Understanding the Epidemiology of Malaria in Zanzibar Through Molecular and Serological Analysis of Samples Collected During Reactive Case Detection
Varun Goel, Wahida Hassan, Caroline Murphy, Barbara B Choloi, Mohamed Ali, Bakari Mohamed, Abdallah Zacharia, Msolo C Dominick, Kyaw Thwai, Safia Mohammed, Shija J Shija, Jeffrey A Bailey, Anders Björkman, Billy E Ngasala, Eric Rogier, Jonathan J Juliano, Jessica T Lin

TL;DR
This study explores the role of non-falciparum malaria species in Zanzibar's recent malaria cases using blood samples and immune responses.
Contribution
The study reveals the underappreciated role of non-falciparum malaria species in Zanzibar's malaria epidemiology.
Findings
35.3% of participants had exposure to any malaria species, with Plasmodium falciparum being the most common.
Travel to mainland Tanzania was a dominant risk factor for seropositivity to all four malaria species.
Non-falciparum species showed higher seroprevalence in rural districts compared to falciparum.
Abstract
The Zanzibar archipelago has been a pre-elimination region for malaria thanks to rigorous control interventions, but recent surges in malaria cases have been observed. The contribution of non-falciparum species to the current malaria situation is unknown. This study investigates the epidemiology of falciparum and non-falciparum malaria species in Zanzibar. Leveraging plasma extracted from dried blood spots (DBS) collected during reactive case detection (RCD) activities across Unguja island from May 2022 to January 2023, we measured immunoglobulin G (IgG) responses to Plasmodium MSP1-19kD antigens using a multiplex bead-based immunoassay. Additionally, active infections were detected using species-specific real-time PCR. Out of 1618 participants surveyed in 35 RCDs, 35.3% had exposure to any malaria species, with Plasmodium falciparum being the most common (29.8%). Seroprevalences for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
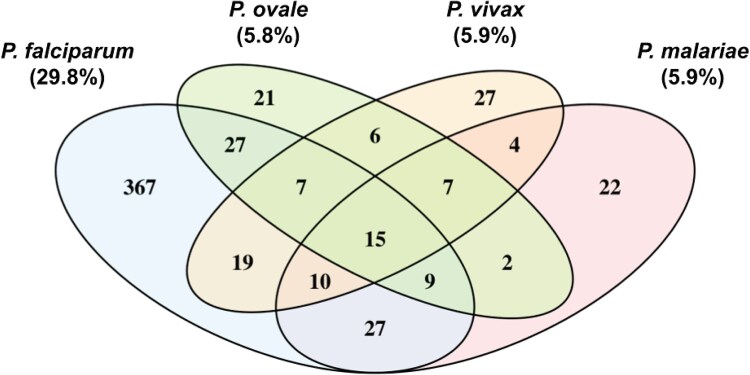 Figure 1
Figure 1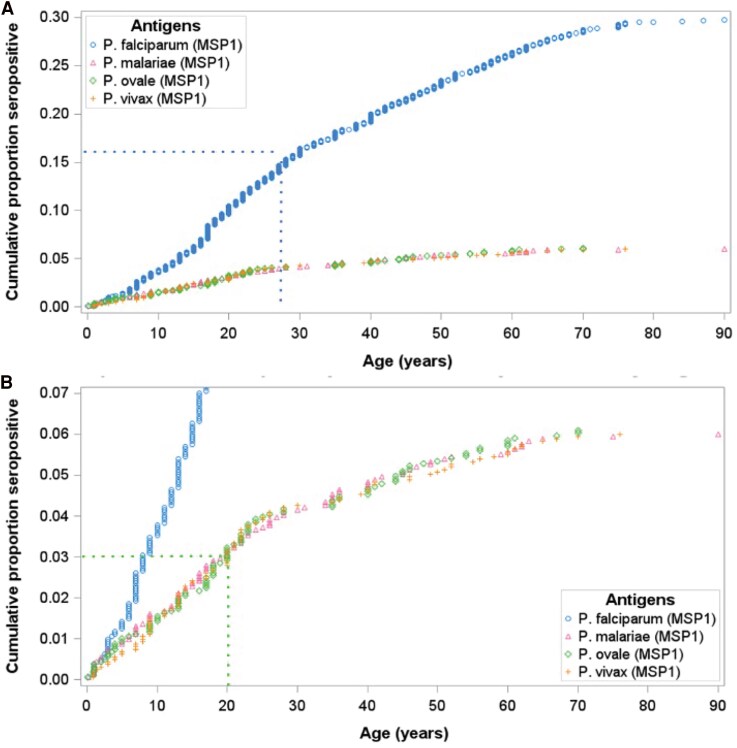 Figure 2
Figure 2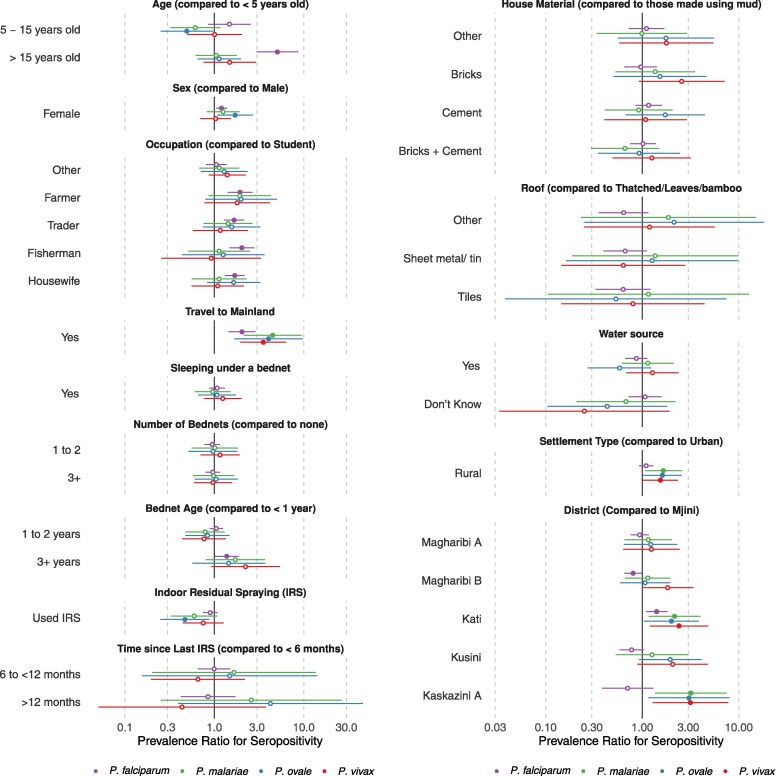 Figure 3
Figure 3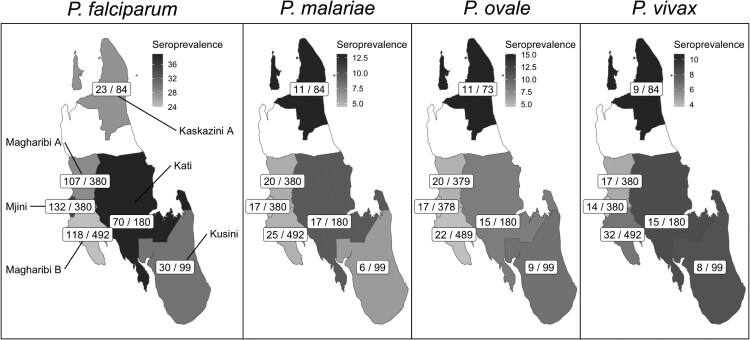 Figure 4
Figure 4| Variable | Seroprevalence | qPCR | ||
|---|---|---|---|---|
