Identification and Evaluation of Benzimidazole- Agonists of Innate Immune Receptor NOD2
Liora Wittle, Karl L. Ocius, Mahendra D. Chordia, Carly van Wagoner, Timothy N. J. Bullock, Marcos M. Pires

TL;DR
This paper identifies benzimidazole compounds, including nocodazole, as new NOD2 agonists that could improve immune-related therapies.
Contribution
The discovery of benzimidazoles as a novel class of NOD2 agonists with potential therapeutic advantages over existing MDP-based agonists.
Findings
Benzimidazoles, including nocodazole, activate NOD2 with nanomolar potency.
Nocodazole induces cytokine release similar to MDP-based NOD2 activation.
Benzimidazole agonists may offer better pharmacological properties than current MDP derivatives.
Abstract
Emerging evidence has demonstrated the importance of pattern recognition receptors (PRRs), including the nucleotide-binding and oligomerization domain receptor 2 (NOD2), in human health and disease states. NOD2 activation has shown promise with aiding malnutrition recovery, lessening irritable bowel disease (IBD) symptoms, and increasing the efficacy of cancer immunotherapy. Currently, most NOD2 agonists are derivatives or analogs of the endogenous agonist derived from bacterial peptidoglycan, muramyl dipeptide (MDP). These MDP-based agonists can suffer from low oral bioavailability and cause significant adverse side effects. With the goal of broadly improving NOD2 therapeutic interventions, we sought to discover a small molecule capable of activating NOD2 by screening a library of total 1917 FDA approved drugs in a phenotypic assay. We identified a class of compounds, benzimidazoles,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 2
2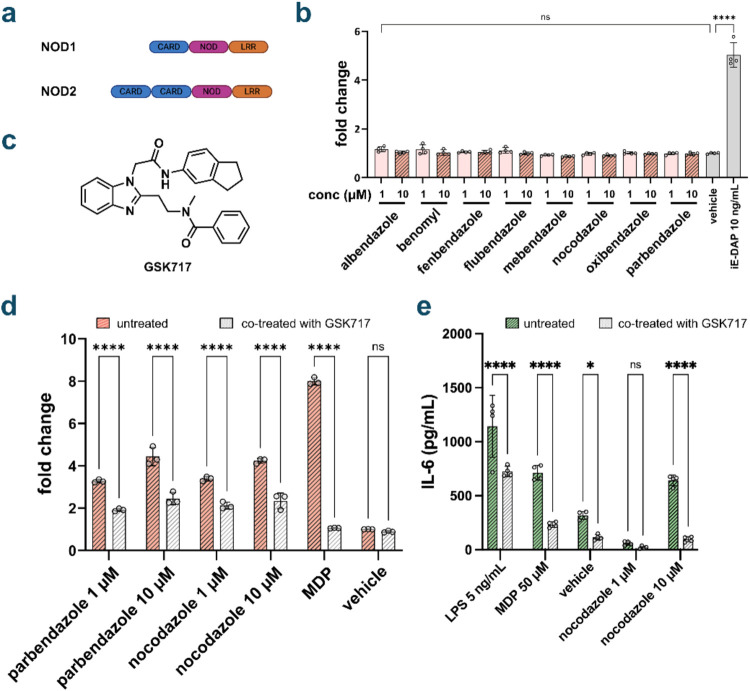 3
3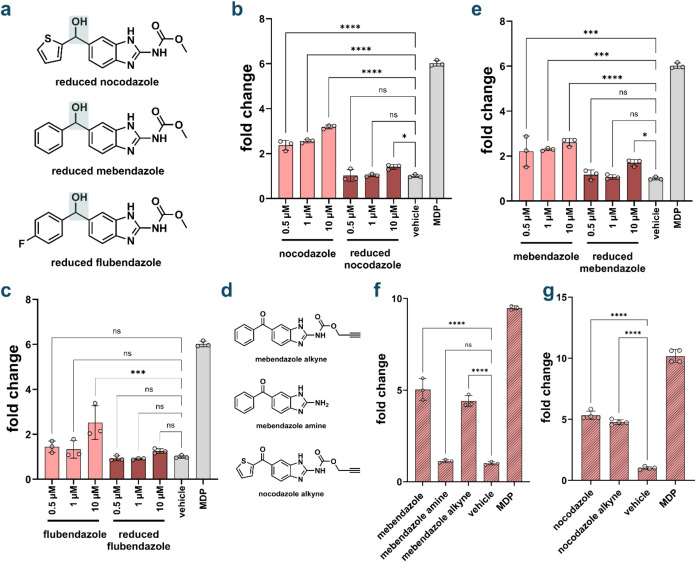 4
4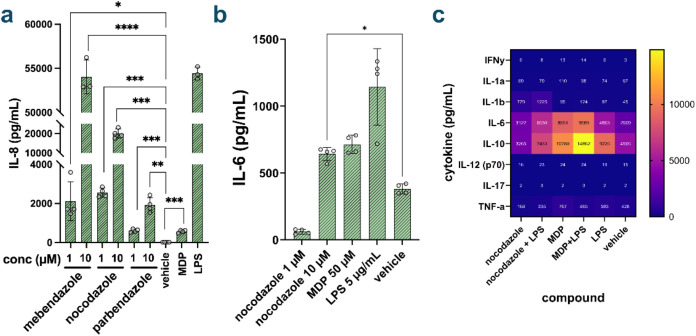 5
5- —National Institute of General Medical Sciences10.13039/100000057
- —National Institute of Allergy and Infectious Diseases10.13039/100000060
- —National Institute of Allergy and Infectious Diseases10.13039/100000060
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmune Response and Inflammation · Helicobacter pylori-related gastroenterology studies · Immune Cell Function and Interaction
Nucleotide binding and oligomerization domain receptor 2 (NOD2) is a pattern recognition receptor (PRR) expressed mainly on immune and gut epithelial cells. ?−? ? ? ? PRRs detect pathogen associated molecular patterns (PAMPs) and activate the immune system against potential infections.? PRRs are increasingly targeted for treating or preventing infections and inflammatory diseases. ?,?,? NOD2–a cytosolic, membrane associated receptor–detects fragments of peptidoglycan (PG), a primary component of bacterial cell membranes expressed by all known species of bacteria, including those associated with the gut microbiome. ?,?,? The structure of PG consists of cross-linked units of N-acetyl glucosamine (GlcNAc) and N-acetyl muramic acid (MurNAc), with a short peptide chain conjugated to MurNAc.? Muramyl dipeptide (MDP), the minimal PG fragment capable of activating NOD2, does so at nanomolar concentrations.? Upon activation, NOD2 initiates signaling through receptor interacting serine/threonine kinase 2 (RIPK2), leading to the activation of nuclear factor kappa β (NF-kB) (Figurea). NF-kB activation, in turn, causes the transcription of several cytokines and other immune signaling molecules, such as interleukin (IL)-6, IL-8, IL-10, and tumor necrosis factor α (TNF-α) that can trigger and modulate inflammatory responses. ?,?,? Despite the promising features of MDP, such as its potency in activating NOD2, early clinical evaluations revealed it to be pyrogenic and to have poor oral bioavailability.?
