Development of Novel Small-Molecule Targeting SCN1A-Associated Severe Myoclonic Epilepsy of Infancy
Dong Gun Kim, Kyu-Seok Hwang, Se Hwan Ahn, Seong Soon Kim, Yuji Son, Sung Bum Park, Won Hoon Jung, Dae-Seop Shin, Sung Hee Cho, Byeong Wook Choi, Pyeongkeun Kim, Yerim Heo, Minhee Kim, Jung Yoon Yang, Kyeong-Ryoon Lee, Hyang-Ae Lee, Jihun Kim, Hoon-Chul Kang, Ki Young Kim

TL;DR
Researchers developed a new drug candidate, compound 20e, that shows strong antiseizure effects in a rare and severe childhood epilepsy caused by SCN1A mutations.
Contribution
A novel small-molecule, compound 20e, was identified and shown to effectively reduce seizures in multiple models of SMEI.
Findings
Compound 20e reduced seizure severity and delayed onset in SCN1A+/– mice.
It normalized abnormal brain activity in patient-derived neurons and improved behavioral seizure parameters in zebrafish.
Compound 20e showed favorable safety and pharmacokinetic profiles, including good blood-brain barrier penetration.
Abstract
Severe myoclonic epilepsy of infancy (SMEI, Dravet syndrome), which is mainly caused by the SCN1A mutation, is a severe epileptic encephalopathy that manifests in infancy and leads to intractable seizures and developmental impairment. To discover new therapeutic chemotypes, we established a Nav1.1 (scn1lab) KO zebrafish model for chemical screening and identified novel 1,3,4-oxadiazol-2(3H)-one derivatives. Among them, compound 20e showed the most potent antiseizure efficacy in zebrafish behavioral assays and significantly reduced locomotion-related seizure parameters compared with repositioned drugs. In SCN1A +/– mice, 20e reduced seizure severity, delayed onset, and suppressed hyperactivity. Notably, 20e normalized pathological spike and burst activity in SMEI patient-derived iPSC neurons. Mechanistically, 20e appears to elevate 5-HT levels via TPH2 upregulation. It demonstrated…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1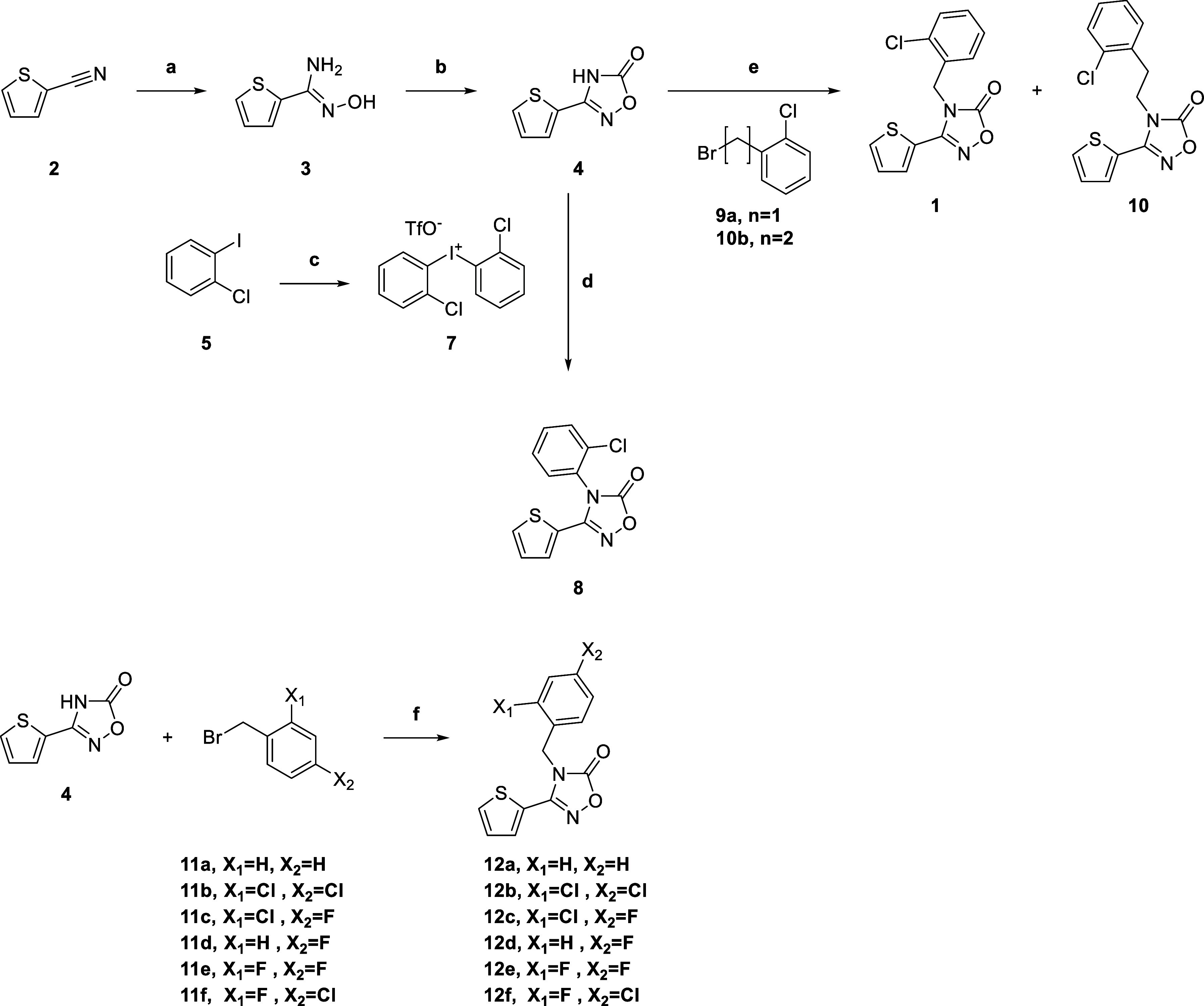 1
1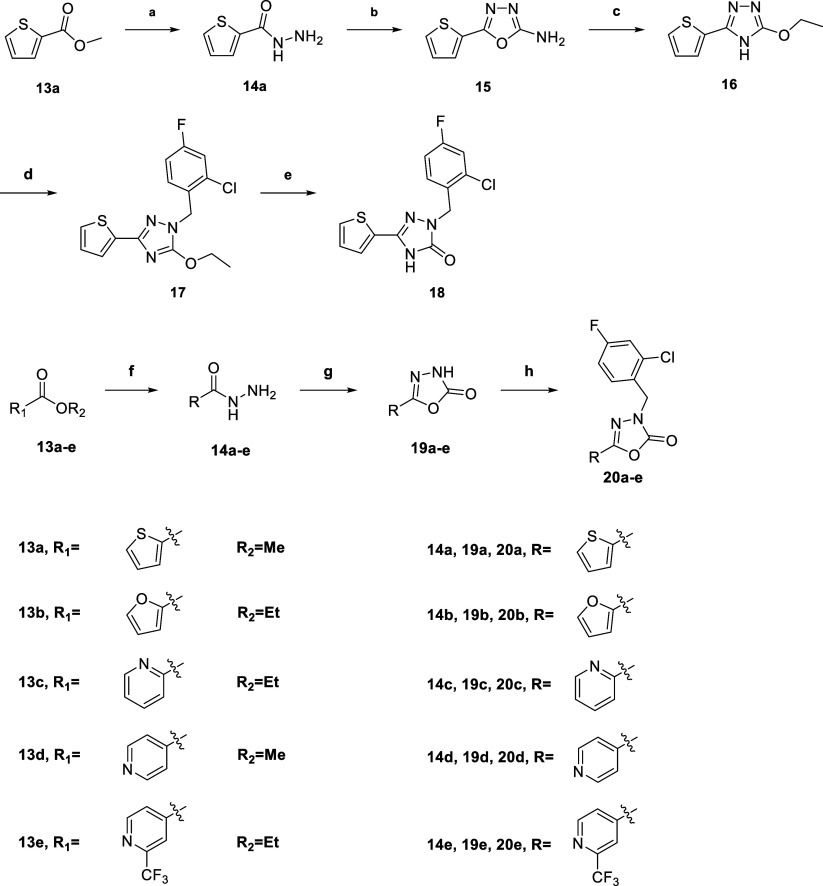 2
2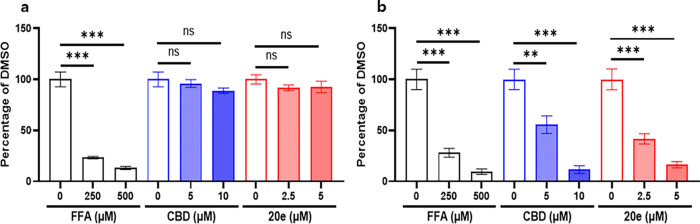 2
2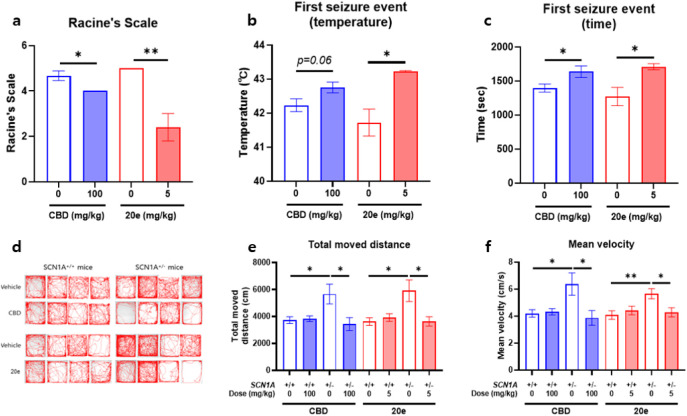 3
3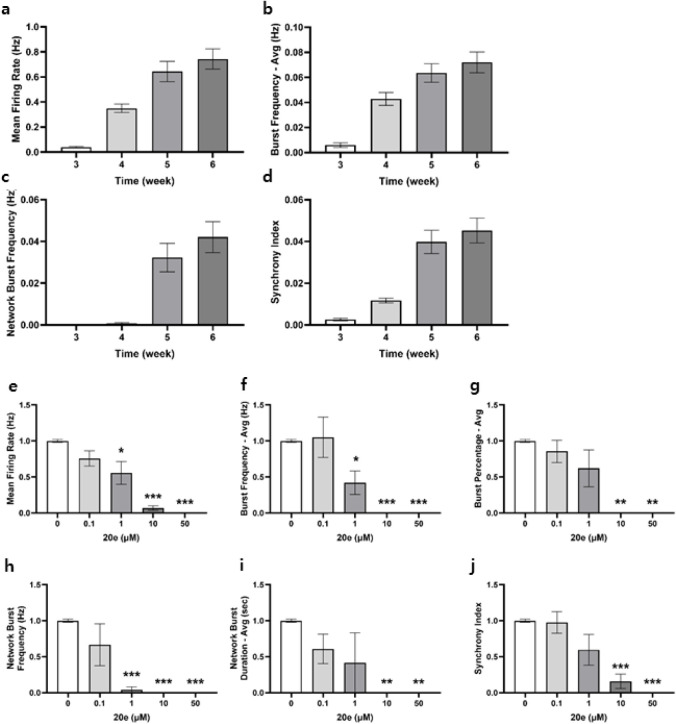 4
4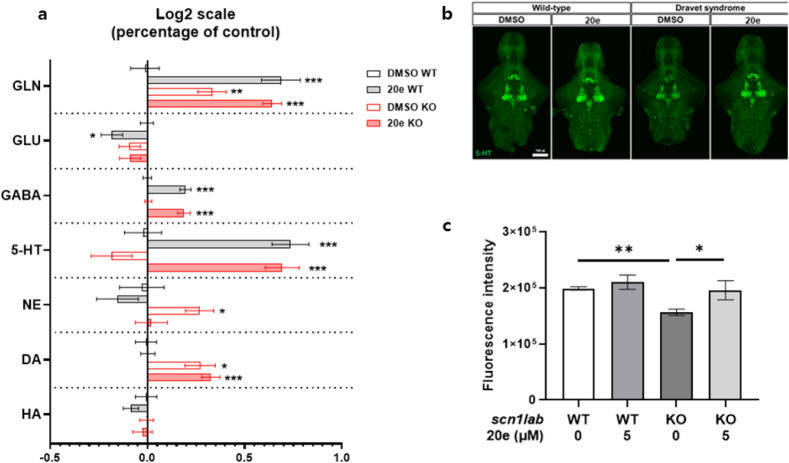 5
5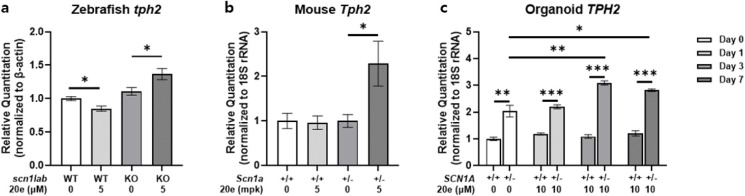 6
6| Matrix | |||||
|---|---|---|---|---|---|
| Test compound | Model | Drug administration | Plasma conc.(ng/mL) | Brain conc.(ng/g) | B/P ratio |
| a) | |||||
|