| … | N | (%) | N | (%) |
| Age | 1618 | … | 927 | … |
| <8 | 344 | (21.3) | 203 | (21.9) |
| 8–15 | 469 | (29.0) | 263 | (28.4) |
| 16–30 | 436 | (26.9) | 246 | (26.5) |
| >30 | 369 | (22.8) | 215 | (23.2) |
| Sex | 1618 | … | 927 | … |
| Female | 957 | (59.1) | 533 | (57.5) |
| Male | 661 | (40.9) | 394 | (42.5) |
| Settlement type | 1618 | … | 927 | … |
| Urban | 1171 | (72.4) | 802 | (86.5) |
| Rural | 447 | (27.6) | 125 | (13.5) |
| District | 1618 | … | 927 | … |
| Magharibi B (urban and rural) | 495 | (30.6) | 311 | (33.5) |
| Magharibi A (urban) | 380 | (23.5) | 265 | (28.6) |
| Mjini (urban) | 380 | (23.5) | 226 | (24.4) |
| Kati (rural) | 180 | (11.1) | 42 | (4.5) |
| Kusini (rural) | 99 | (6.1) | 48 | (5.2) |
| Kaskazini A (rural) | 84 | (5.2) | 35 | (3.8) |
| Occupation | 1608 | … | 912 | … |
| Student | 704 | (43.8) | 421 | (45.8) |
| Housewife | 225 | (14.0) | 119 | (12.9) |
| Child | 216 | (13.4) | 113 | (12.3) |
| Trader/business | 179 | (11.1) | 99 | (10.8) |
| Farming | 73 | (4.5) | 35 | (3.8) |
| Fishing | 66 | (4.1) | 45 | (4.9) |
| Teacher | 23 | (1.4) | 13 | (1.4) |
| Retired or unemployed | 19 | (1.2) | 12 | (1.3) |
| Public servant/NGO | 16 | (1.0) | 8 | (0.9) |
| Food service | 11 | (0.7) | 6 | (0.7) |
| Watchman | 10 | (0.6) | 2 | (0.2) |
| Construction | 8 | (0.5) | 0 | (0.0) |
| Tourism | 6 | (0.4) | 2 | (0.2) |
| Other | 52 | (3.2) | 37 | (3.9) |
| Number of bednets in home | 1618 | … | 927 | … |
| 0 | 487 | (30.1) | 316 | (34.1) |
| 1–2 | 469 | (29.0) | 281 | (30.3) |
| 3+ | 662 | (40.9) | 330 | (35.6) |
| Age of bednets | 1107 | … | 612 | … |
| <1 y | 639 | (57.7) | 372 | (60.8) |
| 1–2 y | 426 | (38.5) | 214 | (35.0) |
| 3+ y | 42 | (3.8) | 26 | (4.2) |
| Slept under bednet last night? | 1099 | … | 600 | … |
| Yes | 624 | (56.8) | 352 | (58.7) |
| No | 475 | (43.2) | 248 | (41.3) |
| Indoor residual spraying | 1559 | … | 911 | … |
| Yes | 369 | (23.7) | 259 | (28.4) |
| No | 1190 | (76.3) | 652 | (71.6) |
| Last indoor residual spraying | 353 | … | 249 | … |
| <6 m | 40 | (11.3) | 38 | (15.3) |
| 6 to <12 m | 280 | (79.3) | 190 | (76.3) |
| >12 m | 33 | (9.3) | 21 | (8.4) |
| House material | 1577 | … | 916 | … |
| Bricks and cement | 767 | (48.6) | 501 | (54.7) |