NOD2 activation has been shown to improve recovery from malnutrition,? regulate appetite,? protect against bacterial infections, ?,? reduce inflammatory bowel disease (IBD) symptoms, ?,? and enhance efficacy of PD-1/PD-L1 cancer immunotherapy (Figureb). ?−? ? ? One promising therapeutic strategy involves the use of NOD2 agonists as adjuvants in cancer checkpoint therapy. Programmed cell death protein 1 (PD-1) is a receptor, widely expressed on immune cells, that recognizes programmed cell death ligand 1 (PD-L1) and subsequently suppresses immune activity.? Many tumors upregulate PD-L1 to evade immune surveillance by preventing immune cells from recognizing and eliminating cancer cells.? To date, clinical responses to checkpoint therapy have been highly variable across cancer patients.? Notably, stronger responses to PD-1/PD-L1 immunotherapy have been associated with higher levels of NOD2 activation. ?−? ? ?,?,? Muramyl tripeptide (MTP) nanoparticles that activate NOD2 have been shown to assist in treating melanoma, especially in concert with immune checkpoint therapies.? In mice, treatment with a probiotic bacterium engineered to express a PG hydrolase enhanced the efficacy of PD-L1 immunotherapy, and this effect was shown to be dependent on NOD2 activation.?
Many of the beneficial effects from NOD2 activation have been demonstrated using probiotics or bacterial-derived molecules (e.g., isolated complete PG). For example, strains that release PG-digesting hydrolases in the gut have been shown to improve NOD2 activation by promoting the depolymerization of PG into NOD2-activating fragments. ?,? Treatment with bacteria that secrete PG hydrolases, or with isolated hydrolases themselves, has been shown to alleviate IBD symptoms and protect against bacterial infection. ?,?,?
While the use of probiotic bacteria to modulate the gut microbiome is promising, it presents several challenges. Probiotics can be difficult to permanently introduce into the host, and if harbored within the gut, their growth rates and release of PG fragments can be challenging to control. Sustaining their residence in the gut is also problematic during dysbiosis, and interactions with the existing microbial community can further complicate outcomes.? In immune compromised patients, probiotic bacteria may also pose additional challenges, as weaker immune systems struggle to control bacterial growth.? To address the limitations of probiotics, alternative strategies have focused primarily on MDP analogs. These efforts include modifying the sugar moiety of MDP, altering the peptide side chains, or adding lipid tails. ?,?−? ? ? Compounds such as N-arylpyrazole and desmuramylpeptide agonists have demonstrated promising NOD2 activation profiles. ?−? ? However, they still remain structurally similar to MDP and have not been evaluated for their stability, oral bioavailability, or their safety in humans. Other molecules, such as mifamurtide, or muramyl tripeptide phosphatidyl ethanolamine, have been approved for testing and treatment in humans.? However, compounds such as mifamurtide only adapt existing PG fragments known to activate NOD2, and do not provide structurally diverse sources of activation. The longevity and stability of PG-based molecules in the body is also limited. Accordingly, there remains a critical need for structurally diverse NOD2 agonists that enable controlled NOD2 activation across a broad spectrum of clinical contexts.
