Inflammatory Factors and Chronic Rhinosinusitis: An Umbrella Review
Stergios T Lialiaris, Konstantinos Chaidas, George Fyrmpas, Theodora Eleftheria Deftereou, Dimitra Spyroulia, Emmanuel P Prokopakis, Michael Katotomichelakis

TL;DR
This umbrella review explores how different inflammatory pathways contribute to chronic rhinosinusitis and highlights the need for better classification and treatment strategies.
Contribution
The study provides a comprehensive synthesis of inflammatory pathways in chronic rhinosinusitis, emphasizing distinct immune patterns and their clinical implications.
Findings
Type 1 inflammation is associated with CRSsNP and involves interferon-gamma and IL-12 signaling.
Type 2 inflammation is linked to CRSwNP and involves Th2 cytokines and epithelial-derived mediators.
Type 3 inflammation involves IL-17 and IL-22 and is relevant to fungal and bacterial rhinosinusitis.
Abstract
Chronic rhinosinusitis (CRS) is a common inflammatory disorder of the nasal mucosa and paranasal sinuses characterized by persistent sinonasal symptoms and objective endoscopic and imaging evidence of the disease. CRS is broadly described as a disease with two different phenotypes: without (CRSsNP) or with nasal polyps (CRSwNP), but substantial heterogeneity in clinical presentation and underlying mechanisms complicates classification and treatment selection. In accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines, we conducted an umbrella review supported by a systematic literature search in Scopus and PubMed, screening publications from May 1968 to August 2025 and ultimately including 64 studies, with findings synthesized qualitatively. The evidence supported the concept that the inflammatory pathways in CRS reflect distinct, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
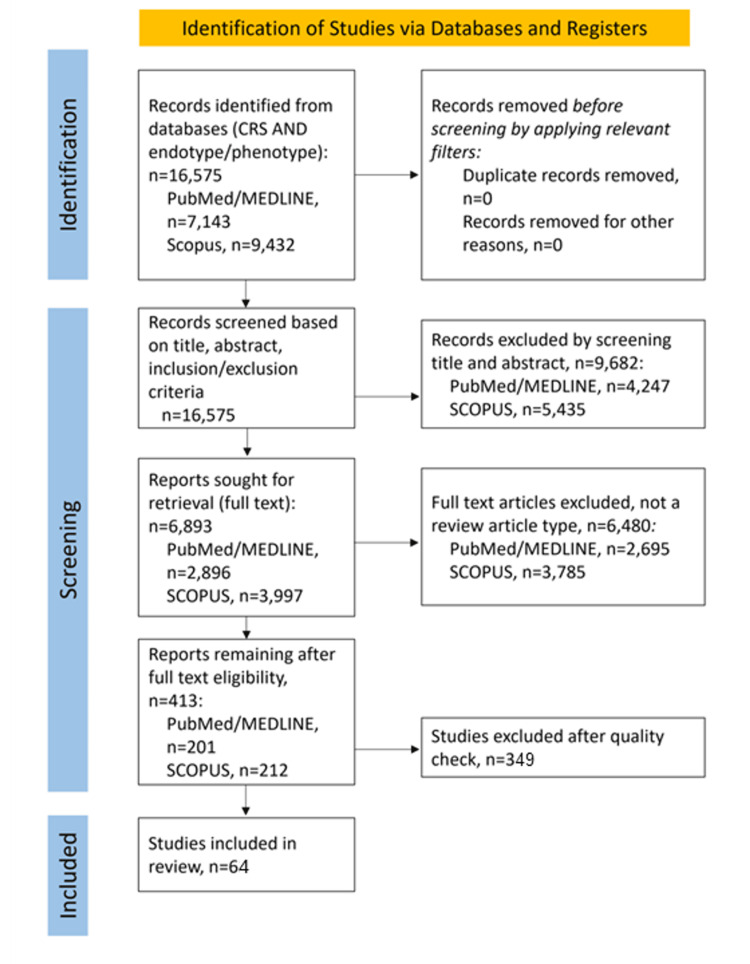 Figure 1
Figure 1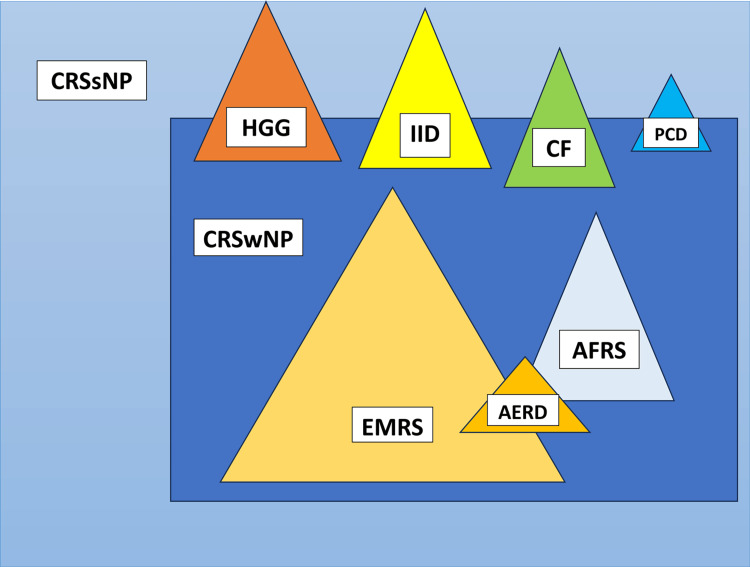 Figure 2
Figure 2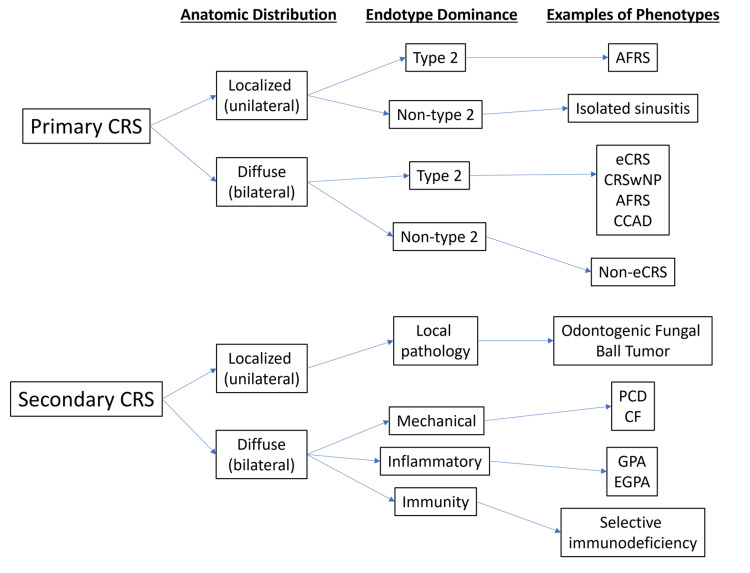 Figure 3
Figure 3| Mediator | Family members | Year of discovery/first reports | Major source cells | Receptor(s) | Primary target cells/tissues | Key actions/functions | CRS inflammatory pattern | Year linked to CRS/CRS therapy | References |
| IL-1 | IL-1α, IL-1β, IL-1Ra, IL-36α, IL-36β, IL-36γ, IL-36Ra, IL-18, IL-33, IL-37, IL-38 | 1943-48 / 1979 | Monocytes, macrophages, B cells | IL1R1, IL1R2 | Natural killer (NK) cells, Th cells, B cells, endothelial cells, other cells | Activation; stimulation; maturation; proliferation; inflammation | All types | 2016 | [ |
| IL-2 | IL-4, IL-7, IL-9, IL-15, IL-21 | 1943-48 / 1979 | T helper 1 (Th1) cells | IL2RA, IL2RB, IL2RG | NK cells, T cells, B cells, macrophages | T-cell stimulation; treatment of cancer; transplantation; HIV | Type 1 and 3 inflammation | 2012 | [ |
| IL-4 | - | 1982 / 1986 | T helper 2 (Th2) cells, mast cells, macrophages, CD4+ T cells | IL4Rα | T cells, B cells, endothelial cells | Differentiation; proliferation; role in allergy | Type 2 inflammation | 2016 | [ |
| IL-5 | IL-5, IL-5Rα | 1986 | Th2 cells, mast cells, eosinophils | IL5Rα, IL5Rβ | B cells, eosinophils | Proliferation; differentiation | Type 2 inflammation | 2006 | [ |
| IL-6 | - | 1986 | B cells, Th2 cells, endothelial cells, macrophages | IL6Rα, IL6Rβ | T cells, B cells, hematopoietic cells | Differentiation; antibody secretion; inflammation | Type 1 and type 3 inflammation | 2016 | [ |