| Cement | 483 | (30.6) | 253 | (27.6) |
| Bricks | 136 | (8.6) | 50 | (5.5) |
| Stone with mud | 58 | (3.7) | 41 | (4.4) |
| Mud | 13 | (0.1) | 10 | (1.1) |
| Bamboo with mud | 11 | (0.1) | 11 | (1.2) |
| Other (combination) | 109 | (6.9) | 50 | (5.5) |
| Roof | 1479 | … | 864 | … |
| Sheet metal/tin/tiles | 1455 | (98.4) | 848 | (98.2) |
| Thatched/leaves/bamboo | 24 | (1.6) | 16 | (1.8) |
| Water source within 50 m of house? | 1512 | … | 903 | … |
| Yes | 189 | (12.5) | 113 | (12.5) |
| No | 1251 | (82.7) | 748 | (82.8) |
| Don’t know | 72 | (4.8) | 42 | (4.7) |
| Travel | 1540 | … | 919 | … |
| None | 1498 | (97.3) | 911 | (99.1) |
| Mainland Tanzania | 42 | (2.7) | 8 | (0.9) |
- —National Institutes for Allergy and Infectious Diseases10.13039/100000060
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Parasitic Diseases Research and Treatment · Travel-related health issues
Zanzibar has been a pre-elimination region for over a decade due to the rigorous implementation of malaria control interventions including test and treat with artemisinin combination therapies (ACTs), reactive case detection (RCD), use of long-lasting insecticide-treated bednets, and vector control [1]. These interventions have reduced malaria prevalence to <1%, except during outbreaks that temporarily elevate case numbers [1]. However, there have been recent surges in malaria cases in the archipelago. In 2020, the annual parasite incidence for Unguja, the main island, reached 11.5 with 12 695 cases being reported [2]. Since then several outbreaks have occurred. Given that Plasmodium falciparum is responsible for the vast majority of morbidity and mortality associated with malaria, the majority of resources have gone into its control. However, in nearby mainland Tanzania, as P. falciparum has declined, transmission of non-falciparum malaria, in particular Plasmodium ovale and Plasmodium malariae, has persisted or even increased, potentially due to differences in how those species respond to malaria control interventions [3, 4]. Amidst high levels of connectivity and intensive control efforts that have reduced falciparum malaria prevalence in Zanzibar, we pursued a dedicated investigation of the epidemiology of non-falciparum malaria on the main island of Unguja.