We aimed to identify NOD2 agonists from a diverse library of FDA-approved small molecule drugs. These libraries represent an attractive entry point for drug discovery, as the compounds have already undergone extensive safety and toxicity assessments, thereby substantially lowering the time and cost required for clinical translation.? Additionally, they contain structurally diverse scaffolds that have demonstrated favorable pharmacokinetic properties, including oral bioavailability and metabolic stability – characteristics that are often lacking in natural product-derived agonists like MDP. To identify NOD2 agonists, we screened an FDA-approved drug library using NOD2-HEK-Blue NF-κB reporter cell lines. Among the full set of screened molecules, two structurally related hit compounds, each containing a benzimidazole moiety, were identified. Subsequent expansion of this chemical scaffold revealed multiple benzimidazoles capable of activating NOD2. Notably, one benzimidazole derivative not included in the original screen, nocodazole, exhibited nanomolar potency and elicited immune responses comparable to those induced by MDP.
Research Discussion
Identification of Benzimidazole NOD2 Agonists
First, we screened 1971 FDA approved drugs using the L1021 DrugDiscovery library (Apexbio Technology). The assay was initiated by incubating human NOD2-HEK-Blue NF-κB reporter cells (HEK-Blue NOD2 cells) with each compound at a concentration of 100 μM. This specific reporter cell line is engineered to have human NOD2 receptors and to express a secreted alkaline phosphatase (SEAP) downstream of NF-κB upon NOD2 activation.? The presence of SEAP in the cell culture medium dephosphorylates a small molecule dye, resulting in a color change that can be detected using UV–vis spectrophotometry (Figure S1). The gastrointestinal (GI) tract is a key site of NOD2 expression and activation, and also one where orally administered compounds can reach millimolar concentrations prior to systemic absorption and dilution. ?,? Accordingly, the primary screen was performed at elevated concentrations (100 μM) to mirror the high local compound levels anticipated in the GI tract following oral administration. ?−? ? ? Each plate included internal controls: MDP as a positive control and DMSO as a negative (vehicle) control. Fold-change values were calculated relative to these internal references.
The primary screen identified 90 candidate compounds that induced at least a 1.5-fold increase in activation relative to background (Figurec). This threshold was chosen to ensure that only compounds with activation levels clearly exceeding background were retained. To narrow down our potential hits, we analyzed the usage and administration methods of each candidate compound, removing those less suitable for the goal of NOD2 activation. We excluded compounds that are not typically administered orally, as they would not encounter NOD2 on the gut epithelium. Additionally, some of the compounds that displayed more than 1.5-fold activation had FDA approval as drugs of last resort with high toxicity profiles, therefore, they were eliminated from further evaluation. The remaining 76 compounds were rescreened in triplicate, and the most potent inducers of NOD2 were selected for further analysis. (Figured). Six of the compounds with the highest levels of NOD2 activation were identified and tested at a lower concentration: rigosertib, colchicine, alendronate sodium, azithromycin, mebendazole, and oxibendazole (Figurese and S2). The last round of testing demonstrated that alendronate sodium and azithromycin performed significantly worse than the other final hits. Based on our results and existing knowledge about these compounds, we excluded two from further evaluation. Rigosertib was excluded due to a lack of dose-dependent activity (Figure S3), and colchicine was excluded due to its high potential for toxicity, despite displaying dose dependence (Figure S4).? Instead, we selected mebendazole and oxibendazole for additional downstream evaluation (Figuref). These compounds displayed high levels of NOD2 activation in the reporter cell line and showed a strong dose dependence. Of note, both mebendazole and oxibendazole are antihelminth drugs and possess a relatively high safety profile (standard dosage in humans being 200 mg/day as a tablet through gut), allowing them to be administered in larger doses. ?,?
Expansion of Benzimidazole Class for NOD2 Activation
The two compounds with the highest potency and dose dependence, oxibendazole and mebendazole, share remarkable structural similarity. As such, we suspected that other compounds within the benzimidazole class might also be NOD2 agonists. A member of the benzimidazole family, GSK717, was previously developed by GSK as a NOD2 inhibitor. ?,? GSK717 has high selectivity toward NOD2 and is structurally dissimilar from oxibendazole or mebendazole outside of the core benzimidazole moeity. ?,? It was described to act as a competitive inhibitor, suggesting an ability to interact with the binding site of NOD2.? This interaction with the binding site supports the concept that benzimidazole containing molecules could potentially also act as agonists.