| Mouse | P.O. (10 mg/kg) | 157 ± 115 | 706 ± 619 | 4.36 ± 0.89 |
| Zebrafish | P.O. (10 mg/kg) | 951 ± 258 | 1699 ± 779 | 1.75 ± 0.57 | |
| Human BBB-chip | 10 μM | 366 ± 10 | 157 ± 10 | 0.43 ± 0.07 | |
- —Ministry of Trade, Industry and Energy10.13039/501100003052
- —Korea Research Institute of Chemical Technology10.13039/501100003704
- —Korea Health Industry Development Institute10.13039/501100003710
- —National Research Foundation of Korea10.13039/501100003725
- —National Research Foundation of Korea10.13039/501100003725
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGlycogen Storage Diseases and Myoclonus · Epilepsy research and treatment · Zebrafish Biomedical Research Applications
Introduction
Severe myoclonic epilepsy of infancy (SMEI), also known as Dravet syndrome, is a rare disease that begins in infancy and proceeds with accumulating morbidity that significantly impacts individuals throughout their lifetime. Infant-onset seizures present as prolonged periods, generalized or unilateral, clonic or tonic-clonic, and fever symptoms, accompanied by intellectual and behavioral disorders.?
In the past decade, three antiseizure drugs (ASDs) were available as therapeutic agents in SMEI: fenfluramine (FFA), stiripentol (STP), and cannabidiol (CBD). These drugs were not originally developed for SMEI. But they have been repositioned in attempts to treat SMEI. In 2018, STP was approved in the US for the treatment of seizures associated with SMEI and proved to increase GABAergic transmission. However, STP has side effects including insomnia, ataxia, hypotonia, and dystonia. CBD is approved as an adjunctive treatment for seizures in SMEI and Lennox–Gastaut syndrome (LGS) in 2018 in the USA and in 2019 in the EU. CBD has side effects such as diarrhea, vomiting, fatigue, fever, sleepiness, and abnormal liver function. Recently approved as an adjuvant therapy for SMEI, FFA was first revealed as an antiseizure agent via modulating the serotonergic system.? Despite being a withdrawn drug as an appetite suppressant, FFA was approved as a drug repositioning for SMEI treatment.? However, FFA can cause side effects, such as sedation, insomnia, and weight loss. Additionally, Brenot and colleagues suggest that fenfluramine derivatives are strongly suspected etiological agents of primary pulmonary hypertension in female cases.? These limitations underscore the need for novel therapeutic approaches with improved efficacy and safety profiles.