Active malaria infection can be detected by multiple methods including microscopy, rapid diagnostic tests (RDTs) and nucleic acid testing; the most specific and sensitive of these being PCR [5–7]. This is particularly true for non-falciparum malaria, which relies on detection through the lactate dehydrogenase (LDH) band of RDTs, which has overall lower sensitivity than histidine-rich protein II (HRP2), which is specific for falciparum malaria [8]. However, in low-transmission settings, active infections may be so infrequent that PCR methods have limited utility. Serological assays can be used to assess prior exposure to malaria in a species-specific manner, thus giving a longer-term picture of malaria transmission and a better sense of exposure in low-transmission settings [9]. Over the last decade, there have been sporadic reports of active infection with P. ovale [10–13], P. malariae [10, 12–14], and Plasmodium vivax [12–14] in Zanzibar. However, there have been no serological surveys to reflect exposure over time of the various malaria species on the archipelago.
Asymptomatic infections that persist and are not treated provide a potential reservoir for malaria transmission [15]. Reactive case detections, typically using malaria RDTs to detect asymptomatic P. falciparum cases in the vicinity of an index case identified in a clinic, is often employed to find asymptomatic infections, but misses individuals with low-density infections. We leveraged dried blood spots (DBS) collected as part of RCD activities by the Zanzibar Malaria Elimination Program on the island of Unguja, to investigate previous exposure to P. falciparum, P. malariae, P. ovale, and P. vivax and compare it to the rates of active infection detected by real-time PCR in asymptomatic individuals. Risk factors for exposure to all 4 malaria species were further investigated.
METHODS
Study Design and Data Collection
We conducted 35 RCD visits among 50–100 nearest individuals to an index case across Unguja in Zanzibar between May 2022 and January 2023, spanning 2 rounds spaced 1 month apart, across rural and urban areas. A total of 1793 participants provided DBS, along with travel histories and detailed demographic and household information. The same individuals were not necessarily sampled at each visit, and individuals who were sampled twice were only included at the first sampling for analysis.
Sample Processing
Three 6 mm punches were placed into a well of a 96-well deep-well plate for DNA extraction, which was performed using a Chelex–Tween extraction protocol [16]. A single 6 mm punch was placed in a separate deep-well plate for plasma extraction for serology, performed as previously described [17].
Immunoglobulin G Assay Analysis and Threshold Selection
We used a multiplex bead-based immunoassay to detect immunoglobulin G (IgG) against Plasmodium MSP1-19kD antigens to measure exposure to P. falciparum, P. malariae, P. ovale, and P. vivax as previously described [17]. Thresholds of mean fluorescent intensity (MFI) at which an individual is considered seropositive for a particular malaria species were selected using the 2-component finite-mixture model (FMM) method [9, 17, 18]. The FMM method has been applied to quantify malaria exposure in various settings including Tanzania. The FMM method utilizes maximum likelihood estimation approaches to determine 2 unweighted subpopulations (components) through calculation of their means and variance, yielding 2 separate distributions for putative seropositive and seronegative individuals for each IgG target. Specifically, the threshold for the analysis was chosen by calculating the exp(lognormal mean + 3 * standard deviations) of the MSP1-19kD9kD antigen values, similar to the analysis by Rogier et al [9].
Real-time PCR Detection of Malaria Species
Plasmodium falciparum parasitemia was determined using a quantitative real-time PCR targeting varATS as previously described [19, 20]. For P. ovale, P. malariae, and P. vivax detection, we conducted separate real-time PCR (qPCR) assays for each species, targeting the 18S ribosomal RNA (18S rRNA) gene, enabling both species detection and a semiquantitative assessment of parasitemia using a standard curve of diluted plasmids for parasitemia estimation (MRA-178, 179, and 180, BEI Resources, Manassas, VA) [16, 21]. A genome equivalent was based on a conversion factor of 6 plasmid copies per genome. All plates included negative controls.
Statistical Analysis
We identified associations between potential geospatial, demographic, and behavioral factors, and previous exposure to falciparum and non-falciparum malaria, by fitting robust Poisson regression models using generalized estimating equations (GEEs), adjusting for individual age and clustering within the same households. Generalized estimating equation models and FMM estimations were implemented using “geepack” [22] and “mixR” package in R version 4.2.2 [23]. We also conducted sensitivity analyses to assess whether effect estimates and results are affected by the choice of thresholds based on the FMM method, and whether there may be potential cross-reactivity in IgG responses among the 4 species-specific Plasmodium antigens. Detailed description of the sensitivity analysis and results are provided in the supplement.