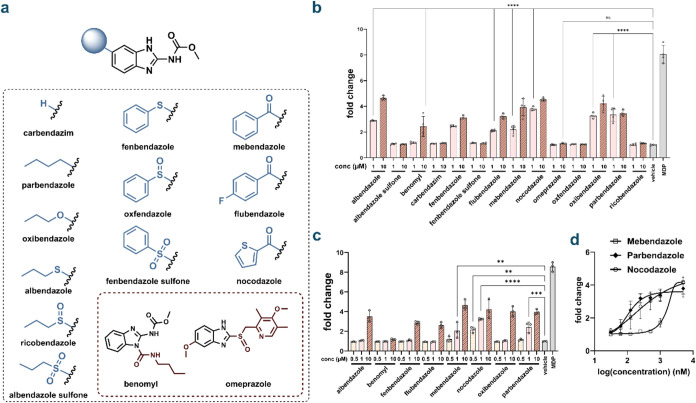
To further explore the benzimidazole pharmacophore, we tested 12 additional commercially available benzimidazole derivatives for their ability to activate NOD2 (Figurea). Of the compounds tested, approximately half exhibited NOD2-activating activity, with some inducing stronger activation than the original hits identified in the primary screen (Figureb). Many of these benzimidazoles are also antihelminth drugs, such as albendazole and fenbendazole. In the body, albendazole and fenbendazole are metabolized into their sulfoxide and sulfone forms (ricobendazole/albendazole sulfone and oxfendazole/fenbendazole sulfone respectively) which exert the anthelmintic effects. ?−? ? Interestingly, none of these metabolites display any NOD2 activity. Except for benomyl, all NOD2-activating benzimidazoles were active at both 1 and 10 μM, showing at least a 2-fold increase over background at the lower concentration. These results suggest that benzimidazoles may represent a new and general class of NOD2-activating compounds. Subsequent testing revealed that two of the 14 compounds, parbendazole and nocodazole, retained their NOD2 activation at submicromolar concentrations (Figurec). Further evaluation analysis showed that these two compounds activate NOD2 with EC_50_ values of 116.2 and 315.5 nM for parbendazole and nocodazole, respectively; notably, these new compounds both demonstrated a substantial improvement over one of the top hits from the primary screen (Figured). However, nocodazole and parbendazole cannot reach the maximum activating capability of MDP in HEK-Blue cells, reaching a plateau at a fold change of around 4 while MDP reaches a fold change of 6 over background (Figure S5). Although MDP induces higher NOD2 activity than both parbendazole and nocodazole, orally administered drugs become concentrated in the gut, occasionally reaching the millimolar range, and NOD2 receptors are present on the intestinal epithelium, allowing for a greater range of clinically relevant concentration for NOD2 agonists. ?−? ? ? The range in their biological activity as NOD2 activators indicates a specific pharmacophore that is responsible for the activation of NOD2 and suggests that these lead agents have potential for further development as NOD2-driven immunomodulatory agents.
(a) Chemical structures of a family of benzimidazole-based compounds that were selected for potential NOD2 activation based on the results from the primary screen. (b) NOD2 activation of benzimidazole compounds. HEK-Blue cells were treated with each compound at indicated concentrations for 16 h using a colorimetric assay (n = 3). MDP (1 μM) was used as a control. The analysis was performed by measuring the absorbance of each well at 655 nm. Fold change calculated by compound absorbance/average background. P values calculated by one-way ANOVA (ns = not significant, **** p < 0.0001). Bars mark the lowest concentration with statistically significant activation over background. (c) Dose dependence of NOD2 agonist benzimidazole compounds. HEK-Blue cells were treated with each compound at indicated concentrations for 16 h using a colorimetric assay (n = 3). MDP (1 μM) was used as a control. (d) EC50 curves of mebendazole, parbendazole, and nocodazole. HEK-Blue cells were treated with each compound at indicated concentrations for 16 h using a colorimetric assay (n = 3). The analysis was performed by measuring the absorbance of each well at 655 nm. Fold change calculated by compound absorbance/average background.
Benzimidazoles are also known to act on the colchicine binding site of tubulin.? However, they are typically designed to bind to tubulin in helminths, since they were synthesized to be antiparasitic agents. There is significant variation between tubulin of different organisms, but mebendazole has been shown to act on mammalian cells and induce apoptosis, especially in brain cancer cell lines. ?,? As such, the cytotoxicity of the most active benzimidazole compounds was tested using WTS. Benzimidazole compounds were added overnight to nonconfluent cells, so any effect on growth would also have been detected, as a lower cell count would also lead to a lower signal. No effect on cell viability or cell growth was observed, suggesting that tubulin remained unnaffected (Figure S6).