More than 80% of SMEI patients exhibit mutations in the sodium voltage-gated channel alpha subunit (SCN1A), which encodes the type I voltage-gated channel (Nav1.1) alpha subunit.? The voltage-gated sodium channel plays an important role in the initiation and propagation of the action potential of neurons in the brain.? Baraban et al. identified a zebrafish with a mutation in scn1lab (sodium channel, voltage-gated, type 1 like, alpha b), didy ^ s552 ^, and demonstrated the presence of spontaneous seizure phenotypes in scn1lab mutants using electrophysiological recording and behavioral analysis. They used this model for SMEI and discovered clemizole, which is the repositioned molecule developed as a histamine H1 receptor antagonist that inhibits convulsive behaviors and electrographic seizures.? It prompted us to identify a novel chemotype using a new phenotype-based screening. We established a new scn1lab knockout (KO), kri111 allele, by inserting four nucleotides in exon 2 using the CRISPR/Cas9 method. This established mutant was used to identify novel antiseizure agents for SMEI. We performed a phenotype-based screening of 6,625 small molecules from the Korea Chemical Bank library. Oxadiazolone (1) was identified as a novel hit scaffold and was optimized to identify candidates for SMEI treatment. Herein, we report the synthesis and biological evaluation of novel oxadiazolone derivatives for SMEI.
Results
In this study, we generated a new scn1lab KO zebrafish using the CRISPR/Cas9 method to evaluate the antiseizure efficacy of synthesized compounds. A 4-bp insertion was created in exon 2 of the scn1lab genome, generating a premature stop and truncated Scn1lab protein (referred to as the kri111 allele) (Figure S1a). scn1lab KO (homozygous kri111 allele) larvae showed similar morphological phenotypes to those observed in didy ^ s552 ^ including hyperpigmentation and an uninflated swim bladder (Figure S1b). Based on the three stages of seizure phases in zebrafish larvae: hyperactivity (stage 1), rapid “whirlpool-like” behavior (stage 2), and loss-of-posture (stage 3), we newly defined seizure-like movements to quantify high-speed movement at a speed above 80 mm/s. ?,? Since hyperactivity showed irregularities in scn1lab KO larvae, it is difficult to quantify seizure-like movement patterns through normal movements. However, only calculating high-speed movements above 80 mm/s can quantify hyperactivity. Indeed, the differences in normal movement between WT and scn1lab KO varied, whereas scn1lab KO movements are more increased than those of WT at a speed above 80 mm/s (Figure S1c).
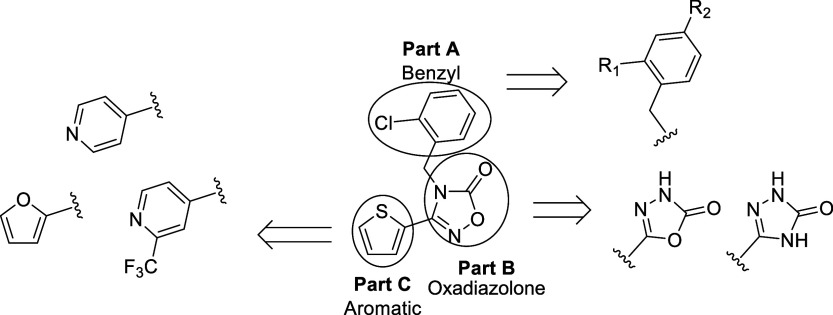
We tried to identify a novel molecule that significantly reduced seizure-like movements compared with control groups when administered to scn1lab KO larvae and showed no behavioral change when administered to WT larvae. As a result of compound screening using the chemical library from the Korea Chemical Bank (comprising 6,625 small molecules), compound 1 was identified as a hit. It significantly decreased seizure-like movements in the scn1lab KO larvae compared to those in the control group (55.3%, 50.0%, 54.6% in distance moved, movement frequency, and movement duration, Table). Whereas compound 1 showed no significant difference in WT larvae compared to the control group (103%, 111%, 105% in distance moved, movement frequency, and movement duration, Table). We optimized the hit compound 1 by systematic modification of structural parts A, B, and C as shown in Figure.
Structure and modification of compound (1).
The synthetic route for 1,2,4-oxadiazolone derivatives is outlined in Scheme. Commercially available 2-cyanothiophene (2) was first coupled with hydroxylamine to give 3, followed by cyclization to produce 1,2,4-oxadiazolone 4. The phenyl-substituted 1,2,4-oxadiazolone (8) was obtained by coupling 4 with 7, which was prepared from 2-chloroiodobenzene (5). Compound 4 was alkylated with benzyl and phenethyl halides (9a and 10b) to afford 1 and 10, respectively. Diverse benzyl derivatives (12a–f) were synthesized via the reaction of 1,2,4-oxadiazolone 4 with benzyl bromide (11a) and substituted benzyl bromides (11b–f).
Synthesis of Compounds 1, 7, 10
The synthetic pathway for 1,2,4-Triazolone and 1,3,4-oxadiazolone derivatives is depicted in Scheme. Methyl thiophene-2-carboxylate (13a) was treated with hydrazine to give hydrazide (14a), which underwent cyclization with BrCN to give oxadiazole (15). This oxadiazole intermediate was converted into triazole (16) under KOH/ethanol condition, followed by benzylation and hydrolysis to give the desired triazolone (18). For the synthesis of 1,3,4-oxadiazolone derivatives, aryl esters (13a–e) were converted into acylhydrazides (14a–e), which were cyclized using triphosgene and subsequently benzylated with substituted benzyl bromide to yield the final 1,3,4-oxadiazolone derivatives (20a–e).
Synthesis of Compounds 18, 20a-–e
To optimize the hit, first, the chain length between 2-chlorobenzene and 1,3,4-oxadiazolone was changed, and the results are summarized in Table. Unlike the hit compound 1, 2-chlorophenyl 8, and 2-chlorophenethyl 10 derivatives showed no significant antiseizure efficacy. Therefore, we optimized the benzyl moiety of 1. The unsubstituted benzyl derivative (12a) exhibited a weaker efficacy (85.8%) compared to compound 1 (53.3%). The 2,4-Dichloro derivative 12b showed improved efficacy, with 65.3%. Further, the 2-chloro-4-fluorobenzyl derivative (12c) showed further enhancement (40.4%) in scn1lab KO larvae compared to compound 1. To investigate the effect of chlorine at position 2 of the benzyl group, two compounds were designed and synthesized by substituting chlorine with hydrogen (12d) and fluorine (12e), which showed no efficacy in scn1lab KO larvae. The derivative with 4-chloro-2-fluorobenzyl (12f) was also examined for its activity but exhibited a weaker activity (63.0%) than that of 12c. Based on these results, the 2-chloro-4-fluorobenzyl moiety (12c) was identified as the most suitable structure for Part A, and subsequent optimization focused on modifications of the five-membered 1,3,4-oxadiazolone ring.
1: SAR Exploration of Compounds 1, 7, 10, 12a–f
The 1,2,4-oxadiazol-5(2H)-one structure (Part B) was modified, and the results are summarized in Table. The derivative with the 1,3,4-oxadiazolone moiety (20a) exhibited an enhanced antiseizure effect (19.7%) than 12c, whereas triazolone 18 exhibited no antiseizure effects (98.7%). These results indicated that among the five-membered heterocyclic core moieties including oxadiazolones (Part B), the 1,3,4-oxadiazolone scaffold was the most suitable structure for antiseizure activity, and subsequent optimization focused on modifying the aromatic ring.
2: SAR Exploration of Compounds 18 and 20a
The aromatic moiety (Part C) was optimized by the introduction of diverse aromatic groups and the results are summarized in Table. We found compound 20b, which replaced the thiophene scaffold with the same 5-membered furan, exhibited dramatically improved efficacy (2.4%); however, it showed a behavioral change in WT larvae (46.5%). Good antiseizure efficacies were observed in 6-membered pyridine derivatives such as the Pyridin-2-yl compound (20c) and the pyridin-4-yl compound (20d). Further, compound 20e, a 3-trifluoromethylpyridine-4-yl derivative, showed good antiseizure efficacy (19.5%) without significant differences in WT larvae. Therefore, we selected 20a and 20e for further evaluation. However, compound 20a showed poor stability in the liver microsomal assay, so compound 20e was chosen for further development.