RESULTS
Study Population
Out of the 1793 samples collected in the context of 2 consecutive rounds of RCD in the index household and surrounding households, 4 (0.2%) were excluded from the analysis due to missing MSP1-19kD IgG data, and 171 (9.5%) were removed as they were associated with participants who had previously participated in the first RCD round. In total, samples from 1618 participants who participated in 35 RCD rounds from May 2022 to January 2023 were included in the final seroepidemiology analytical dataset. The PCR cohort represented a subset of those included in the seroepidemiology analysis, with 928/1618 (57%) participants sampled from May 2022 to September 2022 also tested for both active falciparum and non-falciparum infections using qPCR.
Overall, there were no notable differences between participants included in the seroepidemiology and qPCR cohorts, although the proportion of qPCR participants in urban settlements was higher compared to the seroepidemiology cohort (Table 1). For the seroepidemiology cohort, the median (interquartile range [IQR]) age was 15 (8–29) years, with more female (59%) participants. Most participants lived in urban settlements (72%) in Magharibi B, Magharibi A, and Mjini. Participants mainly consisted of students (44%), followed by housewives (14%), children (13%), and those engaged in trade or business (11%). Only 2.7% of participants reported traveling outside of Zanzibar to mainland Tanzania in the last 28 days. Additionally, 30% of participants reported having no bednets, while 59% reported not sleeping under a bednet the previous night. Twenty-four percent of participants reported indoor residual spraying (IRS) with 11% reporting last IRS within 6 months. Most participants (88%) reported living in houses made of brick, cement, or both, and almost all participants (98%) lived under a sheet metal, tin, or tiled roof.
Seroprevalence Estimates
Out of the 1618 participants tested for anti-MSP1-19kD IgG antibody responses, 35% (n = 571/1617) had evidence for exposure to any malaria species. The most common exposure was P. falciparum with 30% of participants (n = 482/1617) being seropositive. Seroprevalence for each non-falciparum malaria species was much lower; P. ovale was 5.8% (n = 94/1611), while P. malariae and P. vivax seroprevalences were 5.9% (n = 96/1617 and n = 95/1617) each (Figure 1). Among participants with seroprevalence results available for all 4 malaria species (n = 1611), exposure to multiple malaria species was common with 133 (23% out of 570) having exposure to 2 or more species, and 15 (2.8%) exposed to all 4 species. Overall, 36% (203/570) of the malaria-exposed group demonstrated exposure to non-falciparum species; nearly half of these (44%, 89/203) showed exposure to non-falciparum species in the absence of exposure to falciparum.
Overlap of seroprevalence across all 4 malaria species. In total, immunoglobulin G (IgG) versus MSP1-19kD responses were measured across 1618 Zanzibar residents participants in reactive case detection after a malaria case was identified in proximity (in their or a neighboring household). Note: Percentages for Plasmodium ovale were calculated for 1611 individuals with successful seroprevalence results. Percentages for other species were calculated for 1617 individuals. A total of 1611 participants had seroprevalence results available for all 4 species.
We conducted sensitivity analysis by adopting a very stringent threshold at 5SD instead of 3SD, obtaining seroprevalence estimates for P. falciparum, P. ovale, P. malariae, and P. vivax of 24%, 2.4%, 3.0%, and 2.0%, respectively (Supplementary Tables 1 and 2). In this analysis, 23% (100/429) of the malaria-exposed group demonstrated exposure to non-falciparum species; nearly half of these (42%, 42/100) showed exposure to non-falciparum species in the absence of exposure to falciparum. To assess for potential cross-reactivity responses across antigens, we also examined correlation of MFI responses across all 4 MSP1-19kD orthologous antigens (Supplementary Figure 1), and the distributions in MFI responses for individuals with mono-species versus mixed-species exposure (Supplementary Figure 2). Although participants seropositive against multiple species had slightly higher MFIs to Po and Pm MSP1-19kD than those seropositive against only 1 species-specific antigen, those differences were not significant, even when assuming a 5SD threshold (Supplementary Figure 3).
We also used our cross-sectional data to estimate cumulative proportion of exposure over the lifespan to understand when exposure is usually acquired (Figure 2). Overall, approximately 30% participants were exposed to P. falciparum, with a steady increase in the proportion of participants seropositive over childhood years. By age 27 years, approximately 15% (half the total seroprevalence) of individuals demonstrated exposure. For non-falciparum infection, the rate of acquisition was similar between species and perhaps slightly faster than for P. falciparum, with half of those (3%) exposed by the age of 20 years.