Confirmation of NOD2-Specific Activation by Benzimidazoles
To evaluate the specificity of benzimidazole-mediated NOD2 activation, we assessed cross-reactivity with NOD1 using HEK-Blue NOD1 reporter cell lines. NOD1 is another innate immune receptor that detects PG fragments? with its cognate ligand being γ-d-glutamyl-meso-diaminopimelic acid (iE-DAP).? Both NOD1 and NOD2 exhibit remarkable structural homology, sharing caspase-activating and recruitment domains (CARD), nucleotide-binding oligomerization domains (NOD), and leucine-rich repeat regions (LRR) (Figurea). ?,? This high degree of structural similarity makes NOD1 particularly susceptible to off-target effects from NOD2 agonists, rendering it an appropriate counter-screen for assessing compound selectivity. When evaluated in the NOD1 reporter system, benzimidazoles exhibited only background activation at both 10 and 1 μM (Figureb), demonstrating minimal cross-reactivity. The inability of benzimidazoles to activate NOD1, despite its structural similarity to NOD2, provides strong evidence for the specificity of these compounds as selective NOD2 agonists. Additionally, as both reporter cell lines utilized signal receptor activation through NF-kB, the lack of activation in HEK-Blue NOD1 cells indicates that the benzimidazoles are not interacting with other elements involved in NF-kB signaling, and activate the NOD2 receptor.
*(a) Protein domains of NOD1 and NOD2. (b) Activation of NOD1 by benzimidazoles. HEK-Blue NOD1 cells were treated with each compound at indicated concentrations for 16 h using a colorimetric assay (n = 4). P value determined by one-way ANOVA (ns = not significant, **** p < 0.0001). Only one statistical significance bar is shown for clarity. No significance was observed for any compound at either concentration. (c) Structure of GSK717. (d) Inhibition of parbendazole and nocodazole by GSK717 in reporter cells. HEK-Blue NOD2 cells were treated with each compound and GSK717 (10 μM) at indicated concentrations for 16 h using a colorimetric assay (n = 4). MDP (1 μM) was used as a control. P value determined by one-way ANOVA (ns = not significant, **** p < 0.0001). (e) Inhibition of cytokine release with GSK717. THP-1 cells were treated with each compound for 20 h and cytokine release was measured by ELISA from cell media. GSK717 was added at a concentration of 10 μM. P values determined by one-way ANOVA (ns = not significant,
- p < 0.05, **** p < 0.0001).*
To provide additional confirmation of NOD2-specific signaling, we employed the selective NOD2 inhibitor GSK717 to pharmacologically validate our findings. GSK717 functions as a competitive inhibitor of NOD2 and has been extensively characterized for its ability to inhibit NOD2 activation in both HEK-Blue NOD2 cells and THP-1 cells, exhibiting similar IC50 values across both systems (Figurec).? Co-treatment with GSK717 resulted in significant inhibition of NOD2 activation induced by the most potent benzimidazole compounds, nocodazole and parbendazole, in HEK-Blue NOD2 cells (Figured). While the degree of inhibition observed with nocodazole was somewhat reduced compared to MDP, the effect remained statistically significant. THP-1 cells, an immortalized human monocytic cell line, provide a physiologically relevant model for assessing NOD2 activation through quantification of cytokine release, particularly IL-8, which serves as a sensitive readout for both NOD2 and NOD1 signaling pathways.? The inhibitory profile observed in HEK-Blue-NOD2 cells was corroborated in THP-1 cells, where GSK717 significantly attenuated nocodazole-induced IL-6 release (Figuree). The consistent but incomplete inhibition of benzimidazole-mediated NOD2 activation across both cellular systems suggests that these compounds may engage the NOD2 binding site through a distinct molecular mechanism compared to MDP. Ligand docking of nocodazole with the crystal structure of NOD2? using ROSIE ?−? ? ? suggests that nocodazole does interact with the active site of NOD2 in the leucine rich region (LRR), but it may be positioned differently than the natural ligand MDP (Figure S7). Given the competitive nature of GSK717 inhibition, the observed differences in inhibitory efficacy likely reflect unique binding interactions between nocodazole and the NOD2 active site, which may not be fully antagonized by GSK717 under the experimental conditions employed.
Structural Activity Relationship Studies on Benzimidazoles
Structure–activity relationship (SAR) analysis revealed a critical role for carbonyl functionality in benzimidazole-mediated NOD2 activation. Some of the most potent benzimidazole NOD2 agonists, including mebendazole and nocodazole, share a common structural motif consisting of a carbonyl moiety positioned between two aromatic rings. This structural pattern extends to flubendazole, which also exhibits NOD2 agonistic activity (Figurea). In contrast, several benzimidazole analogs bearing sulfone substituents at analogous positions failed to induce NOD2 activation, suggesting that the specific electronic and steric properties of the carbonyl group are essential for biological activity (Figureb). To directly assess the functional importance of the carbonyl moiety, we synthesized reduced analogs of flubendazole, mebendazole, and nocodazole wherein the carbonyl groups were converted to their corresponding alcohols (Figurea). Evaluation of these reduced compounds in HEK-Blue NOD2 cells demonstrated a dramatic reduction in NOD2 activation, with activity decreased by at least 50% at the highest concentrations tested (Figureb–d). More strikingly, at lower concentrations (500 nM and 1 μM), the reduced benzimidazoles were completely inactive, displaying only background-level responses, whereas their parent carbonyl-containing compounds retained measurable NOD2 agonistic activity. These findings establish the carbonyl functionality as a critical pharmacophore element, demonstrating that its presence is essential for dual carbonyl benzimidazole agonists such as mebendazole and nocodazole to activate NOD2.