3: SAR Exploration of Compounds 20a–e
Compound 20e was evaluated for its its blood-brain barrier (BBB), DMPK, and toxicity, and the results are presented in Table. The BBB serves as a gatekeeper of the central nervous system (CNS),? BBB penetration is essential for the efficacy of antiseizure medication in various biological models, including mouse, zebrafish, and human BBB chip systems. Compound 20e exhibited BBB-permeable properties; the brain-to-plasma (B/P) ratios were 4.36 ± 0.89, 1.75 ± 0.57, or 0.43 ± 0.12 in the mouse model, the adult zebrafish model, and the human BBB system, respectively (Tablea). Compound 20e has the good pharmacokinetic properties for an oral drug, with a reasonable AUC_0–24h_ (3.60 ± 1.34 μg·h/mL for the I.V. route and 3.83 ± 0.21 μg·h/mL for the P.O. route), rapid absorption (T max = 1.17 ± 0.76 h), and a moderate half-life with 3.83 ± 0.21 h following P.O. administration at a dose of 10 mg/kg in the mouse model. The bioavailability (F) of compound 20e was estimated to be 84.3% after P.O. administration (Tableb). In liver microsomal stability tests, compound 20e demonstrated reasonable stability during phase I metabolic reactions (Tablec). The percentages of compound remaining after 30 min of incubation were 69.2% in rat, 75.6% in dog, 79.0% in monkey, and 82.4% in human liver microsomes. Compound 20e exhibited excellent stability in plasma from various animals, including mice, rats, and humans. After a 4 h incubation period, the stability was greater than 99.9% in mouse plasma, 89.8% in rat plasma, and 89.9% in human plasma. Additionally, after a 4-h equilibrium state, the plasma protein binding rates of compound 20e were 98.3% in mouse, 98.0% in rat, and 99.2% in human plasma, respectively. Various toxicity studies (cytotoxicity, acute toxicity test, hERG, and AMES) were performed in in vitro and in vivo systems. To determine the cytotoxicity of compound 20e against mammalian cell lines, the cell viability was investigated by WST analysis. WST analysis was performed on mammalian cells (VERO, HFL-1, L929, NIH3T3, CHO-K1) after 24 h of treatment with varying concentrations of compound 20e (0.01, 0.1, 1, 10, and 100 μM). Compound 20e did not affect the normal cell viability. The potential of compound 20e to inhibit hERG was evaluated using the hERG FP (fluorescence polarization) assay. Compound 20e showed a low possibility of cardiac toxicity induced by inhibition of hERG channel activity with <1% inhibition at 10 μM.? A mini-Ames test was conducted with 20e using two strains of Salmonella typhimurium (TA98 and TA100) to assess its mutagenic potential. The results indicated that compound 20e exhibited no mutagenicity with or without the S9 fraction. Moreover, in an acute oral toxicity study, compound 20e exhibited no toxicological effects on body weight or organ morphology after the administration of 250 mg/kg to both male and female mice.
4: Biological Data of Compound 20e
Compound 20e treatment in scn1lab KO larvae effectively decreased seizure-like movements in a dose-dependent manner (Figures S1d,e and S2). We compared the antiseizure efficacy of 20e with FFA and CBD on scn1lab KO larvae. FFA treatment significantly reduced seizure-like movements in scn1lab KO larvae by 27.96% at 250 μM and 9.44% at 500 μM but also decreased normal movements in WT larvae. CBD treatment decreased seizure-like movements by 55.58% at 5 μM and 11.51% at 10 μM without affecting normal movements in WT larvae. Compound 20e treatment effectively reduced seizure-like movements by 41.58% at 2.5 μM and 16.3% at 5 μM without affecting normal movements in WT larvae. Compound 20e demonstrated the most effective reduction in seizure-like movements in scn1lab KO larvae. Additionally, unlike FFA, compound 20e did not affect normal movements in WT larvae, making it a particularly promising candidate for further study (Figure). 20e treatment also exhibited excellent antiseizure efficacy in the pentylenetetrazole (PTZ)-induced seizure zebrafish model (Figure S3). Electroencephalogram data showed that seizure-like events increased in KO larvae compared with the WT group. However, 20e treatment reduced both the number and duration of ictal-like events in the KO zebrafish larvae (Figure S1f). Prolonged or recurrent seizures in epilepsy patients, including those with SMEI, have been reported to cause cognitive impairment.? Because zebrafish have innate color discrimination capability, the cognitive ability of zebrafish larvae can also be evaluated through color preference testing.? WT or heterozygous sibling larvae tended to prefer blue over yellow, whereas scn1lab KO larvae showed the opposite tendency. However, compound 20e treatment in scn1lab KO larvae tended to restore the preference for blue (Figure S1g).
Antiseizure effects of compound 20e compared to FFA and CBD in scn1lab KO zebrafish larvae. A, B Quantification of normal movements in WT larvae and seizure-like movements in scn1lab KO larvae following treatment with FFA, CBD, or compound 20e. FFA reduced seizure-like movements at 250 μM (27.96%) and 500 μM (9.44%) but also decreased normal locomotion in WT larvae. CBD reduced seizure-like movements at 5 μM (55.58%) and 10 μM (11.51%) without affecting the WT movement. Compound 20e reduced seizure-like movements at 2.5 μM (41.58%) and 5 μM (16.3%) without altering normal locomotion in WT larvae. Compound 20e showed the most favorable efficacy–safety profile among the tested compounds.
To evaluate the in vivo efficacy, heat-induced seizure modeling was performed in SCN1A ^+/–^ mice. Generalized tonic-clonic seizures (GTCS) were not observed in animals treated with CBD or compound 20e (data not shown). Treatment with both CBD and compound 20e significantly mitigated seizure severity, assessed by the Racine scale score (Figurea). CBD administration reduced the mean score from 4.67 ± 0.21 (vehicle) to 4.00 ± 0.00 (a reduction of 0.67 points). Compound 20e demonstrated superior efficacy, reducing the score from 5.00 ± 0.00 (Vehicle) to 2.40 ± 0.60, achieving a substantial 2.60-point reduction and a 52% decrease in severity (Figurea). Furthermore, both the first seizure threshold temperature and the latency to the first seizure were significantly increased in the 20e-treated SCN1A ^+/–^ group (Figureb,c), indicating enhanced resistance to hyperthermia-induced and spontaneous seizures. In the open field test, SCN1A ^+/–^ mice exhibited significantly greater basal locomotor activity, including increased distance traveled and average speed, compared to their WT controls, consistent with previous findings.? The effect of CBD and 20e on hyperactivity track plots visually confirmed a reduction in the hyperactivity of mutant mice following treatment with either CBD (100 mg/kg) or 20e (5 mg/kg) (Figured). Quantitatively, both CBD and 20e treatments significantly reduced the total distance traveled (Figuree) and average speed (Figuref) in SCN1A ^+/–^ mice. Importantly, neither CBD nor 20e treatment affected the locomotor behavior of the WT control mice. These data collectively suggest that treatment with CBD or 20e successfully ameliorated the behavioral phenotypes associated with SCN1A haploinsufficiency by reducing the observed hyperactivity.
Compound 20e reduces seizure susceptibility and hyperactivity in SCN1A +/– mice. a Racine scale scores in heat-induced seizure assays. Treatment with CBD (100 mg/kg) or compound 20e (5 mg/kg) significantly reduced seizure severity, with 20e exhibiting a 52% reduction relative to vehicle controls. b, c Compound 20e significantly increased latency to first seizure and seizure threshold temperature, indicating enhanced resistance to hyperthermia-induced seizures. d–f Open field test showing hyperlocomotion in SCN1A +/– mice compared to WT. Both CBD and 20e treatments reduced the total distance traveled and average speed in SCN1A +/– mice without affecting WT behavior. Representative track plots illustrate reduced hyperactivity in treated mutant mice.
To investigate the functional effects of compound 20e, a cellular model of epilepsy was created using patient-derived iPSCs. A six-week differentiation period was required to achieve a functional, epilepsy-like phenotype. During this period, the iPSC-derived neurons showed a noticeable increase in the firing frequency over time. Spike and burst activity gradually rose, and network burst activity first appeared at week 5, becoming more pronounced by week 6. These developmental changes confirmed a hyperexcitable neuronal network suitable for drug evaluation (Figurea–d).
Electrophysiological characterization of compound 20e using microelectrode array. a−d Temporal changes in neuronal activity metrics during differentiation from weeks 3 to 6: a mean firing rate, b burst frequency, c network burst frequency, and d synchrony index. Data were expressed as mean ± SEM (n = 48). e–j Efficacy of compound 20e on electrophysiological metrics: e mean firing rate, f burst frequency, g burst percentage, h network burst frequency, i network burst duration, and j synchrony index.