| IL-8 | - | 1990 | Macrophages, lymphocytes, epithelial cells, endothelial cells | IL8Rα, IL8Rβ | Neutrophils, basophils, lymphocytes | Neutrophil chemotaxis | Type 1 inflammation | 2012 | [ |
| IL-9 | - | 1980s / 1990 | Th2 cells, CD4+ T cells | IL9R | T cells, B cells | Potentiation; stimulation | Type 2 inflammation | [ | |
| IL-10 | - | 1995 | B cells, Th2 cells, CD8+ T cells, mast cells, macrophages | IL10Rα, IL10Rβ | B cells, Th1/Th2 cells, mast cells, macrophages | Activation; stimulation; inhibition of IFN and TNF production | Type 1 inflammation | 2000 | [ |
| IL-12 | - | 1998 | B cells, T cells, macrophages | IL12Rβ1, IL12Rβ2 | NK cells, T cells | Differentiation; stimulation of IL-2, IFN-γ, TNF-α, and IL-10 | Type 1 inflammation | 2007 | [ |
| IL-13 | - | 1993 | NK cells, Th2 cells, mast cells | IL13R | B cells, Th2 cells, macrophages | Stimulation; differentiation; inhibition of cytokine production (e.g., IL-1, IL-6, IL-8, IL-10, IL-12) | Type 2 inflammation | 2016 | [ |
| IL-17 | - | 2002 | T helper 17 (Th17) cells | IL17Rα, IL17Rβ | Endothelial cells, epithelial cells | Angiogenesis; inflammation | Type 3 inflammation | 2001 | [ |
| IL-22 | - | 2000 | Th17 cells | IL22R | - | Production; activation | Type 3 inflammation | 2016 | [ |
| IL-25 | - | 2001 | Mast cells, eosinophils, macrophages, epithelial cells, T cells | LY6Ε | - | Induces IL-4, IL-5, and IL-13 | Type 2 and 3 inflammation | 2010 | [ |
| IL-33 | - | 2010 | Multiple cell types (e.g., macrophages, endothelial cells, epithelial cells) | - | - | Induces T-helper cells to produce type 2 cytokines | Type 2 inflammation | 2014 | [ |
| TNFα | - | 1968 | Macrophages | TNFR1, TNFR2 | Immune system | Adipokine: promotes insulin resistance; cytokine: mediates cell signaling | Type 1 and non–type 2 inflammation | 2016 | [ |
| TGFβ | - | 1997 | All white blood cell lineages, including macrophages | TGF-β1 to 3 | - | Regulates key cellular activities | Type 1, non–type 2, and type 3 inflammation | 2016 | [ |
| IFN-γ | - | 1965 | Adaptive immune cells | IFNGR1, IFNGR2 | NK cells, CD4+ Th1 cells, CD8+ cytotoxic T cells | Type II interferon; regulates target-cell immune responses and shapes signaling within the immune system | Type 1, 2, and 3 inflammation | 2016 | [ |
| ECP | - | 1989 | Eosinophil primary matrix | - | - | Member of the ribonuclease A superfamily; neurotoxic, helminthotoxic, and ribonucleolytic activities | Type 1, 2, and 3 inflammation | 1997 | [ |
| MPO | - | 1983 | Neutrophils | - | - | Catalyzes the conversion of hydrogen peroxide and chloride ions into hypochlorous acid | Type 1 and non–type 2 inflammation | 2005 | [ |
| IgE | - | 1966 | Plasma cells | FcεRI (type I Fcε receptor), FcεRII (type II Fcε receptor) | - | Antibody in mammals | Type 3 inflammation | 2005 | [ |
| Study | Study Design | Risk of Bias | Score | Tool | Main Domains Assessed | Overall Risk |
| Fokkens et al., 2020 (EPOS 2020) [ | Clinical practice guideline | Assessed | AGREE II (domain-based) | AGREE II | Scope & purpose; Stakeholder involvement; Rigor; Clarity; Applicability; Editorial independence | Low |
| Payne et al., 2025 [ | Clinical practice guideline | Assessed | AGREE II (domain-based) | AGREE II | Scope; Rigor; Clarity; Applicability; Editorial independence | Low–Moderate |