Cumulative exposure to malaria. (A) Overall cumulative exposure to all 4 malaria species tested. (B) Limited view at lower proportions to assess non-falciparum cumulative exposure.
PCR Testing and Active Infections
Compared to seroprevalence estimates, rates of active infection were much lower. Overall, 56/928 (6.0%) were infected with any malaria species. Forty-three (4.6%) tested positive for P. falciparum parasitemia, with 77% of those participants (n = 33) having very low parasite density of <1 parasite/μL (Supplementary Figure 4). Similarly, few active non-falciparum infections were detected; P. malariae parasitemia was detected in 11 (1.2%), P. ovale in 4 (0.4%), and P. vivax in only 2 (0.2%) participants. Among these cases, only 2 reported recent mainland travel, to the Simayu region, Lake Zone (P. malariae case), and to Dar es Salaam (P. ovale case). Both P. vivax parasitemias were detected in the same household, in a housewife and a 4-month-old infant, both Zanzibari residents without recent travel.
Factors Associated With Seropositivity
Given low active infection rates by qPCR, especially for non-falciparum species, we only examined potential factors associated with prior malaria exposure. In this exploratory analysis, demographic factors associated with seropositivity showed differing trends for falciparum versus non-falciparum malaria (Figure 3; Supplementary Table 3). For P. falciparum, seroprevalence among persons greater than 15 years of age was 5.1 (95% CI: 3.0–8.7) times compared to under-5 children. In contrast, for P. ovale, children between 5 and 15 years of age had half the seroprevalence (PR = 0.49, 95% CI: 0.25–0.97) of P. ovale compared to under-5 children, with a similar trend for P. malariae (PR = 0.61, 95% CI: 0.32–1.16). Females, compared to males, had higher seroprevalence ratios for P. falciparum (PR = 1.2, 95% CI: 1.1–1.4) and P. ovale (PR = 1.7, 95% CI: 1.1–2.7) while controlling for age and clustering of individuals within households. Compared to students, occupations such as farming, trading, and fishing or being a housewife were associated with higher seroprevalence for P. falciparum, but not for any non-falciparum species. Associations among demographic factors and non-falciparum malaria were not significant after assuming a 5SD seropositivity threshold (Supplementary Figure 5).
Factors associated with seropositivity for all 4 species. Seroprevalence ratios were estimated by fitting robust Poisson regression models using generalized estimating equations (GEEs) to each malaria species, adjusting for individual age and clustering of individuals within the same households. Error bars denote 95% confidence intervals. Filled point estimates denote results with P < .05.
Overall, travel to mainland Tanzania in the last 28 days was the factor most strongly associated with high seropositivity for all 4 species. Mainland travel was associated with twice the seroprevalence for P. falciparum (PR = 2.0; 95% CI: 1.4–2.9), and upwards of 3 times the seroprevalence for all non-falciparum malaria species (Figure 3). Sleeping under a bednet the previous night or number of household bednets was not associated with seropositivity for both falciparum and non-falciparum species. Bednets older than 3 years, however, were associated with higher P. falciparum exposure (PR = 1.37; 95% CI: 1.01–1.87) compared to less than a year-old bednets. Indoor residual spraying was associated with a protective effect only against P. ovale seroprevalence (PR = 0.47, 95% CI: 0.25–0.88). Household construction materials, roof type, and water source were not associated with seropositivity for any species. In our sensitivity analysis assuming a 5SD threshold, these associations persisted, with the exception of the association between IRS and P. ovale seroprevalence (Supplementary Figure 5).
Geographic Distribution of Malaria Exposure
We observed geographic differences in seroprevalence across Unguja for falciparum and non-falciparum malaria. Rural areas were associated with higher seroprevalence for all 3 non-falciparum species, but not with P. falciparum (Figure 3). Additionally, districts with the highest P. falciparum seroprevalence were not the same as districts with the highest seroprevalence for each non-falciparum species (Figure 4). The highest P. falciparum seroprevalence was found in the central district of Kati followed by the western district of Mjini, while higher non-falciparum seroprevalence (P. malariae, P. ovale, and P. vivax) was concentrated in Kaskazini A district in northern Unguja. Models controlling for age and household clustering also show that when compared to an urban district such as Mjini, non-falciparum seroprevalence ratios were significantly higher for rural districts such as Kati and Kaskazini A, but not other urban districts. Even with a 5SD seroprevalence threshold, the positive associations between non-falciparum seroprevalence and rurality persist, especially for P. malariae, although 95% CIs crossed the null for other non-falciparum species (Supplementary Figure 5). However, we also note that overall P. falciparum seroprevalence within the districts of Unguja is much higher (ranging from 25% to 35%), compared to non-falciparum species (ranging from 6% to 14%).
