and diverse polymicrobial environments, including chronic airway diseases and colorectal tumors. Yet, its interactions with co-colonizing microbiota remain poorly defined. Here, we investigate cross-feeding interspecies dynamics between F. nucleatum and Segatella oris, a glycan-foraging anaerobe enriched in the airways, oral cavity, and gastrointestinal tumors. Using broth cultures, cell-free supernatants, and co-culture on primary human airway epithelial cells, we identify microbe-microbe and microbe-host interactions that shape nutrient acquisition, biofilm formation, gene expression, and host responses. While mucin or S. oris supernatants modestly enhanced F. nucleatum growth, both conditions triggered transcriptional remodeling, including induction of the nan operon for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
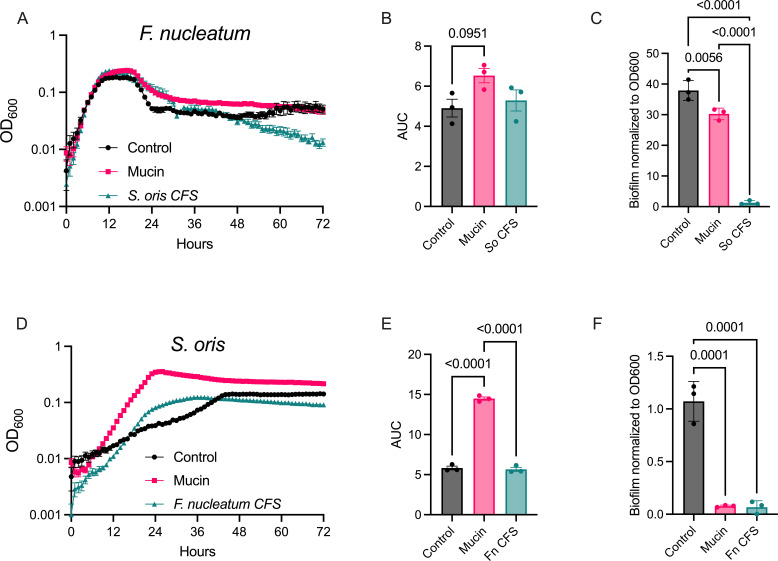 Fig 1
Fig 1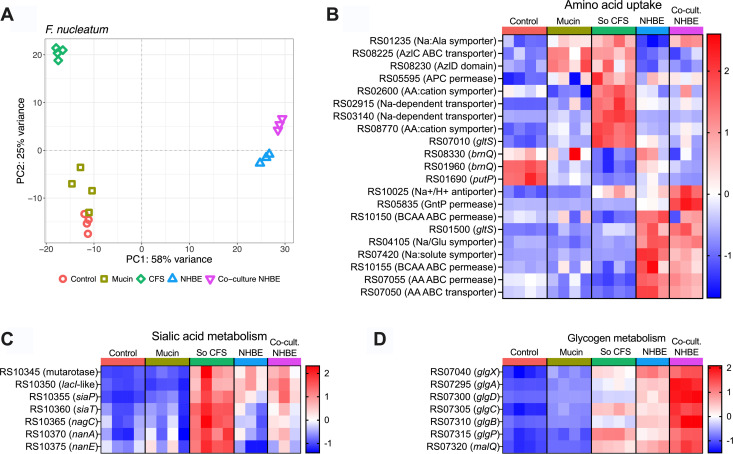 Fig 2
Fig 2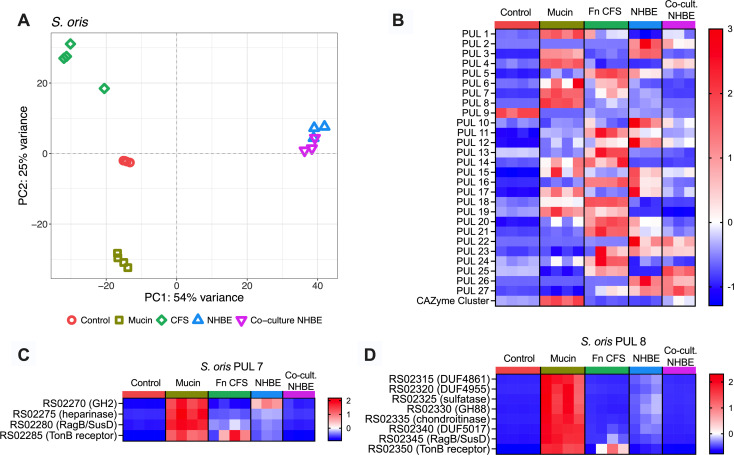 Fig 3
Fig 3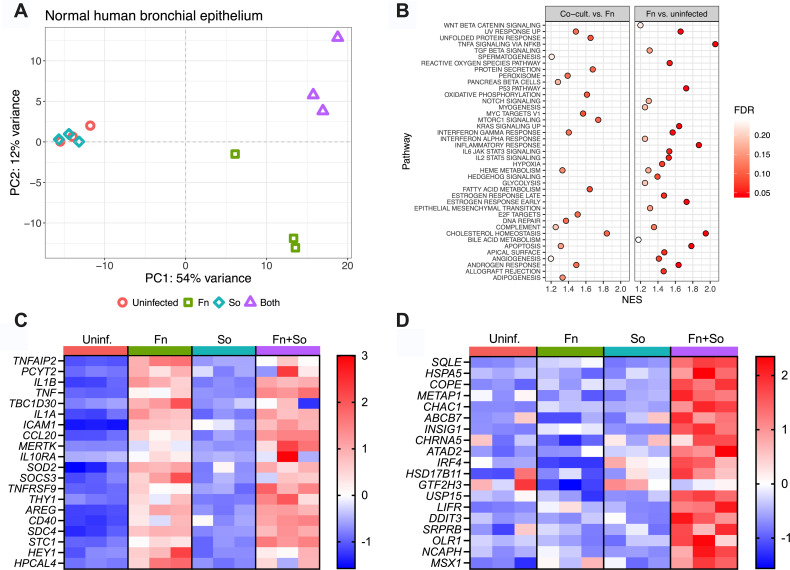 Fig 4
Fig 4- —National Institute of Allergy and Infectious Diseaseshttp://dx.doi.org/10.13039/100000060
- —Cystic Fibrosis Foundationhttp://dx.doi.org/10.13039/100000897
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Oral microbiology and periodontitis research · Otolaryngology and Infectious Diseases
INTRODUCTION
Fusobacterium nucleatum is a Gram-negative member of the oral microbiota that contributes to biofilm assembly, architecture, and community organization (1, 2). Although best known for its roles in dental plaque and periodontal disease, F. nucleatum is implicated in a range of extraoral infections and is frequently detected in tumors across multiple cancer types (3–6). While it can function as both a commensal and pathogen, F. nucleatum is often found in diverse polymicrobial host environments, including colorectal tumors and the airways of individuals with chronic rhinosinusitis (CRS), cystic fibrosis (CF), or chronic obstructive pulmonary disease (COPD) (7–14). This tendency raises questions about whether and how F. nucleatum interacts with co-colonizing microbiota and how these interactions shape its behavior and pathogenic potential. Equally important is understanding how F. nucleatum influences the activity of other taxa in these communities, many of which also exhibit context-dependent commensal or pathogenic behavior (15).
Segatella oris (formerly Prevotella oris) is also a member of the oral microbiota though less is known about its physiology or microbial interactions. Notably, S. oris supernatants inhibit the growth of the airway pathogen Moraxella catarrhalis in vitro, suggesting a connection between S. oris residence in the upper airways and colonization resistance against some pathogens (16). However, S. oris also exhibits pathogenic traits, including hemolysin production with activity against human erythrocytes (17). Like F. nucleatum, it is detected in extraoral infections, gastric tumors, CRS and CF airway mucus and is co-enriched with F. nucleatum in the oral microbiomes of cancer patients with chemotherapy-induced oral mucositis (9, 18, 19). More broadly, both Fusobacterium and Prevotella genera are co-detected in esophageal tumors (20), suggesting that cross-feeding interactions between these taxa may facilitate colonization and promote disease.