| Mattos et al., 2019 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Van Crombruggen et al., 2011 [ | Narrative review | Assessed | SANRA 10/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Tomassen et al., 2016 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Meltzer et al., 2004 [ | Consensus review | Assessed | SANRA 11/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Cho et al., 2020 [ | Narrative review | Assessed | SANRA 10/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Avdeeva & Fokkens, 2018 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Papacharalampous et al., 2024 [ | Systematic review | Assessed | AMSTAR-2: Moderate | AMSTAR-2 | Protocol; Study selection; Risk of bias; Synthesis | Moderate |
| Staudacher et al., 2020 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Ahern & Cervin, 2019 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Wang et al., 2016 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Schleimer, 2017 [ | Narrative review | Assessed | SANRA 11/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Cao et al., 2019 [ | Narrative review | Assessed | SANRA 10/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Bachert et al., 1997 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Gevaert et al., 2006 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Hirano et al., 1986 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Seyfizadeh et al., 2015 [ | Narrative review | Assessed | SANRA 8/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Aggarwal & Gurney, 2002 [ | Narrative review | Assessed | SANRA 8/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Tesmer et al., 2008 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Massagué, 2012 [ | Narrative review | Assessed | SANRA 11/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Klebanoff, 2005 [ | Narrative review | Assessed | SANRA 10/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Takhar et al., 2005 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Akdis et al., 2013 [ | Consensus review | Assessed | SANRA 11/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Hirschberg et al., 2016 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Bachert et al., 2001 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Van Bruaene et al., 2008 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Gratz et al., 2013 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Van Bruaene et al., 2009 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Al-Alawi et al., 2014 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Licona-Limon et al., 2013 [ | Narrative review | Assessed | SANRA 10/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Low |
| Shaw et al., 2013 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Kato et al., 2007 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Hamilos et al., 1995 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Zhang et al., 2018 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Seshadri et al., 2015 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Cho & Kim, 2018 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Ni Mhurchu et al., 2017 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Montone et al., 2012 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Turner et al., 2013 [ | Systematic review | Assessed | AMSTAR-2: Moderate | AMSTAR-2 | Search strategy; Selection; Outcome synthesis | Moderate |