Map of geographic distribution of seroprevalence across all 4 species. Each district is shaded based on the species specific seroprevalence. Within each district is the number of seropositive samples (# positive/# tested).
DISCUSSION
We conducted a comprehensive epidemiological study of the four human malaria species in Zanzibar and compared exposure to active infection detected by qPCR in asymptomatic individuals. After nearly two decades of maintaining pre-elimination status, exposure to P. falciparum remains common (30%), perhaps attesting to both the longevity of IgG responses and ongoing malaria exposure, whether on Zanzibar or elsewhere. The longevity of anti-MSP1-19kD IgG responses varies by age and previous exposure as well as the type of stimulus, but MSP1-19kD is known to be a highly immunogenic malaria antigen with durable and detectable IgG levels in plasma for months to years, possibly decades [24–27]. Cumulative prevalence of exposure increased steadily with age, with about 50% of exposure occurring in the first three decades of life. Serological tools to measure (possibly distant) previous malaria exposure are especially important in low-transmission settings—as highlighted by the low rates of PCR-detected active infection in this study with only 4.6% of individuals infected with P. falciparum, mostly at very low density, under <1 parasite/μL blood.
Exposure to non-falciparum infection was lower but non-negligible and similar between the three species tested, at approximately 6%. Cumulative exposure for these species reached its midpoint slightly earlier, around 20 years of age. Though active infections were uncommon, they were more prevalent than previously seen in a large PCR survey conducted 6 years earlier, in 2016 [13]. Though only 2.7% of the study participants reported traveling outside of Zanzibar to mainland Tanzania in the last 28 days, exposure to non-falciparum infection was significantly higher in travelers compared to those not reporting recent travel. Although we did not collect data on more remote travel, Plasmodium malariae, P. ovale, and P. vivax all have biological mechanisms that can lead to persistent or chronic latent infection that may support maintenance of detectable IgG levels even if exposure was more remote.
Similarity of seroprevalence among non-falciparum species might raise concerns about cross-reactivity of IgG responses to orthologous MSP1-19kD antigens. Several of our analyses support that these signals represent true exposure. In this study, MFI intensity did not seem to vary with co-infection. Cross-reactivity between antibodies generated by a falciparum exposure and a non-falciparum antigen would likely present as lower non-falciparum MFI in the co-infections. Also, no specific patterns of correlation were determined. These antigens have been used extensively for seroepidemiology studies, and the risk of cross-reactivity has been studied and deemed to be low [18]. Given this, the reported seroprevalence is likely to be representative of exposure in this population. Applying more stringent cutoffs for seropositivity still yielded evidence of exposure to each non-falciparum species in the study population.
The significantly higher seroprevalence for P. falciparum among those older than 15 years of age may be explained by the fact that they had been exposed to falciparum before intensified malaria control and elimination in 2007–2008 in Zanzibar [1, 28]. While some seropositivity may have persisted, re-exposure in more recent years might have made the immune responses more significant, durable, and thus, detectable [24, 25]. In contrast, seropositivity to non-falciparum malaria was not significantly higher in adults compared to children, which may reflect relatively higher exposure to these parasites in more recent years. Interestingly, P. ovale and P. malariae seropositivity appeared higher in younger under-5 children than older children (5–15 years). This may also suggest a recent exposure in the population. Besides local exposure in rural areas, younger children are likely exposed when traveling with their parents to the mainland [29]. Potential reasons for a new relative emergence of non-falciparum malaria infections may be that they are not as readily diagnosed and treated. They cause less symptoms and are less detectable by mRDTs, leading to cryptic transmission, especially in the absence of P falciparum infections [30]. Plasmodium ovale and P vivax also require primaquine treatment of liver stages and are therefore not radically treated by ACTs only.
The relative geographic distribution of exposure to different malaria species was not the same. Plasmodium falciparum exposure was highest in Kati district in central-east Unguja. Kati has had relatively high annual parasite incidence (API) in the years leading into this study, with an API of 18.3 in 2020 and 11.6 in 2021 [2]. In 2022, the API was lower (5.3), which may in part be reflected in the infrequently detected active infections by qPCR during the study (43/928). The seroprevalence of all three non-falciparum species was highest in Kaskazini A, a lower-transmission area with APIs of 7.6, 3.3, and 1.7 in 2020, 2021, and 2022, respectively [2]. This is not surprising, as studies of active infection by non-falciparum malaria in Tanzania have shown elevated risks in lower-transmission regions [3, 4, 16].