The effects of compound 20e on the electrophysiological characteristics of the epileptic neuronal network were assessed in a dose-dependent manner. Concentrations up to 100 nM showed no significant change in the mean firing rate. However, a significant decrease was observed starting at 1 μM (Figuree). A similar pattern was seen for burst activity, with the burst index and burst percentage both showing a significant reduction at concentrations of 1 μM and above (Figuref,g). The drug’s impact on synchronized network activity was also evaluated. While no significant changes in network burst frequency or duration were seen at concentrations below 1 μM, both metrics were significantly and dose-dependently reduced at 10 and 50 μM (Figureh,i). Furthermore, the synchronization indexa key measure of collective network functionshowed a significant decrease at concentrations exceeding 1 μM (Figurej). Compound 20e effectively suppresses both individual neuronal firing and coordinated network-level activity in this patient-derived epilepsy model, with its inhibitory effects becoming significant at concentrations of 1 μM and above.
We investigated the mode of action of 20e. Neurotransmitter profiling has emerged? as a critical tool in elucidating CNS functions and uncovering related behavioral profiles across various animal models, including zebrafish. Mass spectrometry-based targeted analysis was used to determine the endogenous concentrations of neurotransmitters in zebrafish larvae. This study explored a broad spectrum of brain neurochemistry, including histaminergic, cholinergic, dopaminergic, serotonergic, and GABAergic systems, each closely associated with behavioral profiles in these models. Using this technology, significant changes induced by compound 20e were observed in the endogenous levels of neurotransmitters such as dopamine (DA), norepinephrine (NE), serotonin (5-HT), gamma-aminobutyric acid (GABA), glutamate (GLU), and glutamine (GLN) in zebrafish larvae. Notably, scn1lab KO zebrafish showed a decreased trend in 5-HT levels compared with WT counterparts. However, compound 20e exhibited significant neurological effects by increasing the levels of 5-HT and GABA in both WT and scn1lab KO zebrafish larvae as compared to the control group (Figurea). KO zebrafish also showed reduced 5-HT immunoreactivity in the hypothalamus; however, 5-HT levels were effectively restored following treatment with 20e (Figureb,c). These neurotransmitters exhibit sedative and antiseizure effects, potentially alleviating symptoms associated with SMEI.
Compound 20e modulates neurotransmitter levels in zebrafish larvae. a Quantification of endogenous neurotransmitters (DA, NE, 5-HT, GABA, GLU, and GLN) in WT and scn1lab KO zebrafish larvae following treatment with compound 20e. Treatment significantly increased 5-HT and GABA levels in both genotypes compared to control. KO larvae showed a reduction in basal 5-HT levels relative to WT. b, c Representative images and quantification of 5-HT immunoreactivity in the hypothalamus of scn1lab KO larvae. Reduced 5-HT immunoreactivity in KO was rescued by compound 20e treatment.
Tryptophan hydroxylase 2 (TPH2) is an enzyme variant of tryptophan hydroxylase that is found in vertebrates, and TPH2 plays an important role in 5-HT synthesis in the brain. In humans, TPH2 is predominantly expressed in the serotonergic neurons within the brain, with the highest levels of expression observed in the raphe nucleus located in the midbrain.? In zebrafish, the genome contains three tph gene paralogs, but among these, only tph2 is expressed specifically in the raphe nucleus.? Compound 20e treatment increased the endogenous 5-HT level in both WT and scn1lab KO larvae. Based on this result, quantitative RT-PCR was performed to analyze the expression of tph2. As a result, compound 20e treatment in scn1lab KO larvae increases the expression of tph2, whereas it is not altered in WT larvae (Figurea). Furthermore, TPH2 expression measured by real-time PCR showed no significant difference between the wild type and mutant type in both mouse and cerebral organoid models. However, upon 20e treatment, only the SCN1A ^+/–^ group exhibited a significant upregulation of TPH2 expression at mRNA levels (Figureb,c). This effect was consistent across in vivo and in vitro systems. These results suggest a mutation-specific sensitivity of the serotonergic pathway to pharmacological modulation.
Compound 20e selectively upregulatesTPH2 expression inSCN1A -deficient models. a, Quantitative PCR analysis of tph2 expression in WT and scn1lab KO zebrafish larvae following 20e treatment. 20e significantly increased tph2 expression in KO larvae. b, c The tph2 mRNA levels in mouse brain and cerebral organoids showed no baseline difference between WT and SCN1A +/– groups. Upon 20e treatment, tph2 expression was significantly upregulated only in SCN1A +/– models. These findings indicate a mutant-dependent serotonergic response to 20e across both in vivo and in vitro systems.
Discussion
In this study, we aimed to identify a novel drug candidate for SMEI. We successfully established scn1lab KO zebrafish through a 4-bp insertion via the CRISPR/Cas9 method. Unlike the missense mutant didy ^ s552 ^ zebrafish, kri111 zebrafish produces the Scn1lab protein, which is truncated by a premature stop. However, the morphological and behavioral phenotypes of the two mutants are similar. By measuring only high-speed movements above 80 mm/s, we can effectively assess hyperactivity and loss-of-posture, and scn1lab KO zebrafish showed a marked increase in movement compared with WT at these speeds. Based on this scn1lab KO zebrafish model, we conducted phenotype-based screening and identified a novel hit compound with an oxadiazolone scaffold. In a subsequent SAR study, we identified compound 20e, which showed outstanding antiseizure efficacy without significant differences in WT larvae. In an in vivo BBB penetration test, 20e demonstrated B/P ratios of 4.36 ± 0.89 in the mice and 1.75 ± 0.57 in the adult zebrafish, suggesting good BBB penetration and its effective antiseizure potential. Furthermore, the bioavailability of 20e was estimated to be 84.3% after oral administration. 20e exhibited good liver microsomal stability and no significant inhibition of hERG, did not show mutagenic potential in the AMES test, and had an LD_50_ value greater than 250 mg/kg in the acute toxicity study. Electroencephalogram analysis is a widely considered tool to diagnose epileptic seizures.? Recently, noninvasive multichannel electroencephalographic recording in larval zebrafish became available for evaluating seizure-like events.? Treatment of 20e decreased both the number and duration of ictal-like events in the KO zebrafish larvae. Furthermore, 20e demonstrated antiseizure effects at lower concentrations than FFA and CBD, while showing no significant differences in WT larvae.