Although environmental conditions vary across infection sites, shared features such as host glycans, proteoglycans, and glycosaminoglycans are common. These features are particularly prevalent in mucus-laden airways of patients with chronic respiratory diseases and the mucus layer lining the intestinal tract, where F. nucleatum is frequently detected (21). F. nucleatum primarily ferments amino acids to generate ATP although glucose and fructose metabolism have also been reported (22–26). While it produces intracellular polyglucose granules in vitro, the genetic basis and functional significance of this pathway, including possible links to virulence, remain unclear (27). Notably, F. nucleatum lacks sialidases and polysaccharide utilization loci (PULs) but can scavenge sialic acids liberated by glycan foraging microbes (28), potentially enhancing community persistence through cross-feeding. In contrast, the genome of the S. oris type strain NCTC 13071 encodes 27 predicted PULs, likely involved in sensing, importing, and degrading of complex polysaccharides such as mucins and other host glycans (29, 30).
Given their frequent co-occurrence in mucin-rich environments in vivo and the possible complementarity of their metabolic repertoires, we hypothesized that F. nucleatum and S. oris engage in cross-feeding interactions. To test this, we examined how each species adjusts its growth and gene expression to mucin glycoproteins and each other’s metabolic byproducts. We found that while neither mucins nor S. oris supernatant substantially altered the growth of F. nucleatum, both conditions triggered significant transcriptional changes. Conversely, S. oris exhibited distinct growth dynamics and widespread transcriptional responses in mucin and F. nucleatum supernatants. Although F. nucleatum forms robust biofilms, biofilm formation was greatly reduced in S. oris supernatants and in co-culture despite biofilm growth in mucin or control media. We further extended these analyses using co-culture of each species, individually and together, on the apical surface of primary human airway epithelial cells to model aspiration-based colonization relevant to CF, CRS, and COPD. Dual (host-S. oris, host-F. nucleatum) and triple (host-S. oris-F. nucleatum) RNA sequencing revealed that host-associated growth conditions strongly influence bacterial gene expression, including unexpected variability in S. oris PUL expression mediated by F. nucleatum. Host transcriptomic responses were primarily driven by F. nucleatum, with prominent induction of pro-inflammatory mediators TNFA and TNFAIP2, as well as several genes linked to cancer. Together, these data capture key information about bacterial interactions on relevant host cell types and highlight how co-colonizing, cross-feeding partners may modify each other’s physiology and influence host responses.
MATERIALS AND METHODS
Bacterial strains and media conditions
Fusobacterium nucleatum subsp. nucleatum strain ATCC 25586 and Segatella oris strain NCTC13071 were propagated anaerobically in a Coy anaerobic chamber (90% nitrogen, 5% carbon dioxide, 5% hydrogen) using BBL Brucella broth (BD) or 1.5% agar plates supplemented with hemin (250 μg/mL) and vitamin K (50 μg/mL) (Hardy Diagnostics). A semi-defined control medium was prepared by combining Brucella broth with minimal salts (50:50) as previously described (31). A minimal mucin medium (MMM) was generated by autoclaving porcine gastric mucin (PGM; Sigma) at 30 g/L in water, diluting to 15 g/L in 2× minimal salts, and combining 1:1 with Brucella broth to yield the final mucin-containing experimental medium (hereafter referred to as mucin medium), adapted from Flynn et al. (32) with dialysis steps omitted. Cell-free supernatants (CFS) were prepared in biological triplicate by culturing each strain for 48 h in mucin medium (10 mL) under anaerobic conditions. Cultures were centrifuged at 4,000 rpm at 4°C for 20 min, and the resulting supernatants were then passed through 0.22 μm filters. CFS were stored at −80°C and thawed once immediately prior to use.