| Steinke & Borish, 2016 [ | Narrative review | Assessed | SANRA 9/12 | SANRA | Justification; Literature search; Referencing; Scientific reasoning | Moderate |
| Aeumjaturapat et al., 2003 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Dhong et al., 2000 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Szczeklik et al., 2000 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
| Rajan et al., 2015 [ | Meta-analysis | Assessed | AMSTAR-2: Moderate | AMSTAR-2 | Study selection; Bias assessment; Statistical synthesis | Moderate |
| Farrell, 2008 [ | Observational study | Assessed | NOS 7–8/9 | Newcastle–Ottawa Scale | Selection; Comparability; Outcome | Low |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSinusitis and nasal conditions · Allergic Rhinitis and Sensitization · Asthma and respiratory diseases
Introduction and background
Chronic rhinosinusitis (CRS) is a serious health problem with a prevalence ranging from 5% to 12% [1]. CRS is described as inflammation of the nasal mucosa and/or the paranasal sinuses and the patients often complain about two or more of the following symptoms: nasal blockage, nasal obstruction, nasal congestion, or nasal discharge (anterior/posterior nasal drip); facial pain or pressure; and reduced sense of smell [1]. Rhinosinusitis can also lead to intracranial and ocular complications [2]. These symptoms should correlate with endoscopic signs and computed tomography (CT) findings [1]. To qualify as chronic, symptoms should persist for at least 12 weeks. Environmental and genetic factors, as well as factors associated with modern lifestyles (e.g., travel, environmental pollution, and diet), contribute to CRS and may help explain differences across populations [3].
CRS occurs either without (CRSsNP) or with nasal polyps (CRSwNP). It is heterogeneous in its clinical presentation and molecular pathophysiology, with variability in both phenotypes (observable clinical features) and endotypes (underlying biologic mechanisms). However, the specific mechanisms remain incompletely understood, which limits accurate classification [4,5].
Despite these differences, treatment options for CRS are typically focused on corticosteroids or sinonasal surgery. In 2019, dupilumab, a monoclonal antibody, was approved in the United States of America (USA) as a biologic therapy for CRSwNP. Dupilumab blocks interleukin IL-4 and/or IL-13 signaling by targeting the IL-4 receptor alpha chain. Other monoclonal antibodies, including those targeting the IL-5 receptor (benralizumab), immunoglobulin E (IgE; omalizumab), or IL-5 (mepolizumab), have been evaluated in clinical trials for polypoid CRS [6-8]. These newer therapies may enable more individualized treatment by matching therapy to specific CRS subtypes; however, available options remain limited, and further categorization is needed. Therefore, the objective of this umbrella review was to synthesize evidence on key inflammatory mediators and immune endotypes associated with CRS phenotypes (CRSsNP and CRSwNP) and to summarize how these pathways inform emerging endotype-driven management.