Oral administration of compound 20e significantly reduced the seizure severity and prevented GTCS in SCN1A ^+/–^ mice. It also delayed seizure onset and increased seizure threshold temperature, indicating enhanced resistance to heat-induced seizures. These effects suggest that 20e modulates neuronal excitability relevant to SMEI pathology. Additionally, 20e reduced hyperactivity in SCN1A ^+/–^ mice without affecting the WT behavior, indicating mutation-specific behavioral rescue. Neurotransmitter profiling was conducted and the endogenous concentrations of neurotransmitters were determined using mass spectrometry. The application of mass spectrometry-based neurotransmitter profiling has proven beneficial in biomarker discovery and the optimization of drug candidates for CNS research.? 20e showed notable neurological changes by elevating the levels of 5-HT and GABA in both WT and scn1lab KO zebrafish larvae. This study underscores the importance of neurochemical research in advancing our understanding of CNS disorders and refining therapeutic strategies. Additionally, the qRT-PCR results showed that TPH2, which plays a key role in 5-HT synthesis in the brain, exhibited a dose-dependent increase in expression upon 20e treatment, suggesting that 20e increases 5-HT levels through TPH2. By enhancing endogenous 5-HT production rather than directly stimulating receptors, 20e may provide more physiological regulation of serotonergic tone while reducing the risks of receptor desensitization or overstimulation associated with chronic agonist exposure. Critically, this TPH2 upregulation occurred specifically in SCN1A-deficient modelsno significant TPH2 upregulation was observed in WT zebrafish, WT mice, or control organoids, whereas significant dose-dependent TPH2 upregulation was found only in scn1lab KO zebrafish, SCN1A ^+/–^ mice, and mutant cerebral organoids. Furthermore, we confirmed that the antiseizure effect of 20e was significantly attenuated when coadministered with telotristat, a TPH1/2 inhibitor, in the zebrafish Dravet syndrome model (Figure S6). In addition, 20e treatment was associated with enhanced GABAergic function, as evidenced by increased GABA levels and normalization of electrophysiological network activity, including reduced mean firing rate and burst frequency in GABAergic neuronal networks. These GABAergic changes are consistent with the stabilization of abnormally disrupted network activity in SCN1A-deficient systems. Taken together, this mutant-specific serotonergic restoration accompanied by GABAergic network stabilization provides a mechanistic explanation for the selective suppression of seizure-related behaviors observed in our studies, wherein 20e suppresses seizures in disease models without affecting normal behavior in WT animals. Additionally, scn1lab KO zebrafish larvae displayed elevated levels of stress markers, cortisol and cortisone,? compared to WT zebrafish. These levels were restored to normal by compound 20e (Figure S4). Expression of inflammatory markers, interleukin 1β (il1β) and tumor necrosis factor α (tnfα), was increased in KO zebrafish larvae at 9 dpf. Interestingly, while the level of expression of il1β was significantly reduced following 20e treatment, the level of expression of tnfα was upregulated in KO zebrafish larvae (Figure S5). This suggests that the 20e does not act as a broad-spectrum anti-inflammation but rather exerts selective immunomodulatory effects. The combined data suggest that compound 20e could exhibit antiseizure efficacy by modulating these endogenous substances. This modulation could potentially reduce the seizure behavior observed in SMEI models. Therefore, our findings propose that the endogenous alterations induced by compound 20e, including 5-HT, GABA, cortisol, and cortisone, could serve as therapeutic targets in biological systems.
We demonstrate that compound 20e exerts a potent, dose-dependent inhibitory effect on neuronal hyperactivity in patient-derived iPSC-based neuronal networks, with significant reductions in spike and burst activity particularly evident at concentrations above 1 μM. These effects were more pronounced in SCN1A^+/–^ models, indicating that 20e preferentially targets hyperexcitable circuits relevant to SMEI. In addition to reducing overall neuronal firing, 20e also disrupted network-level synchronization, a hallmark of epileptiform activity. Although variability in burst duration and frequency was observed, likely reflecting the complexity of neural network regulation, the consistent suppression of hyperactivity highlights the therapeutic potential of compound 20e.
Conclusion
We established Nav1.1 (scn1lab) KO zebrafish to conduct chemical screening and identified novel 1,3,4-oxadiazol-2(3H)-one derivatives for SMEI treatment. In the KO zebrafish model, compound 20e showed significant antiseizure efficacies in behavioral analysis, which are better than those of marketed drugs. Compound 20e exhibited favorable in vivo pharmacokinetic (PK) properties for oral administration with reasonable BBB penetration in mice. Safety evaluation indicated no significant hERG inhibition, cytotoxicity, mutagenic potential, and acute toxicity. Compound 20e efficiently reduced the total distance moved and the mean velocity in the SCN1A ^+/–^ mice model of SMEI. It improved pathological spike and burst activity in severe myoclonic epilepsy of infancy (SMEI) patient-derived neurons. For the mode of action, compound 20e increased the concentration of 5-HT through KO-specific upregulation of the TPH2 gene, representing a mechanistically distinct approach from fenfluramine’s direct receptor activation. The multimechanistic profile encompassing serotonergic, GABAergic, anti-inflammatory, and network modulatory effects, combined with superior potency and KO-specific selectivity, distinguishes 20e as a novel therapeutic candidate. For mode of action, compound 20e increased the concentration of 5-HT through the increase of the TPH2 gene. Taken together, these results underscore the therapeutic potential of compound 20e as a promising candidate for the treatment of SMEI.
Experimental Section
General
All solvents and chemicals were used as purchased without further purification. All reported yields are isolated after column chromatography or crystallization. ^1^H and ^13^C NMR spectra were recorded on JEOL JNM-ECS400 spectrometers at 400 MHz for ^1^H NMR and 100 MHz for ^13^C NMR, respectively. Chemical shifts (δ) are expressed in parts per million relative to tetramethylsilane as an internal standard, and CDCl_3_, DMSO-d 6, and CD_3_OD were used as solvents. The multiplicity of the peak is expressed by s (singlet), d (doublet), t (triplet), q (quartet), dd (doublet of doublets), td (triplet of doublets), qd (quartet of doublets), dt (doublet of triplets), and m (multiplet). HRMS data were obtained by Impact II (Bruker, USA). HPLC analyses were performed with a Waters Agilent HPLC system equipped with a PDA detector and an Agilent SB-C18 column (1.8 μm and 2.1 × 50 mm). The mobile phase consisting of buffer A (ultrapure H_2_O containing 0.1% trifluoroacetic acid) and buffer B (chromatographic-grade CH_3_CN) was applied at a flow rate of 0.3 mL min^–1^. All compounds are >95% pure by HPLC analysis.
Synthesis of Compound 20e
3-(2-Chloro-4-fluorobenzyl)-5-(2-(trifluoromethyl)pyridin-4-yl)-1,3,4-oxadiazol-2(3H)-one (20e)
Step 1. Ethyl 2-trifluoromethylpyridine-4-carboxylate 13e as an aryl ester (500 mg, 2.283 mmol) was dissolved in ethanol. Hydrazine hydrate (457.18 mg, 9.132 mmol) was added in the reaction mixture. The mixture was heated for reflux and stirred for 6 h. The solvent was removed under reduced pressure. The residue was diluted by water and extracted with ethyl acetate. The organic phase was dried over anhydrous sodium sulfate and concentrated in vacuo to acquire 2-(Trifluoromethyl)-4-pyridinecarboxylic acid hydrazide 14e (413 mg, 2.015 mmol, 88%). ^1^H NMR (400 MHz, DMSO-d 6) δ 10.35 (s, 1H), 8.92 (d, J = 4.9 Hz, 1H), 8.21 (s, 1H), 8.06 (dd, J = 4.9, 0.9 Hz, 1H), 4.64–4.82 (2H).
Step 2. 2-(Trifluoromethyl)-4-pyridinecarboxylic acid hydrazide 14e (413 mg, 2.015 mmol) and 2 equiv of diethylpropylamine (0.702 mL, 4.029 mmol) were dissolved in THF (10 mL) at 0 °C. Triphosgene (239.74 mg, 0.806 mmol) dissolved in THF was added slowly dropwise. After the mixture was stirred for 1 h at 0 °C, the reaction mixture was allowed to reach room temperature and heated to reflux for 6 h. After cooling to room temperature, the solvent was removed under reduced pressure. The residue was purified by silica gel column chromatography to give 5-(2-(trifluoromethyl)pyridin-4-yl)-1,3,4-oxadiazol-2(3H)-one 19e (315 mg, 1.364 mmol, 68%). ^1^H NMR (400 MHz, DMSO-d 6) δ: 13.11 (s, 1H), 8.96 (d, J = 5.2 Hz, 1H), 8.09 (s, 1H), 8.06 (d, J = 4.9 Hz, 1H).
Step 3. 5-(Pyridin-4-yl)-1,3,4-oxadiazol-2(3H)-one 19e (378.4 mg, 2.318 mmol) and 2-chloro-4-fluorobenzyl bromide (570.17 mg, 2.551 mmol) were dissolved in DMF (2 mL) under N_2_ condition. Sodium hydride 60% dispersion in mineral oil (101.87 mg, 2.551 mmol) in DMF was added slowly, dropwise, to the solution. The solution was heated to 60 °C and stirred for 4 h. The solvent was removed under reduced pressure. The residue was diluted with water (20 mL) and extracted with ethyl acetate (3 × 50 mL). The combined organic layer was washed with brine (20 mL). The crude product was purified by silica gel column chromatography to give 3-(2-chloro-4-fluorobenzyl)-5-(pyridin-4-yl)-1,3,4-oxadiazol-2(3H)-one 20e (463.7 mg, 1.517 mmol, 65%). ^1^H NMR (400 MHz, DMSO-d 6) δ 8.96 (d, J = 4.6 Hz, 1H), 8.05 (d, J = 5.5 Hz, 2H), 7.62 (dd, J = 8.4, 6.3 Hz, 1H), 7.55 (dd, J = 8.9, 2.4 Hz, 1H), 7.28 (td, J = 8.4, 2.2 Hz, 1H), 5.09 (s, 2H); ^13^C NMR (101 MHz, DMSO-d 6) δ 164.0, 161.5, 153.2, 152.7, 151.2, 149.0, 148.6, 148.3, 147.9, 134.5, 134.4, 133.8, 133.4, 133.3, 129.6, 129.6, 126.2, 123.6, 123.5, 120.7, 118.1, 117.8, 116.8, 115.8, 115.6, 47.6; HRMS (ESI) m/z calculated for C_15_H_9_ClF_4_N_3_O_2_ [M + H]^+^ 374.03139, found 374.03036; HPLC purity 98.1616%.