Growth curves
F. nucleatum and S. oris were grown overnight in Brucella broth supplemented with hemin (0.35 mg/mL) and vitamin K (0.05 mg/mL) and then subcultured 1:5 in fresh medium and grown for an additional 4 h. Optical density at 600 nm (OD600) was determined via spectrophotometry and adjusted to 0.01 in the respective growth medium (control, mucin, or cell-free mucin medium supernatants). Two hundred microliters of each culture was added to individual wells in a clear, flat-bottomed 96-well plate in technical triplicate across three biological replicates. Plates were sealed with a Breathe-Easy gas-permeable membrane and incubated in a Tecan Sunrise plate reader in the anaerobic chamber at 37°C for 72 h. OD600 was recorded hourly following 5 s of linear shaking. Viable bacteria were enumerated from 2 mL cultures at 0, 4, 8, and 24 h after inoculation by serial 10-fold dilution and plating onto Brucella agar supplemented with hemin and vitamin K. Plates were incubated anaerobically for 48 h prior to colony counting.
Biofilm assays
Biofilm assays were performed in parallel using identical media conditions as described for growth curves, following the protocol of Merritt et al. (33). Briefly, microtiter plates were inoculated with each species individually or in co-culture and incubated anaerobically at 37°C for 48 h. Following incubation, plates were removed from the chamber and planktonic cultures were discarded. Plates were washed three times with 250 μL water, stained with 200 μL 0.1% crystal violet (in water) for 15 min, and then washed again three times and air-dried overnight. Crystal violet was solubilized with 200 μL of 30% acetic acid, and absorbance at 550 nm was determined using a BioTek Synergy H1 microplate reader. Wells containing sterile media were used as negative controls for background subtraction.
Colonization of primary human airway epithelial monolayers
Normal human bronchial epithelial (NHBE) cells (Lonza Bioscience) obtained from healthy donors were expanded B-ALI Growth Basal Medium (Lonza) and seeded on 6.5 mm Transwell inserts (0.4 µm pore; STEMCELL Technologies). These cells produce mucus in vitro, primarily MUC5AC and MUC5B, making them excellent models for bacterial glycan foraging and cross-feeding (34, 35). Upon reaching confluency after ~2–4 days post-seeding, apical medium was removed, and basolateral medium was replaced with B-ALI Differentiation Basal Medium (Lonza). Cells were maintained at air-liquid interface (ALI) for 3–4 weeks at 37°C and 5% CO_2_ in a humidified incubator.
Overnight bacterial cultures were grown in Brucella broth supplemented with hemin and vitamin K. These were subcultured 1:5 in fresh medium and grown to OD600 = 0.5. Cultures were diluted 1:10 into infection medium (DMEM supplemented with 2% FBS, 10 mM HEPES, 0.1 mM nonessential amino acids, 4 mM L-glutamate, and 1 mM sodium pyruvate). One hundred microliters of bacterial suspension was applied to the apical surface of each Transwell and incubated for 4 h at 37°C under anaerobic conditions, resulting in an approximate MOI of 10:1 bacteria per host cell. After incubation, apical supernatants were gently removed by pipette, and co-cultures were maintained for an additional 20h prior to RNA extraction.
RNA extraction
Each species was grown in 10 mL of control, mucin, and CFS media in 15 mL conical Falcon tubes for 24 h and collected by centrifugation at 4,000 rpm for 20 min at 4°C. Pellets were dissolved by gentle pipetting in 1 mL of TRIzol Reagent (ThermoFisher). For airway epithelial co-cultures, 250 μL TRIzol was added to the apical surface and a 1 mL pipette tip was used to scrape cellular material from the transwell surface. Scraping was performed four times per insert to yield 1 mL of total lysate in TRIzol. All TRIzol samples were incubated for 5 min at room temperature, followed by the addition of 200 μL chloroform, hand agitation for 15 s, followed by another 5-min incubation on the benchtop. Phase separation was performed by centrifugation at 12,000 rpm for 15 min at 4°C. The aqueous phase was mixed 1:1 with 95% ethanol, and RNA was column purified with the Zymo Clean & Concentrator-5 kit including an on-column DNase-I treatment according to manufacturer’s instructions.
RNA sequencing and analysis
Total RNA from broth cultures was submitted to Seq Center (Pittsburgh, PA), where rRNA depletion was performed with the Illumina Stranded Total RNA Prep with Ribo-Zero Plus Microbiome kit prior to library preparation and sequencing (2 × 150 bp). RNA from bacterial and epithelial co-cultures was sent to SeqCoast Genomics (Portsmouth, NH). rRNA depletion was also performed on these samples prior to Illumina sequencing (2 × 150 bp). Sequencing was performed on the Illumina NextSeq2000 platform with a 300 cycle flow cell kit. Read quality was assessed using FastQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Given the consistently high quality and the risk of biasing downstream analyses (36), no read trimming was performed.