Limitations to this study include its cross-sectional nature and possible bias toward increased malaria exposure compared to the general population, based on its RCD study design. This sampling strategy targets at-home asymptomatic persons during visits by the malaria surveillance officer. Therefore, it does not address the epidemiology of symptomatic malaria and likely under-samples populations engaged in occupational activities outside home. Our study was not large enough to perform fully adjusted multivariate analyses for factors associated with seropositivity. While the longevity of anti-MSP1-19kD IgG responses has been studied for P. falciparum, less is known about the longevity of response to the orthologous antigen in non-falciparum species. Additionally, despite the increased sensitivity of PCR for detecting non-falciparum species, the assays used likely still miss the lowest-density parasitemias that can be found in these species. Finally, given the possibility of persistent, latent, and relapsed infection, we are unable to determine whether the few active parasitemias of non-falciparum species were acquired locally or not.
Despite relatively low transmission, exposure to malaria, in particular falciparum malaria, remains high in Zanzibar with over one-third of the population being seroreactive to any malaria species. Immediately prior to data collection, there was a major surge of malaria in 2021, potentially highlighted by the high seroprevalence despite lower levels of active infection during 2022–2023. Seroreactivity to non-falciparum infections was also prevalent, including to P. vivax, which has historically been low in Africa [31]. Our data suggest that these species appear to have become relatively more prevalent and epidemiologically more important in recent years in Zanzibar. Further research is needed to better understand the context of exposure to non-falciparum malaria species. Questions remain over whether non-falciparum infections are escaping being adequately treated and/or whether they are escaping present vector control measures. Beyond targeting travelers, it is unclear whether current elimination efforts focused on P. falciparum will adequately address exposure to non-falciparum species.
Supplementary Material
ofag051_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Björkman A, Shakely D, Ali AS, et al From high to low malaria transmission in Zanzibar-challenges and opportunities to achieve elimination. BMC Med 2019; 17:14.30665398 10.1186/s 12916-018-1243-z PMC 6341737 · doi ↗ · pubmed ↗
- 2Ali MH, Kitau J, Ali AS, et al Malaria elimination in Zanzibar: where next? Pan Afr Med J 2023; 45:7.10.11604/pamj.supp.2023.45.1.39804 PMC 1039511137538363 · doi ↗ · pubmed ↗
- 3Yman V, Wandell G, Mutemi DD, et al Persistent transmission of Plasmodium malariae and Plasmodium ovale species in an area of declining Plasmodium falciparum transmission in eastern Tanzania. P Lo S Negl Trop Dis 2019; 13:e 0007414.31136585 10.1371/journal.pntd.0007414 PMC 6555537 · doi ↗ · pubmed ↗
- 4Sendor R, Mitchell CL, Chacky F, et al Similar prevalence of Plasmodium falciparum and non-P. falciparum malaria infections among schoolchildren, Tanzania. Emerg Infect Dis 2023; 29:1143–53.37209670 10.3201/eid 2906.221016 PMC 10202886 · doi ↗ · pubmed ↗
- 5Dinko B, Ayivor-Djanie R, Abugri J, et al Comparison of malaria diagnostic methods in four hospitals in the Volta region of Ghana. Malariaworld J 2016; 7:5.38601360 10.5281/zenodo.10797112 PMC 11003212 · doi ↗ · pubmed ↗
- 6Laban NM, Kobayashi T, Hamapumbu H, et al Comparison of a Pf HRP 2-based rapid diagnostic test and PCR for malaria in a low prevalence setting in rural southern Zambia: implications for elimination. Malar J 2015; 14:25.25888818 10.1186/s 12936-015-0544-3PMC 4340619 · doi ↗ · pubmed ↗
- 7Mpina M, Stabler TC, Schindler T, et al Diagnostic performance and comparison of ultrasensitive and conventional rapid diagnostic test, thick blood smear and quantitative PCR for detection of low-density Plasmodium falciparum infections during a controlled human malaria infection study in Equatorial Guinea. Malar J 2022; 21:99.35331251 10.1186/s 12936-022-04103-y PMC 8943516 · doi ↗ · pubmed ↗
- 8Hopkins H, Kambale W, Kamya MR, Staedke SG, Dorsey G, Rosenthal PJ. Comparison of HRP 2- and p LDH-based rapid diagnostic tests for malaria with longitudinal follow-up in Kampala, Uganda. Am J Trop Med Hyg 2007; 76:1092–7.17556616 · pubmed ↗