Structural alterations of benzimidazole agonists. (a) Chemical structures of reduced benzimidazole compounds and alterations to carbamate group. (b) Effect of reduced nocodazole on NOD2 activation. HEK-Blue NOD2 cells were treated with each compound at indicated concentrations 16 h using a colorimetric assay (n = 4). MDP (1 μM) was used as a control. P value calculated by one-way ANOVA (ns = not significant, * p < 0.05, *** p < 0.001, **** p < 0.0001). (c) Effect of reduced mebendazole on NOD2 activation. HEK-Blue NOD2 cells were treated with each compound at indicated concentrations for 16 h using a colorimetric assay (n = 4). MDP (1 μM) was used as a control. (d) Effect of reduced flubendazole on NOD2 activation. HEK-Blue NOD2 cells were treated with each compound at indicated concentrations for 16 h using a colorimetric assay (n = 4). MDP (1 μM) was used as a control. (e) Chemical structures of alterations to carbamate group on nocodazole and mebendazole. (f) Impact of carbamate group alteration on mebendazole induced NOD2 activation HEK-Blue NOD2 cells were treated with each compound at 10 μM for 16 h using a colorimetric assay (n = 4). MDP (1 μM) was used as a control. (g) Impact of carbamate group alteration on nocodazole induced NOD2 activation HEK-Blue NOD2 cells were treated with each compound at 10 mM for 16 h using a colorimetric assay (n = 4). MDP (1 μM) was used as a control.
All NOD2 active benzimidazole compounds identified feature a carbamate substituent attached to the benzimidazole core. The only two compounds evaluated that lack this functional group, benomyl and omeprazole, failed to consistently exhibit NOD2 activation (Figureb). The carbamate group is metabolically active and could be changed within cells to a different form. To systematically evaluate the functional contribution of the carbamate moiety, we synthesized structural analogs of mebendazole wherein the carbamate was replaced with either an amine or alkyne functionality (Figuree). The alkyne was selected for its potential as a reactive click handle, and to demonstrate that the molecule could be modified to contain additional functionalities that allow for future studies. The alkyne analog, which preserved the carbonyl component of the original carbamate, maintained comparable NOD2 activation to unmodified mebendazole (Figuref). In contrast, the amine analog exhibited dramatically reduced activity, with NOD2 activation diminished to near-background levels. Following these results, and alkyne analog of nocodazole was also synthesized. Nocodazole alkyne also retained similar NOD2 activity to its parent compound, nocodazole, displaying only a slight decrease in signal (Figureg). These SARs indicate that while the carbamate group is essential for optimal NOD2 activation by benzimidazoles, the functionality can tolerate modest structural modifications without complete loss of biological activity.
Benzimidazole Agonists Induce Cytokine Secretion from Immune
Cells
The therapeutic potential of NOD2 activation stems primarily from its downstream cytokine signaling cascades. Key inflammatory mediators associated with NOD2 signaling include IL-6 and IL-8, both pro-inflammatory cytokines regulated by NF-κB transcriptional activity. ?,?,? We employed THP-1 cells to assess the capacity of benzimidazole compounds to stimulate IL-8 and IL-6 release. THP-1 cells have been validated for quantifying NOD1 and NOD2 activation through measuring cytokine release.? Higher concentrations of MDP than expected were required induce significant IL-8 or IL-6 secretion from THP-1 cells, so the control concentration was increased from 1 μM to 50 μM. As such, lipopolysaccharide (LPS) served as an additional positive control, activating Toll-like receptors 2 and 4 (TLR2/TLR4) endogenously expressed in THP-1 cells.? Comparative analysis revealed that nocodazole induced more robust IL-8 secretion than parbendazole (Figurea). Interestingly, mebendazole also elicited stronger IL-8 release than parbendazole and maintained activity at lower concentrations more effectively than both nocodazole and parbendazole, despite showing weaker performance in HEK-Blue-NOD2 cells. In THP-1 cells, nocodazole exhibited particularly potent activity, with IL-8 levels frequently approaching the upper detection limit of the ELISA standard curve (Figurea). Nocodazole also induced IL-6 secretion, though to a lesser extent than IL-8 (Figureb). Reducing the nocodazole concentration to 1 μM led to a marked decrease in both IL-8 and IL-6 secretion, a reduction more pronounced than that observed in HEK-Blue-NOD2 cells (Figurea,b). Notably, nocodazole triggered higher cytokine secretion than MDP, even though MDP was used at a 5-fold higher concentration. These results contrast with those from the HEK-Blue-NOD2 reporter assay, in which MDP consistently outperformed the benzimidazole compounds at lower concentrations. When treated with GSK717, cytokine release by nocodazole is inhibited (Figurec), and the HEK-Blue NOD1 results (Figureb) When treated with GSK717, cytokine release by nocodazole is inhibited (Figurec), and the HEK-Blue NOD1 results demonstrate that benzimidazoles do not display off target affects with similar receptors or the downstream NF-kB signaling pathway. As such, the differing NOD2 activation profiles across concentrations and cell types suggest cell-type-specific responses to benzimidazoles and MDP, or differences in downstream biochemical processes.