For bacterial RNA seq, coding sequences for all annotated genes and riboswitches, gene names, and locus tags were extracted from the F. nucleatum ATCC 25586 (NZ_CP028101) and S. oris NCTC 17031 (NZ_LR134384) genomes in Geneious Prime (2024.0.7) and converted to FASTA format via the “Tabular-to-fasta” tool via the Galaxy server (https://usegalaxy.org/), and indexed with Salmon (37) for quasi-mapping of reads. For host RNA-seq, the Homo sapiens transcriptome (GRCh38.p14, release 45) was retrieved from Gencode (https://www.gencodegenes.org/human/release_45.html). Transcriptome indices were built in Salmon, to which reads were quasi-mapped. The quant.sf files generated by Salmon were imported into RStudio via the tximport package (38), and differential expression analysis was performed with DESeq2 (39). The threshold for differential expression was a log2 fold change ≥1 at an adjusted P-value <0.05. Code for each analysis is available at https://github.com/Hunter-Lab-UMN/Fletcher_FnSo_2025.
RESULTS
Growth and biofilm formation of F. nucleatum and S. oris are influenced by mucin and cross-feeding in vitro
F. nucleatum and S. oris were cultured under three conditions: (i) control medium, (ii) control medium supplemented with porcine gastric mucin (hereafter “mucin medium”), or (iii) cell-free supernatants (CFS) derived from each species grown in mucin medium for 48 h. In the control medium, F. nucleatum exhibited distinct growth phases with a pronounced exponential phase followed by stationary and death phases, with optical density remaining consistent after ~36 h. Exponential growth rates were similar across conditions; however, cultures grown in mucin medium or S. oris CFS reached higher stationary-phase densities and exhibited a slower decline during death phase compared to controls (~12–36 h; Fig. 1A). This elevated density in mucin medium was sustained until ~60 h, after which it declined to control levels. Area-under-curve (AUC) analysis reflected this trend, with mucin medium showing a modest, though not statistically significant, increase relative to the control (P = 0.0951; Fig. 1B). In contrast, F. nucleatum density in S. oris CFS declined sharply after 48 h, yielding an AUC comparable to the control. Experiments were repeated in larger culture volumes, and CFUs were enumerated at 0, 4, 8, and 24 h (Fig. S1A and B). Each strain grew well in all media, with S. oris CFUs being more consistent between conditions than was apparent during growth in microtiter plates.
Mucin and interspecies metabolite exchange influence growth and biofilm formation of F. nucleatum and S. oris. (A) Growth curves of F. nucleatum in control medium, mucin medium, and S. oris cell-free supernatants (CFS) (n = 3). (B) Area under the curve (AUC) analysis for F. nucleatum growth. (C) Biofilm formation by F. nucleatum in each condition after 48 h, quantified by crystal violet staining. (D) Growth curves of S. oris in control medium, mucin medium, and F. nucleatum CFS. (E) AUC analysis for S. oris growth. (F) Biofilm formation by S. oris across all conditions. Data represent the mean of three biological replicates ± standard error of the mean (n = 3). Statistical significance in panels B, C, E, and F was determined using ordinary one-way ANOVA with Tukey’s multiple comparisons test. Error bars in panels A and D are the standard error of the mean (SEM).
Biofilm formation by F. nucleatum was robust in both control and mucin media, with a trend toward increased biofilm in mucin medium. However, biofilm growth was nearly abolished in S. oris CFS (Fig. 1C). Although nutrient depletion in S. oris CFS could partially contribute to reduced biofilm formation, this is unlikely given that F. nucleatum exhibited comparable planktonic growth in CFS and control medium. These findings suggest that S. oris produces one or more metabolites that inhibit F. nucleatum biofilm development. This suppression is unexpected given F. nucleatum’s well-established role as a so called bridging species in the highly structured and spatially stratified polymicrobial biofilms of the oral cavity (40).
In contrast, S. oris exhibited markedly different growth dynamics across conditions. Growth in control medium was slow with apparent diauxie following a brief stationary phase at ~24 h, a second exponential phase, and final plateau at ~40 h (Fig. 1D). This pattern may reflect initial depletion of readily metabolizable sugars followed by utilization of more complex structures such as mucin glycans, consistent with its predicted PUL repertoire. In mucin medium, S. oris grew much more rapidly, reaching a higher maximum density and exhibiting a threefold increase in AUC compared to the control (Fig. 1E). Likewise, diminished growth in F. nucleatum CFS could result from reduced availability of simple carbohydrates though the enhanced growth in mucin medium suggests that S. oris efficiently forages host glycans when preferred carbon sources are limited. These findings are consistent with genomic predictions of glycan utilization via PULs. Growth in F. nucleatum CFS yielded a faster growth rate than control medium but lower stationary phase density than mucin medium; AUC for CFS and control were comparable. Biofilm formation by S. oris was minimal in the control medium and nearly undetectable in mucin medium or CFS conditions (Fig. 1F). In dual-species cultures grown in control or mucin media, biofilms phenocopied the minimal single-species biofilms of S. oris (Fig. S1).