Biological Experiments
Experiments of SMEI Model in Zebrafish Larvae
Larvae clutches were bred from scn1lab ^ kri111/+ ^. Homozygous scn1lab KO larvae showing hyperpigmentation and age-matched sibling larvae were used. 6-dpf larvae were placed individually into a 96-well plate. The microplate was placed inside a behavioral tracking device and acclimated to the dark condition for 30 min. DMSO or compounds were added into each well. The behavioral data of each larva were recorded using EthoVision 15 software (Noldus) connected to the DanioVision chamber (Noldus) for 30 min. Seizure-like movements were defined as previously described. Immunohistochemistry for 5-HT was conducted as previously described.? Larvae were placed into a 6-well plate with a cell strainer (SPL, 93100) to ensure rapid fixation. After treatment for 4 h, each cell strainer was immediately transferred into 4% paraformaldehyde/4% sucrose in PBS at 4 °C. After overnight fixation, larvae were washed with 0.25% Triton X-100/PBS (PBTx). Samples were used to dissect the brain from larvae to highly penetrate antibody. Dissected brains were incubated in 1 mg/mL collagenase (Sigma-Aldrich, C9891) for 1 h and blocked overnight in 2% normal goat serum/2% DMSO in PBTx at 4 °C. Then, dissected brains were incubated overnight in blocking solution with antibodies at 4 °C. Rabbit 5-HT antibody (Sigma-Aldrich, S5545) and goat anti-Rabbit IgG (Invitrogen, A-11034) were used as the primary and secondary antibodies, respectively. After some washing with PBTx, dissected brains were mounted in 1% low-melting agarose and imaged using a K1-Fluo confocal fluorescence laser scanning microscope (Nanoscope Systems).
Neurotransmitter and Neurosteroid Analysis
To investigate brain-specific metabolites, including neurotransmitters and neurosteroids, in zebrafish larvae, we performed targeted analysis using LC-MS/MS, according to our previously reported methods.? Briefly, 30 pooled zebrafish larvae at the 6 dpf stage were exposed to 5 μM 20e for 4 h in a 6-well plate, then washed and collected into a 1.7 mL tube. For neurotransmitter analysis, wet larvae were snap-frozen using liquid nitrogen, and 300 μL of distilled water (DW) was added, followed by homogenization using a sonicator in a 1.7 mL tube on ice. The homogenates were mixed with methanol containing 1% formic acid. Endogenous neurotransmitters were extracted by vortexing for 5 min, and the clear supernatant was transferred into LC vials after centrifugation at 15,000 rpm for 10 min. For neurosteroid analysis, 1 mL of methanol/acetic acid (99:1 v/v) was used to homogenize the samples. The homogenate was then centrifuged for 5 min at 12,000 rpm, and the remaining pellet was extracted twice with methanol/acetic acid (99:1 v/v). The pellet was subsequently resuspended in 1 mL of methanol/water (10:90 v/v), and the mixture was loaded onto a solid-phase extraction cartridge (Oasis PRIME HLB, Waters). After eluting twice with 1 mL of methanol, the sample was evaporated under nitrogen gas. Final eluates were transferred to an LC vial. Neurotransmitters and neurosteroids were quantitatively analyzed using LC-MS/MS.
Experiments of SMEI Model in Mouse
Scn1a ^ tm1Kea ^ mice on a pure 129S6/SvEvTac (129) inbred strain background were generated by homologous recombination in TL1 ES cells (129S6/SvEvTac). The mouse line Scn1a^tm1Kea^ has been maintained as a coisogenic strain by continuous backcrossing of null heterozygotes to 129 (129.Scn1a+/–. 129S-Scn1a^tm1Kea/Mmjax^ (#024761) heterozygous mice and WT animals were obtained from Jackson Laboratory, USA, and male 129S-Scn1atm1Kea/Mmjax heterozygous mice (Jackson Laboratory, USA) were crossed with female WT C57BL/6 animals. Thus, Scn1a+/– and WT littermate mice were obtained. These lines were maintained, bred together, and used in this study (n = 5 to 7 per group). All experiments were approved by the Institutional Animal Care and Use Committee at the Korea Research Institute of Chemical Technology and were conducted in accordance with the guidelines of the Ministry of Food and Drug Safety for the care and use of laboratory animals, as well as the policy on humane care and use of laboratory animals (approval number: 2024-7A-09-01).
The mice were housed in a specific pathogen-free (SPF) barrier facility with a 12 h light/12 h dark cycle and had ad libitum access to food and water. The genotype of the mice was confirmed by PCR, ensuring the successful generation and maintenance of the Scn1a mutant mouse model. CBD was purchased from Sigma. CBD was dissolved in a 1:1:18 ratio in 100% ethanol, cremophor, and 0.9% saline, respectively.? 20e was prepared in DMSO:PEG400:water = 10:60:30 (v/v) for acute administration. This is because it is a good general-purpose solvent and maintains excipient consistency from test to test. All drugs were administered at a dose of 10 mL/kg. All drug compounds were dosed and tested based on their previously determined time-to-peak effect in the maximal seizure.? Seizures were induced as previously described? with some modifications. Briefly, mice were placed in a Plexiglas cylinder with an infrared heating lamp (250 W, HL-1, Physitemp Instruments Inc.) held in a fixed position. A rectal temperature probe (RET-4, Physitemp Instruments Inc.) was carefully inserted and taped to the mouse’s tail. The animal’s core body temperature was monitored by connecting a temperature probe to a temperature controller (TCAT-2DF, Physitemp Instruments Inc.). In this study, male and female mice aged 13 to 15 weeks were administered an intraperitoneal injection of 100 mg/kg of CBD 1 h prior to heat induction. Additionally, 30 min before the heat exposure, they received a gavage dose of 5 mg/kg of 20e. The temperature of the rectal probe inserted into the mouse was adjusted at 1 °C intervals to 37.5 °C, 38.5 °C, 39.5 °C, 40.5 °C, 41.5 °C, 42.5 °C, and 43.5 °C. The temperature was observed and recorded for approximately 2 min. The effects of each drug were confirmed through a t-test by comparing the seizure time and temperature. The overall Modified Racine’s Scale (seizure severity)? reached levels 3 (first seizure) and 5 (GTCS; tonic-clonic seizures). Scale scores were evaluated as significant. In particular, because the Racine Scale cannot reliably assess seizures with severity <3 in mice, we limited the assessment of seizure severity to those with a Racine score of 3–5. Each mouse was placed near the lower left wall of a 40 × 40 cm open arena. Mice were placed in an apparatus and allowed to freely explore for 15 min. Mouse movements were analyzed with a GigE camera with a lens and PC-based video tracking software (EthoVision XT 17, Noldus Technology). Based on the data profile, we analyzed the results for total travel distance and speed.
Neuron Culture
Neuron culture began on Day 1, defined as the day doxycycline was added. The cells were cultured in NGM Media, replaced daily, for about 1 week. The day before reaching 1 week, astrocytes (1 × 10^5^ cells) were seeded onto Matrigel-coated coverslips in 24-well plates. Neurons were dissociated into single cells by using Accutase. A portion (5 × 10^4^ cells) was used for electrophysiology measurements, while another (5 × 10^4^ cells) was cultured in NGM Media supplemented with 10% Mouse Astrocyte Conditioned Medium (ScienCell Research Laboratories, Carlsbad, California, United States) for ICC and cDNA synthesis. For microelectrode array (MEA) recordings, astrocytes were seeded onto Matrigel-coated coverslips (5 × 10^4^ cells), followed by the addition of 1.5 × 10^4^ GABAergic and 3.5 × 10^4^ glutamatergic neurons, making a total of 1 × 10^5^ cells. Media changes were performed every 2 days, with 50% of the media refreshed during each change.