(a) Release of IL-8. THP-1 cells were treated with each compound for 20 h and cytokine release was measured by ELISA from cell media (n = 4). MDP was added at 50 μM and LPS at 5 ng/mL. P value calculated by t test (ns = not significant, *** p < 0.001 **** p < 0.0001). (b) Release of IL-6. THP-1 cells were treated with each compound for 20 h and cytokine release was measured by ELISA from cell media (n = 4). P value calculated by one-way ANOVA ( p < 0.05). (c) Luminex assay results. Mouse BMDMs were treated with each compound overnight at 20 μM, with 10 ng/mL LPS when applicable. Cytokine levels were measured by Luminex.*
Beyond individual cytokine quantification, we sought to determine whether benzimidazole agonists generate the broader pro-inflammatory cytokine signature characteristic of MDP-induced NOD2 activation. Extensive profiling is critical, as the multifaceted therapeutic effects of NOD2 activation depend on the comprehensive immune response to multiple cytokines released rather than a single mediator. ?,?,?,? We therefore employed LUMINEX multiplex immunoassays to analyze supernatants from mouse bone marrow-derived macrophages (BMDMs) stimulated with benzimidazoles or MDP with LPS. The results demonstrate that MDP and benzimidazole agonists produce remarkably similar cytokine release patterns (Figurec). Both compound classes strongly induced IL-6 and IL-10 secretion, cytokines canonically associated with NOD2 signaling. ?,? Notably, nocodazole elicited more robust IL-1β release compared to MDP. Given that IL-1β represents another critical mediator of NOD2-dependent responses, this enhanced induction suggests potentially superior therapeutic efficacy. Collectively, these data indicate that nocodazole can engender the complex cytokine milieu associated with physiological NOD2 activation.
Conclusion
We have identified and characterized a novel class of FDA-approved benzimidazole compounds as potent NOD2 agonists. Among them, nocodazole, demonstrating nanomolar potency, emerged as the lead compound. Pharmacological validation confirmed that nocodazole activates NOD2-specific signaling and induces a cytokine release profile consistent with that triggered by the natural ligand MDP. Importantly, nocodazole features a scaffold structurally distinct from peptidoglycan-derived NOD2 agonists, offering the potential for improved pharmacological properties and reduced side effects. Given NOD2′s established roles in enhancing cancer immunotherapy and mitigating inflammatory bowel disease, these benzimidazole-based agonists represent promising therapeutic candidates. The discovery of nocodazole and related compounds also provides valuable tools for mechanistic studies of NOD2 biology and for the development of synthetic small-molecule immunomodulators.
Experimental Section
Cell Culture
HEK-Blue-NOD2 cells and HEK-Blue-NOD1 cells (InvivoGen, hkb-hnod2v2, and hkb-hnod1) were grown in DMEM (DMEM, Sigma-Aldrich, D6429) supplemented with 10% heat inactivated fetal bovine serum (FBS) (FBS, Thermo Scientific, A5670801) and 1% penicillin/streptomycin (P/S, Sigma-Aldrich, P4333). THP-1 (ATCC, TIB-202) cells were grown in RPMI-1640 media (RPMI-1640, Fisher Scientific, 11–875–119) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin. Bone marrow derived macrophages (BMDM) were obtained from the Bullock Lab. Cells were incubated at 37 °C with 5% CO_2_.
Screen of the L10121 Discovery Probe FDA Approved Library
When HEK-Blue cells reached 80–90% confluency, they were removed from their flasks with Trypsin/EDTA (ThermoFisher, 25200056) and resuspended in HEK-Blue detection media (HEK-Blue Detection, InvivoGen, hb-det2). Cells were then plated at 50,000 cells/well on a 96-well plate. Test compounds library plates were thawed 30 min beforehand, and DMSO solutions were pipetted directly into assay plates and then mixed with cells via pipetting. The same final volume of DMSO was used as a control. Cells were incubated at 37 °C and 5% CO_2_ for 16 h, and subjected to measure absorbance at 655 nm on a UV–vis plate reader. One μM MDP (N-Acetylmuramyl-L-alanyl-D-isoglutamine hydrate, Sigma-Aldrich, A9519) and 2 μL of DMSO were used as positive and negative controls, respectively.
HEK-Blue NOD2 Assay
When HEK-Blue cells reached 80–90% confluency, they were removed from their flasks with Trypsin/EDTA and resuspended in HEK-Blue detection media. Benzimidazole compounds (see materials) were diluted sequentially from DMSO stocks into acetonitrile and then into HEK-Blue Detection media to obtain desired final concentration. The same final volume of acetonitrile was added to cells as the “vehicle” control. were then plated at 50,000 cells/well on a 96-well plate and treated with compounds for 16 h, incubating at 37 °C and 5% CO_2_. Data was acquired at 655 nm on a UV–vis plate reader.