F. nucleatum and S. oris transcriptomes are modulated by mucin medium, cross-feeding, and interactions with human airway epithelia
Both species exhibited growth phenotypes responsive to the nutritional composition of each medium (Fig. 1). Mucin supplementation modestly enhanced F. nucleatum growth but had a pronounced effect on S. oris, suggesting species-specific nutrient utilization. Growth in CFS derived from either species grown in mucin medium was comparable to or greater than growth in the control medium, indicating that neither species fully exhausted nutrients essential to the other. Interestingly, S. oris growth was diminished in F. nucleatum CFS compared to mucin medium despite F. nucleatum lacking known glycan-degrading capabilities. Given their potential clinical relevance and prevalence of each species in chronic airway disease, we also sought to connect broth culture data to more physiologically relevant conditions with host airway epithelia. To model how each species influences the host airway response to colonization, we employed our recently described Dual Oxic-Anoxic Co-Culture (DOAC) model, in which oxygen-dependent airway epithelial cells are supplied oxygenated blood gas basolaterally while exposing their apical surfaces to the anaerobic chamber environment (41). This advancement allows for colonization and growth of obligate anaerobes on epithelial cells, enabling study of host-pathogen-microbiota interactions. We performed RNA-seq on each bacterium in control, mucin, and CFS media, as well as dual- and triple RNA-seq on primary normal human bronchial epithelial (NHBE) cells challenged with each bacterium alone or in co-culture for 24 h. This allowed us to determine how different growth environments affect the expression of genes involved in nutrient acquisition and metabolism. Differentially expressed genes are provided in File S1 (F. nucleatum) and File S2 (S. oris) and visualized in MA plots in Fig. S2 and S6. Principal component analysis ([Fig. 2A and 3A](#F2 F3)) revealed that the greatest transcriptomic separation occurred between broth cultures media and airway epithelial co-cultures, with CFS exerting the most pronounced transcriptional shift among media conditions.
Transcriptomic responses of F. nucleatum to mucin, S. oris supernatants, and epithelial co-culture is (A) Principal component analysis (PCA) of F. nucleatum transcriptomes across control medium, mucin medium, CFS from S. oris grown in mucin medium, on normal human bronchial epithelia individually or in co-culture with S. oris. (B) Heatmap of amino acid transporter gene expression across conditions. (C) Expression of sialic acid catabolism (nan operon) genes. (D) Expression of glycogen metabolism genes (see Supplemental Files for complete gene lists). Each column in the heatmaps is an individual biological replicate, and each row is a transcript. Normalized read counts from DESeq2 are scaled by row and presented as Z-scores.
The S. oris transcriptome is significantly altered across media and co-culture conditions in vitro. (A) PCA of S. oris transcriptomes, colored by growth medium or co-culture status on NHBE, across growth conditions, lot depicting the transcriptome of individual S. oris samples, which are colored by growth medium or co-culture status on NHBE. Heatmaps of (B) average expression of every gene in a given PUL across conditions, (C) PUL 7, and (D) PUL 8 in S. oris. Data are scaled by row and presented as Z-scores. Statistical significance for PUL expression comparisons provided in File S4.
F. nucleatum amino acid uptake and metabolism
Since F. nucleatum preferentially ferments amino acids, we surveyed the expression of genes involved in amino acid uptake (Fig. 2B) and metabolism (Fig. S3) to determine which substrates might be available across each growth medium or the airway epithelial environment. Two brnQ homologs encoding branched-chain amino acid (BCAA) importers and a putP proline transporter homolog were highly expressed in control medium, moderately expressed on NHBE, and showed the lowest expression in mucin medium, S. oris CFS (So CFS), and NHBE co-cultures. In contrast, homologs of the azlC and azlD BCAA importer family and a sodium:alanine symporter were upregulated in mucin medium and So CFS. Interestingly, the symporter exhibited low expression on NHBE alone but was elevated when S. oris was co-cultured on NHBE.
Several sodium-dependent symporters and transporters were specifically induced in So CFS, while multiple ABC-type transporters and permeases were selectively upregulated in NHBE co-culture, irrespective of S. oris presence. A sodium:proton antiporter and GntP-family permease were strongly induced in NHBE co-culture, but only in the presence of S. oris, suggesting interaction-dependent metabolic shifts. Genes involved in amino acid catabolism and short-chain fatty acid production were generally upregulated in NHBE co-culture with or without S. oris. Among these genes were those encoding butyrate production, which have recently been linked to reactive oxygen/nitrogen resistance in F. nucleatum (42). Overall, while these transcriptional patterns suggest condition-specific substrate utilization, further experimental validation will be required to define precise substrate specificities.
F. nucleatum carbohydrate metabolism is modulated by S. oris
Although F. nucleatum is known to catabolize glucose and fructose, other carbohydrate metabolic pathways were differentially expressed in So CFS and during co-culture with S. oris on NHBE cells. Because So CFS was derived from mucin medium-grown cultures, glycan degradation by S. oris could liberate component sugars that F. nucleatum subsequently metabolizes. For example, the nan operon (encoding a sialic acid catabolism pathway) was strongly induced in both So CFS and in NHBE co-culture conditions (Fig. 2C). Since F. nucleatum lacks a sialidase, its access to sialic acid is likely dependent on exogenous glycosidase activity from S. oris. The absence of nan induction in mucin medium supports this model.
F. nucleatum also encodes genes annotated for glycogen production and degradation though their function has not been experimentally confirmed. These genes were minimally expressed in control and mucin media but showed increased expression in So CFS and during NHBE co-culture, particularly when co-colonized with S. oris (Fig. 2D). This pattern suggests that glycogen-related pathways may participate in the utilization of glycan fragments generated by S. oris. Similarly, F. nucleatum encodes two putative α-1,4-polygalactosaminidases, enzymes that in some Pseudomonas, Streptomyces, and Aspergillus species degrade galactosaminoglycans such as chondroitin sulfate and dermatan sulfate (43–46). Alhough their expression was not strongly affected by the tested growth conditions, whether F. nucleatum homologs also degrade galactosamioglycans remains to be determined*.* More broadly, numerous genes predicted to encode carbohydrate active enzymes (CAZymes) exhibited condition-dependent expression (Fig. S4). However, most of these are not involved in host glycan degradation, as they include functions related to cell wall homeostasis (mrcB homolog C7Y58_RS05695) and LPS assembly or modification (lpxCB genes).