Review
We conducted an umbrella review supported by a systematic literature search in two electronic databases (Scopus and PubMed) in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [9]. Given the heterogeneity of evidence in this field, we synthesized findings qualitatively across multiple evidence types rather than performing a quantitative meta-analysis. We applied strict selection criteria, including English language, clear study protocols, and well-documented results. We searched the databases using the following keywords: “Chronic Rhinosinusitis,” “endotype,” “review,” and “phenotype.” We used keywords individually and in combination (e.g., “Chronic Rhinosinusitis” AND “endotype”) (Figure 1).
PRISMA flow diagram showing the selection of articlesPRISMA: Preferred Reporting Items for Systematic Reviews and Meta-Analyses.
We initially screened the published literature from May 1968 to August 2025 that included an English-language abstract. We excluded non-human studies and studies published in languages other than English or without an English abstract. We included studies involving only adult populations. Based on these criteria, we ultimately included 64 studies/texts/works in this review.
Results
To date, no widely accepted criteria guide the optimal therapeutic approach for CRSsNP or CRSwNP, and for many years, experts largely accepted the clinicopathological heterogeneity of CRS. Since 2004 [6], however, a group of rhinosinusitis experts has proposed that CRS can be divided into three main types: CRSwNP, CRSsNP, and allergic fungal rhinosinusitis (AFRS). Figure 2 shows the possible phenotypes of CRS, with CRSsNP comprising approximately 80% and CRSwNP approximately 20%; within CRSwNP, AFRS, eosinophilic mucin rhinosinusitis, and aspirin-exacerbated respiratory disease (AERD) exist as subsets [7-11].
Various CRS phenotypesCRS, Chronic Rhinosinusitis; CRSsNP, Chronic Rhinosinusitis without nasal polyps; CRSwNP, Chronic Rhinosinusitis with nasal polyps; IID, Innate Immune defects; HGG, Hypogammaglobulinemia; CF, Cystic fibrosis; PCD, Primary ciliary dyskinesia; EMRS, Eosinophilic mucin rhinosinusitis; AFRS, Allergic fungal sinusitis; AERD, Aspirin-exacerbated respiratory disease.Image credit: Created by Lialiaris S using Microsoft Word (Microsoft Corp., Redmond, WA, USA).
Figure 2 also depicts the subsets of CRS associated with innate immune defects, hypogammaglobulinemia, cystic fibrosis (CF), and primary ciliary dyskinesia, which overlap both CRSwNP and CRSsNP with controversial rates. Other rare phenotypes are not listed in the diagram [7-11]. Nevertheless, CRS symptoms can vary substantially across patients.
Factors causing inflammation in CRS
To understand CRS in depth, the key pathophysiological mechanisms underlying its clinical manifestations must be clarified. Current concepts propose that inflammation of the nasal mucosa may be driven, in part, by epithelial barrier disruption and immune dysregulation. To fully explain the diversity of CRS, investigators must evaluate multiple inflammatory response patterns, and additional patterns may be identified in the future. Available evidence suggests that CRS inflammatory pathways are dominated by T helper cells (Th1, Th2, Th17, Th22), and regulatory T cells (Treg) (Figure 3) [1,12].
Classification of primary and secondary CRSAFRS, allergic fungal rhinosinusitis; CCAD, central compartment allergic disease; CRS, chronic rhinosinusitis; CRSwNP, chronic rhinosinusitis with nasal polyps; eCRS, eosinophilic CRS; OMC, ostiomeatal complex; PCD, primary ciliary diskinesia; CF, cystic fibrosis; GPA, granulomatosis with polyangiitis (Wegner’s disease); EGPA, eosinophilic granulomatosis with polyangiitis (Churg-Strauss disease) [1].Image credit: Created by Lialiaris S using Microsoft Powerpoint (Microsoft Corp., Redmond, WA, USA).