ELISA
When THP-1 cells reached a confluence in the flask approaching 8 × 10^6^ cells/mL, they were spun down at 1000g for 5 min, resuspended in fresh growth media, and plated at 500,000 cells/well on a 24-well plate. Benzimidazole compounds were diluted sequentially from DMSO stocks into acetonitrile and then into RPMI media. Cells were incubated with compounds for 20–24 h. The cell media was extracted and used for IL-8 and IL-6 ELISAs (ELISA-MAX Deluxe Set Human IL-8/IL-6, BioLegend, 431504/430504). The ELISA was conducted following the procedure laid out in the product information for the BioLegend Deluxe ELISA kit.
BMDM Isolation
Mouse BMDMs were derived from BM isolated from the femurs and tibias of 8- to 10-week-old C57BL/6J mice and cultured in RFHP10 media (RPMI 1640 supplemented with 10% heat-inactivated FBS, 10 mM N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic acid, and 1% penicillin–streptomycin–l-glutamine) supplemented with 20 ng/mL of M-CSF for 10 days. Media was replaced with fresh RFHP10 supplemented with 20 ng/mL of M-CSF on day 5 and BMDMs were used from day 6–10. The animal study protocol was approved by the Animal Care and Use Committee of the University of Virginia (protocol code 4270).
LUMINEX
BMDMs were removed from their growth plates with Trypsin/EDTA and resuspended in RFHP10 supplemented with M-CSF. Compounds were added at a concentration of 20 μM. Benzimidazole compounds were diluted sequentially from DMSO stocks into acetonitrile and then into DMEM media. A fresh stock of LPS from E. coli was added at 10 ng/mL in relevant conditions. BMDMs incubated at 37 °C and 5% CO_2_ overnight with the compounds and controls. Media was removed and frozen at −80 °C for storage. The media was thawed and sent off for LUMINEX at the UVA flow core.
Inhibition by GSK717
HEK-Blue NOD2 cells were grown in appropriate media (listed above). The cells were grown to approximately 80% confluency, removed from their flasks with Trypsin/EDTA and resuspended in HEK-Blue detection media. Benzimidazole compounds were diluted sequentially from DMSO stocks into acetonitrile and then into HEK-Blue Detection media. GSK717(MedChem Express, HY136555) was diluted from DMSO stock into ethanol and then added to the plate directly at the same time as compounds and positive controls. Cells were then plated at 50,000 cells/well on a 96-well plate and treated with compounds for 16 h, incubating at 37 °C and 5% CO_2_. Results were read at 655 nm on a UV–vis plate reader.
Statistical Analysis
Unless otherwise specified, statistical analysis was conducted using GraphPad Prism 9.5. Experiments were conducted in triplicate and significant experiments were repeated at least twice. One-way ANOVA was used to calculate statistical significance. Error bars represent standard deviation.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al Nabhani Z.Dietrich G.Hugot J.-P.Barreau F.Nod 2: The Intestinal Gate Keeper P Lo S Pathog.2017133 e 100617710.1371/journal.ppat.100617728253332 PMC 5333895 · doi ↗ · pubmed ↗
- 2Irazoki O.Hernandez S. B.Cava F.Peptidoglycan Muropeptides: Release, Perception, and Functions as Signaling Molecules Front. Microbiol.20191050010.3389/fmicb.2019.0050030984120 PMC 6448482 · doi ↗ · pubmed ↗
- 3Wolf A. J.Underhill D. M.Peptidoglycan Recognition by the Innate Immune System Nat. Rev. Immunol.201818424325410.1038/nri.2017.13629292393 · doi ↗ · pubmed ↗
- 4Liu Z.Zhang Y.Jin T.Yi C.Ocansey D. K. W.Mao F.The Role of NOD 2 in Intestinal Immune Response and Microbiota Modulation: A Therapeutic Target in Inflammatory Bowel Disease Int. Immunopharmacol.202211310946610.1016/j.intimp.2022.10946636435061 · doi ↗ · pubmed ↗
- 5Trindade B. C.Chen G. Y.NOD 1 and NOD 2 in Inflammatory and Infectious Diseases Immunol. Rev.2020297113916110.1111/imr.1290232677123 PMC 8928416 · doi ↗ · pubmed ↗
- 6Medzhitov R.Toll-like Receptors and Innate Immunity Nat. Rev. Immunol.20011213514510.1038/3510052911905821 · doi ↗ · pubmed ↗
- 7Oliveira-Nascimento L.Massari P.Wetzler L.The Role of TLR 2 in Infection and Immunity Front. Immunol.201237910.3389/fimmu.2012.0007922566960 PMC 3342043 · doi ↗ · pubmed ↗
- 8Mc Inturff J. E.Modlin R. L.Kim J.The Role of Toll-like Receptors in the Pathogenesis and Treatment of Dermatological Disease J. Invest. Dermatol.200512511810.1111/j.0022-202X.2004.23459.x 15982296 · doi ↗ · pubmed ↗