Expression of F. nucleatum autotransporters and adhesins is modulated by nutrient and host conditions
Virulence in F. nucleatum is mediated by several Type V autotransporters that facilitate attachment to and invasion of host cells, and modulation of immune signaling (47–51). Therefore, we evaluated the expression of all annotated autotransporter genes across each broth and host-associated condition (Fig. S5). Expression clustered into three groups: (i) genes whose expression is highest in the control medium, (ii) those with elevated expression in mucin medium or So CFS (e.g., radD), and (iii) genes with maximal expression on NHBE in both mono- and co-culture. The latter group included several well-characterized F. nucleatum virulence factor genes, such as fadA, cbpF, fusolisin, and fap2. An interesting exception to this grouping was fplA, which was highly expressed in control, mucin medium, and on NHBE, but was repressed in So CFS and in co-culture with S. oris on NHBE. These data are consistent with the increasing appreciation of the connections between bacterial metabolism and regulation of virulence factors and suggest that co-colonizing S. oris could modulate fplA-mediated contributions to F. nucleatum virulence (52).
S. oris polysaccharide utilization loci expression is responsive to both host and F. nucleatum interactions
The S. oris genome encodes 27 PULs and 1 putative CAZyme cluster, some homologous to validated systems in the Bacteroides genus (53, 54). Such systems are used by bacteria to sense and degrade polysaccharides typically found in plant fibers (e.g., xylan) or host glycans like mucins (55). We averaged the expression of each gene in every PUL to determine which PUL(s) respond to glycans present in mucin medium and on the apical surface of airway epithelial cells, as well as whether F. nucleatum supernatants or co-culture on epithelia modulate PUL expression(Fig. 3; File S3; Fig. S7). All but PUL9 were minimally expressed in control medium. Eleven PULs and the CAZyme cluster were induced in mucin medium, consistent with mucin glycan utilization. Unexpectedly, PUL expression patterns were significantly altered in Fn CFS, with decreased expression of several mucin-induced PULs and induction of multiple mucin-insensitive PULs. A third group of PULs was induced during NHBE co-culture, including PULs 3, 5, 10, 12, and 15–17, many of which were suppressed in co-culture with F. nucleatum. In contrast, PULs 4 and 25, minimally expressed in NHBE monoculture, were induced by F. nucleatum. This variable expression of PULs was anticipated between growth in mucin medium and control media and on NHBE, but modulation of PUL expression in Fn CFS and co-culture on NHBE was unexpected. These data suggest that F. nucleatum may influence glycan foraging by S. oris, supporting a dynamic bidirectional interaction.
S. oris encodes a putative Type VI secretion system
Like many members of the Bacteroidota, S. oris encodes a predicted contact-dependent multi-protein Type VI secretion system (T6SS) that delivers toxins to adjacent bacteria in intensely competitive conditions (56). Given that S. oris typically resides in polymicrobial settings and, therefore, could plausibly benefit from such a system, we scanned the S. oris NCTC 13071 genome for T6SS genes using SecReT6 (File S4) (57). Three genomic loci were identified, including one containing 13 clear homologs of known T6SS genes and 2 smaller clusters each containing 4 homologs. Expression profiles (Fig. S8) revealed moderate expression in control medium and mucin media, with higher expression of select genes in Fn CFS. The hcp homolog was only expressed in control medium, while the tssC homolog was only expressed on NHBE, irrespective of the presence of F. nucleatum. A tssI homolog (locus tag EL210_RS03325) exhibited low-to-moderate expression across broth cultures and minimal expression in monoculture on NHBE but was highly induced in NHBE co-culture with F. nucleatum, suggesting potential functional activation in response to interspecies contact. Given the lack of coordinated expression, it is unclear whether they comprise a functional T6SS and will require further experimental validation.
F. nucleatum drives airway epithelial inflammation, modulated by S. oris co-colonization
We next profiled the transcriptional response of NHBE cells following 24 h colonization with F. nucleatum or S. oris, alone or in co-culture, relative to uninfected controls (Fig. 4A; Fig. S9, File S5). As expected, several mucin genes were expressed across all conditions, consistent with the presence of an epithelial mucus layer (Fig. S10). PCA analysis revealed that S. oris alone had little effect on the host transcriptome, whereas F. nucleatum induced a distinct and divergent host response. Co-colonization clustered separately from F. nucleatum monocolonization, suggesting S. oris modulates epithelial responses either directly or indirectly via interactions that modulate F. nucleatum virulence.
Bacterial colonization alters the NHBE transcriptome. (A) PCA plot of host transcriptomes following mono- and co-colonization with F. nucleatum and S. oris. (B) Gene set enrichment analysis (GSEA) in samples from NHBE colonized with both F. nucleatum and S. oris compared to mono-colonization with F. nucleatum (left) and NHBE mono-colonized with F. nucleatum compared to uninfected cells. Normalized enrichment scores (NES) are shown on the x-axis, and the color of each point indicates false discovery rate (FDR). High NES with low FDR indicates a pathway enriched in a given comparison. Pathways enriched in both comparisons were removed from the Co-cult versus Fn plot to highlight pathways unique to this comparison, and only those pathways with an FDR < 0.25 are shown. Expression heatmaps of transcripts enriched in (C) cells mono-colonized with F. nucleatum or (D) cells dual-colonized with F. nucleatum and S. oris. Data in panels C and D are scaled by row and presented as Z-scores.