Investigators commonly describe three immune-inflammatory patterns that differ in the sequence of the inflammatory mediators, the immune cell populations involved, and the associated physiologic functions. Type 1 inflammation (often associated with CRSsNP) is associated with the presence of interferon-gamma (IFN-γ) and IL-12 produced by Th1 cells [13]. Type 2 inflammation (associated with CRS, particularly CRSwNP) is defined by producing IL-4, IL-5, and IL-13 by Th2 cells. Type 3 inflammation (also referred to as type 17 by some authors) is characterized by increased IL-17 and IL-22 produced by Th17 and Th22 cells, respectively [14-16]. In addition, Treg cell maturation promotes production of transforming growth factor beta (TGF-β) [17]. Table 1 describes the cytokines and other factors involved in CRS [1,5,18-40].
Type 1 inflammation
The literature often describes type 1 inflammation as non-type 2 inflammation, and CRS most commonly presents without nasal polyps. This pattern also shows features of mixed type 1 and type 3 inflammation with prominent neutrophil infiltration. Pathogen invasion of the nasal epithelium triggers epithelial release of IL-6, IL-8, tumor necrosis factor alpha, and other chemokines. Pattern-associated molecular pattern/Toll-like receptor immune responses appear to stimulate production of IFN-γ and IL-8 [40,41]. These responses promote IL-8 release and recruit immune cells (e.g., neutrophils) to the paranasal sinuses, thereby shaping downstream immune responses. Neutrophils subsequently release mediators including IL-1, IL-6, IL-8, and myeloperoxidase, an enzyme derived from neutrophil granules [42]. In response to pathogens, epithelial IFN-γ promotes differentiation of CD4+ T cells into Th1 cells [40,41]. Epithelial IL-6 promotes differentiation of CD4+ T cells into Th17 and Th22 cells; Th17 cells secrete IL-17 and IL-22, whereas Th22 cells secrete IL-22 [16].
Studies have reported upregulation of Treg cells in patients with CRSsNP compared with normal individuals, whereas patients with CRSwNP show downregulation [43]. In immune regulation, Treg cells downregulate Th1 and Th2 activity and influence IL-10 production [44]. Evidence indicates that TGF, a cytokine family member that may contribute to tissue growth in CRSsNP, is produced by Treg cells [44]. In addition, TGF-β plays a key role in fibroblast growth. Conversely, increased extracellular matrix synthesis, which contributes to upper airway epithelial remodeling, may increase symptom burden in CRSsNP [45]. TGF-β also participates in the differentiation of CD4+ T cells toward Th17 cells and in the maturation of Treg cells [43,45]. The precise role of Treg cells in CRS, including their capacity to reduce inflammation via IL-10 production and their potential contribution to airway remodeling and fibrosis, remain under investigation.
Type 2 inflammation
Type 2 inflammation in CRS has been well described. Nasal epithelial cells can be stimulated to secrete thymic stromal lymphopoietin (TSLP) [46,47], IL-25, and IL-33, which promote production of IL-4, IL-5, and IL-13 from epithelial cells and mucosal mast cells [48,49]. In addition, TSLP and IL-13 induce type 2 cytokine production by innate lymphoid cells [50]. Evidence also suggests that TSLP stimulates myeloid dendritic cells (mDCs) [49,51]. Once activated, mDCs present antigen and provide costimulatory signals that promote CD4+ T-cell differentiation toward Th2 cells. Th2 cells produce type 2 cytokines, which are central to type 2 inflammation. In an IL-4-rich environment that promotes Th2 expansion (Table 1), IL-5 supports eosinophil infiltration, which can induce eosinophilic extracellular traps, release of toxic proteins, and inflammation [52]. IL-4 and IL-13 upregulate mucin 5AC and mucin 5B gene activity, which encodes mucins and increases pendrin production [53]. Pendrin, an epithelial anion transporter, increases mucus production [54] and may enhance type 3 (or type 17) inflammation, with increased IL-17 and IL-22 levels [14,15] and by causing hypoxia [55].
Type 3 inflammation: fungal rhinosinusitis
Fungal diseases of the nose and paranasal sinuses are collectively termed fungal rhinosinusitis (FR). FR is broadly classified as invasive or noninvasive. Invasive FR involves fungal hyphae infiltrating the mucosa of the nose and paranasal sinuses [56]. Acute, chronic, and granulomatous invasive FR have been described [57]. Immunocompromised patients, including those with uncontrolled diabetes, are prone to invasive FR. Poor survival has been reported in patients of advanced age, those with intracranial extension, and those who do not undergo surgery as part of the treatment protocol [58].