MEA Measurement and Data Analysis
MEA recordings were conducted using a 24-well Maestro Edge system (Axion BioSystems, Atlanta, Georgia, United States). Each MEA well contained a total of 1 × 10^5^ cells, comprising 1.5 × 10^4^ GABAergic neurons, 3.5 × 10^4^ glutamatergic neurons, and 5 × 10^4^ astrocytes. Neural network activity from iPSC-derived neurons and their calibrated counterparts was recorded for 15 min at 37 °C in a chamber maintained with 95% O_2_ and 5% CO_2_. The recordings were sampled at 10 kHz, filtered with a 100 Hz high-pass and a 3500 Hz low-pass filter. Spikes were detected at ±4.5 standard deviations. Metrics such as mean firing rate were calculated as the average spike frequency across all channels in a well. Burst activity was defined as channels exhibiting at least five spikes per burst, with a minimum interburst interval of 100 ms and a threshold of 0.4 bursts/s. Network bursts were characterized as synchronized bursts across more than 35% of channels in a well. Data analysis was performed using Axion software (AxIS) following the manufacturer’s guidelines. Patient-derived neurons were analyzed from 3 weeks to 6 weeks. Drug treatment was administered at 6 weeks, followed by measurements. Each drug was applied in a stepwise manner, starting from a low concentration and gradually increasing to higher concentrations. After drug application, a reaction time of 10 min was allowed, and spontaneous neuronal activity was recorded for 15 min.
Experiments of Cerebral Organoid
Induced pluripotent stem cells (iPSCs, IMR90-4, WiCell, Madison, USA) served as the initial seeding material for the culture of cerebral organoids. These cells were seeded into U-bottom ultralow-attachment 96-well plates (Corning). The neural induction medium was composed of Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM/F12) supplemented with 1% GlutaMAX, 1% MEM-NEAA (Minimum Essential Medium Non-Essential Amino Acids), 15% knockout serum (Thermo Scientific), 0.1 nM β-mercaptoethanol, 100 nM LDN-193189, 10 μM SB431542, and 2 μM XAV939 (Sigma-Aldrich, St. Louis, MO, USA). The cells underwent static culture conditions for 10 days, with media changes performed every other day. On day 10, the cells were transferred to ultralow-attachment 6-well plates (Corning) to initiate neural differentiation. The neural differentiation medium (NDMI) was formulated by combining DMEM/F12 and neurobasal medium in a 1:1 ratio. To this mixture, an N2 supplement, a B27 supplement without vitamin A (both from Invitrogen), 1% MEM-NEAA, 1% GlutaMAX (Thermo Scientific), and human insulin solution (Sigma-Aldrich) were added. Subsequently, the cells were cultured on an orbital shaker at 80 rpm for 8 days. From day 18 onward, the cells were transitioned to neural differentiation medium (NDMII) supplemented with B27 with vitamin A (Thermo Scientific), brain-derived neurotrophic factor, cyclic adenosine monophosphate, and ascorbic acid (Sigma-Aldrich) to facilitate neural maturation. The medium was refreshed every 4 days to sustain the maturation process of the cerebral organoids. The culture period spanned 120 days, during which the cells underwent successive stages of neural induction, neural differentiation, and neural maturation, culminating in the development of mature cerebral organoids.
Quantitative Real-Time PCR
Total RNA was isolated from zebrafish larvae (20 pooled) after exposure of 20e in WT or homozygous scn1lab KO using TRIzol reagent (Invitrogen, catalog no. 15596026) and purified according to manufacturer’s protocol. qRT-PCR was performed using Verso SYBR Green 1-Step qRT-PCR Low ROX Mix (Thermo Scientific, cat# AB-4106/A). PCR cycling conditions were as follows: cDNA synthesis at 50 °C for 15 min, initial denaturation at 95 °C for 15 min, followed by 40 cycles of 95 °C for 15 s, 60 °C for 30 s, and 72 °C for 30 s. Total RNA was extracted from frozen mouse brain tissues and human cerebral organoids using the RNeasy Plus Mini Kit (Qiagen, Valencia, CA, USA), following the manufacturer’s instructions. RNA concentration and purity were assessed using a NanoDrop One spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Subsequently, 2 μg of total RNA from each sample was reverse transcribed into complementary DNA (cDNA) using the AccuPower RT PreMix (Bioneer Inc., Seoul, Korea). Quantitative gene expression analysis was performed using gene-specific primers for human TPH2 and mouse Tph2. qRT-PCR was conducted using the Rotor-Gene Q system (Corbett Research, Mortlake, Australia) with the QuantiTect SYBR Green PCR Kit (Qiagen). Each 20 μL PCR reaction contained 10 μL of SYBR Green master mix, 2 μL of 10 pmol/μL forward and reverse primers, and 1 μL of cDNA template. PCR cycling conditions were as follows: initial denaturation at 95 °C for 15 min, followed by 40 cycles of 95 °C for 30 s, 60 °C for 30 s, and 72 °C for 30 s. Relative mRNA expression levels were calculated using the ΔCt method, where ΔCt = Ct (target gene) – Ct (endogenous gene). All data were normalized to β-actin, 18S rRNA expression to account for variability in input RNA and transcriptional activity. Oligos and primer sets are listed in Table S1.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wirrell E. C.Laux L.Donner E.Jette N.Knupp K.Meskis M. A.Miller I.Sullivan J.Welborn M.Berg A. T.Optimizing the diagnosis and management of Dravet syndrome: recommendations from a North American consensus panel Pediatr. Neurol 2017681834.e 310.1016/j.pediatrneurol.2017.01.02528284397 · doi ↗ · pubmed ↗
- 2a Sourbron J.Partoens M.Scheldeman C.Zhang Y.Lagae L.de Witte P.Drug repurposing for Dravet syndrome in scn 1Lab–/– mutant zebrafish Epilepsia 2019602 e 8–e 1310.1111/epi.1464730663052 PMC 6850687 · doi ↗ · pubmed ↗
- 3Strzelczyk A.Schubert-Bast S.A practical guide to the treatment of Dravet syndrome with anti-seizure medication CNS Drugs 202236321723710.1007/s 40263-022-00898-135156171 PMC 8927048 · doi ↗ · pubmed ↗
- 4Brenot F.Herve P.Petitpretz P.Parent F.Duroux P.Simonneau G.Primary pulmonary hypertension and fenfluramine use Heart 199370653754110.1136/hrt.70.6.537PMC 10253858280518 · doi ↗ · pubmed ↗
- 5a Villas N.Meskis M. A.Goodliffe S.Dravet syndrome: characteristics, comorbidities, and caregiver concerns Epilepsy Behav.201774818610.1016/j.yebeh.2017.06.03128732259 · doi ↗ · pubmed ↗
- 6a Meisler M. H.Kearney J. A.Sodium channel mutations in epilepsy and other neurological disorders J. Clin. Invest.200511582010201710.1172/JCI 2546616075041 PMC 1180547 · doi ↗ · pubmed ↗
- 7Baraban S. C.Dinday M. T.Hortopan G. A.Drug screening in Scn 1a zebrafish mutant identifies clemizole as a potential Dravet syndrome treatment Nat. Commun.201341241010.1038/ncomms 341024002024 PMC 3891590 · doi ↗ · pubmed ↗
- 8Hwang K.-S.Kan H.Kim S. S.Chae J. S.Yang J. Y.Shin D.-S.Ahn S. H.Ahn J. H.Cho J.-H.Jang I.-S.Efficacy and pharmacokinetics evaluation of 4-(2-chloro-4-fluorobenzyl)-3-(2-thienyl)-1, 2, 4-oxadiazol-5 (4H)-one (GM-90432) as an anti-seizure agent Neurochem. Int.202014110487010.1016/j.neuint.2020.10487033035603 · doi ↗ · pubmed ↗