Gene set enrichment analysis (GSEA) (Fig. 4B; Files S6 and S7) identified enrichment of inflammation- and cancer-associated pathways in F. nucleatum-colonized epithelia, including TNF-α, IL-6/JAK/STAT3 signaling, IL-2/STAT5 signaling, and Wnt/β-catenin signaling. Anti-inflammatory pathways were also suppressed, including downregulation of HAS2, which encodes hyaluronan synthase. Because hyaluronan is protective against airway inflammation and hyperresponsiveness, its suppression by F. nucleatum may exacerbate these processes in diseased airways (58).
Co-colonization with S. oris altered the NHBE transcriptome relative to monocolonization with either species, underscoring the influence of microbial interactions on host inflammatory signaling. For example, the most highly induced gene in co-colonized NHBEs relative to F. nucleatum alone was PLEKHF2, whose product is involved in micropinocytosis and TNF-driven apoptosis (59). EDEM3, associated with the unfolded protein response and endoplasmic reticulum stress (60), was also upregulated during co-colonization. Interestingly, EDEM3 has been reported to translocate to the HeLa cell surface in response to the pore-forming listeriolysin O toxin from Listeria monocytogenes (61), suggesting possible parallels in epithelial stress responses.
Co-culture of both species on NHBE also influenced the expression of genes that were uniquely enriched relative to F. nucleatum mono-colonization (Fig. 4C and D). Some inflammatory markers, such as IL1-β, were induced by F. nucleatum alone and did not change in co-culture, while others, including SQLE (encoding squalene epoxidase), were upregulated only under co-colonization conditions. Together, these data reveal that (i) F. nucleatum elicits a strong pro-inflammatory transcriptional program in airway epithelial cells and (ii) co-colonization with S. oris further reshapes this response, highlighting complex, interspecies interactions that modulate host inflammation in ways not apparent during monocolonization.
DISCUSSION
The frequent co-colonization of F. nucleatum and S. oris across mucosal environments suggests potential functional interactions that influence bacterial colonization, persistence, and host responses. Using both in vitro and airway epithelial culture models, we show that these species engage in bidirectional interactions involving glycan degradation, metabolite exchange, and transcriptional modulation. Although some transcriptional overlaps were observed, the global gene expression profiles of both species in broth culture differed markedly from those during epithelial colonization, particularly in pathways related to metabolism and F. nucleatum virulence regulation. These differences likely reflect two non-mutually exclusive factors: (i) the distinct physical and nutritional environments of liquid culture vs the static, host-associated surface of NHBEs and/or (ii) variation in bacterial growth phase between conditions. Although CFUs were not measured for either species on NHBEs to directly assess growth phase, these findings underscore the importance of modeling microbial interactions under physiologically relevant conditions. Transcriptomic analyses were performed on 24 h cultures corresponding to early stationary phase for both species (Fig. 1A and D), a time point that captured active metabolism while minimizing nutrient depletion. Nonetheless, inherent growth differences between F. nucleatum and S. oris may also contribute to the observed variability in gene expression.
Despite these contextual differences, two transcriptional signals indicative of glycan-mediated interactions were consistently observed across both culture and host-associated conditions. First, the F. nucleatum nan operon, which encodes a sialic acid catabolism pathway, was not expressed in mucin medium but was strongly induced in S. oris supernatants and during co-colonization on NHBE cells. Given that F. nucleatum lacks sialidase activity, these data support a model in which sialic acid utilization depends on glycosidase activity from co-resident microbiota. This finding is consistent with previous work (28) that showed that F. nucleatum exploits sialidase-positive species to access host-derived sialic acids in dysbiotic vaginal communities. Although the nan operon is not conserved across all F. nucleatum subspecies, the broader concept of F. nucleatum promoting dysbiotic bacterial community structures through metabolic cross-feeding likely extends to multiple mucosal environments. Indeed, Queen et al. identified F. nucleatum within polymicrobial biofilms on colorectal tumors and noted enrichment of sialic acid-adjacent metabolic pathways (62). Similarly, F. nucleatum was shown to bind to and interact with Clostridioides difficile on MUC2-coated surfaces in a bioreactor model, promoting biofilm formation and broad transcriptional changes in C. difficile, including metabolic reprogramming (63). Furtado et al. further observed induction of a fructose/mannose PTS system when C. difficile was grown in the mucus layer of intestinal epithelial cells in vitro (64). Interestingly, we also detected increased expression of a homologous system in F. nucleatum grown in mucin. Although our study focused on transcriptional responses, future biochemical analyses of cell-free supernatants (e.g., LC-MS) will be important to identify specific metabolites exchanged between species and to validate the cross-feeding interactions inferred here.
A second glycan-associated transcriptional signal involved modulation of S. oris PULs. While several PULs were robustly induced by mucin medium, their expression was suppressed in the presence of F. nucleatum supernatants or during co-colonization on epithelial cells. This is notable for two reasons: (i) the large number of PULs affected and (ii) the fact that F. nucleatum has not been reported to degrade host-associated glycans and is, therefore, not expected to appreciably alter the glycan landscape of mucin medium. Porcine gastric mucin used in this study is a crude proteolytic digest and likely contains non-mucin glycans such as glycosaminoglycans, which may explain the broad induction of PULs. For example, PULs predicted to target heparin and chondroitin (e.g., PULs 7 and 8) were induced in both mucin medium and on NHBE cells but were repressed in the presence of F. nucleatum, suggesting nutrient depletion or transcriptional repression. One possible mechanism is that F. nucleatum secretes metabolic byproducts that modulate S. oris gene expression. These products could include secreted polyglucose or glycogen, which could then be available to S. oris or scavenged from dead F. nucleatum cells. Notably, F. nucleatum glycogen metabolism genes were upregulated on NHBE, particularly during co-culture with S. oris, but not in mucin medium, suggesting that glycogen is unlikely to be present in Fusobacterium supernatants.
The airway epithelial response was strongly influenced by F. nucleatum and further modulated by co-colonization with S. oris. Consistent with prior single-cell RNA-seq studies (41), F. nucleatum elicited robust inflammatory signaling, including induction of TNF-α, IL-1β, and mitochondrial stress responses suggestive of apoptosis. In contrast, NHBE colonized with S. oris alone exhibited no differentially expressed genes compared to controls, consistent with its commensal nature. Nevertheless, co-colonization with S. oris altered the epithelial response, indicating that microbial interactions can modulate host sensing and inflammation. For example, TNS4, a gene linked to poor prognosis in lung adenocarcinoma (65), was significantly downregulated in NHBEs co-colonized with S. oris and F. nucleatum relative to F. nucleatum alone. Conversely, co-culture induced expression of several genes not responsive to either bacterium individually (Fig. 4D). Because S. oris alone had minimal effect, we hypothesize that co-colonization alters F. nucleatum behavior in ways that modulate host responses. However, we cannot rule out F. nucleatum-driven changes in S. oris behavior that directly influence the airway epithelial response. Regardless, these data collectively highlight the need to consider microbial interactions when modeling host-pathogen dynamics and support integrating microbiome-derived hypotheses with mechanistic approaches to uncover host-pathogen-microbiome interactions.
In summary, our findings demonstrate that interactions between nutrient availability, the host environment, and co-colonizing bacteria shape the behavior of F. nucleatum, S. oris, and airway epithelial cells. Limitations should be acknowledged. While transcriptional and translational outputs are generally correlated, differences in mRNA and protein stability may result in temporal or spatial discrepancies in bacterial and host behavior that are not captured here. Likewise, site-directed mutagenesis is required to confirm several bacterial interactions suggested by our data, but genetic tools for F. nucleatum are only recently emerging and remain unavailable for S. oris. Finally, we evaluated only one strain of each species, which constrains our ability to assess strain- and subspecies-level genetic (and thus phenotypic) diversity. Despite these limitations, our results provide a framework for exploring glycan-driven cross-feeding and its impact on pathobiont behavior and host responses in mucosal environments.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Segata N, Haake SK, Mannon P, Lemon KP, Waldron L, Gevers D, Huttenhower C, Izard J. 2012. Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome Biol 13:R 42. doi:10.1186/gb-2012-13-6-r 4222698087 PMC 3446314 · doi ↗ · pubmed ↗
- 2Loozen G, Ozcelik O, Boon N, De Mol A, Schoen C, Quirynen M, Teughels W. 2014. Inter-bacterial correlations in subgingival biofilms: a large-scale survey. J Clin Periodontol 41:1–10. doi:10.1111/jcpe.1216724102517 · doi ↗ · pubmed ↗
- 3Kostic AD, Gevers D, Pedamallu CS, Michaud M, Duke F, Earl AM, Ojesina AI, Jung J, Bass AJ, Tabernero J, Baselga J, Liu C, Shivdasani RA, Ogino S, Birren BW, Huttenhower C, Garrett WS, Meyerson M. 2012. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res 22:292–298. doi:10.1101/gr.126573.11122009990 PMC 3266036 · doi ↗ · pubmed ↗
- 4Tahara T, Shijimaya T, Yamazaki J, Kobayashi S, Horitani A, Matsumoto Y, Nakamura N, Okazaki T, Takahashi Y, Tomiyama T, Honzawa Y, Fukata N, Fukui T, Naganuma M. 2024. Fusobacterium detected in barrett’s esophagus and esophageal adenocarcinoma tissues. Cancer Invest 42:469–477. doi:10.1080/07357907.2024.235998038913915 · doi ↗ · pubmed ↗
- 5Battaglia TW, Mimpen IL, Traets JJH, van Hoeck A, Zeverijn LJ, Geurts BS, de Wit GF, Noë M, Hofland I, Vos JL, Cornelissen S, Alkemade M, Broeks A, Zuur CL, Cuppen E, Wessels L, van de Haar J, Voest E. 2024. A pan-cancer analysis of the microbiome in metastatic cancer. Cell 187:2324–2335. doi:10.1016/j.cell.2024.03.02138599211 · doi ↗ · pubmed ↗
- 6Zhang JW, Zhang D, Yin HS, Zhang H, Hong KQ, Yuan JP, Yu BP. 2023. Fusobacterium nucleatum promotes esophageal squamous cell carcinoma progression and chemoresistance by enhancing the secretion of chemotherapy-induced senescence-associated secretory phenotype via activation of DNA damage response pathway. Gut Microbes 15:2197836. doi:10.1080/19490976.2023.219783637017266 PMC 10078122 · doi ↗ · pubmed ↗
- 7Lucas SK, Yang R, Dunitz JM, Boyer HC, Hunter RC. 2018. 16S r RNA gene sequencing reveals site-specific signatures of the upper and lower airways of cystic fibrosis patients. J Cyst Fibros 17:204–212. doi:10.1016/j.jcf.2017.08.00728826586 PMC 5817045 · doi ↗ · pubmed ↗
- 8Lucas SK, Feddema E, Boyer HC, Hunter RC. 2021. Diversity of cystic fibrosis chronic rhinosinusitis microbiota correlates with different pathogen dominance. J Cyst Fibros 20:678–681. doi:10.1016/j.jcf.2021.03.02233931358 PMC 8403624 · doi ↗ · pubmed ↗