AFRS is characterized by Th2 inflammation and fungal colonization of the nasal mucosa. Dematiaceous fungi and Aspergillus species account for most cases [57,59]. In AFRS, allergic inflammation occurs without fungal tissue invasion. AFRS may be unilateral or bilateral and can produce “allergic mucus” or “eosinophilic mucus,” which are thick inflammatory secretions characterized by tenacious mucus containing clusters of eosinophils and free eosinophilic cocci [60]. Another noninvasive form is the fungal ball, which typically occurs in middle-aged or older patients and presents as unilateral sinusitis, most often involving the maxillary sinuses, with microcalcification patterns on CT [61]. Fungal ball disease is often asymptomatic and may be detected incidentally on CT or magnetic resonance imaging (MRI).
Aspirin-exacerbated respiratory disease (AERD) and cystic fibrosis (CF)
AERD is a CRS syndrome characterized by eosinophilic nasal polyps, asthma, and respiratory reactions induced by aspirin and other nonsteroidal anti-inflammatory drugs (NSAIDs) that inhibit cyclooxygenase-1, with onset typically in adulthood [62]. Its prevalence is approximately 30% in patients with asthma and CRSwNP [63]. Respiratory reactions to aspirin and other NSAIDs are common in patients with CF, and mutations in the CF transmembrane conductance regulator gene are also reported in non-CF patients with CRS [64]. Across all ages, patients with CF have an increased prevalence of CRS. Management of CRS in CF is challenging due to increased mucus viscosity, chronic sinus infections, bacterial colonization and biofilm formation, and the presence of distinct sinus pathogens.
Discussion
CRS is a major focus in otorhinolaryngology, and research in this field continues to expand. The number of patients affected by CRS appears to be increasing, and investigators continue to evaluate mechanisms and therapies to improve outcomes [4,5]. The defining feature of CRS is persistent inflammation of the nasal mucosa, which may occur with or without nasal polyps. CRS is heterogeneous in both clinical presentation and underlying pathophysiology, including variability in phenotypes and genotypes. However, pathogenic mechanisms remain incompletely understood, limiting precise classification [4,5].
Our review of the international literature suggests that Th1 responses are characterized by IFN-γ and IL-12 expression, whereas Th2 responses are associated with elevated IL-4, IL-5, and IL-13 levels. More recently, type 3 (or type 17) inflammation has been described and is associated with increased IL-17 and IL-22 production [14,15]. Th1 responses primarily protect against intracellular microbes, including viruses. In contrast, type 2 inflammation contributes to immune responses against parasitic infections and is also linked to allergic rhinosinusitis. Type 3 inflammation is thought to support host defense against bacteria and fungi [14,15,19-23].
Geographic variation, age, asthma, fungi, and other infectious agents or medications may influence inflammatory pathways in CRS. Accordingly, CRS inflammation can show mixed type 1, type 2, and type 3 profiles across classic phenotypes with or without nasal polyps [1-5]. Endotype-based evaluation may help distinguish biologically distinct forms of CRS and support the selection of more individualized treatment strategies. Defining each patient’s inflammatory pattern may facilitate targeted therapy, and several biologic therapies are already available for selected patients with CRS. Continued research is needed to refine CRS phenotypes and endotypes and to advance individualized treatment approaches.
Limitations
This umbrella review has several limitations. First, the search strategy was limited to two electronic databases (Scopus and PubMed) and a predefined set of keywords, potentially missing relevant studies indexed in other databases or described using different terminology. Second, we restricted inclusion to studies published in English or with an English abstract and to adult populations, which may limit generalizability and introduce language- and selection-related bias. Third, the review spans literature published between May 1968 and August 2025. During this period, definitions, diagnostic criteria, endoscopic scoring systems, imaging practices, and molecular characterization methods for CRS have evolved substantially, thereby increasing heterogeneity and reducing comparability across included studies. Fourth, the included body of evidence likely reflects variability in how phenotypes and endotypes were defined and measured, including differences in tissue sampling sites, laboratory platforms, cytokine panels, and thresholds used to classify inflammatory patterns, which may limit synthesis into a unified framework. Given these limitations, we added a risk-of-bias table (Table 2) to provide a structured appraisal of study quality; however, we did not quantitatively pool findings across studies.
Finally, publication bias and selective reporting cannot be excluded, particularly for emerging biologic targets or therapies.
Conclusions
Multiple studies have examined CRS phenotypes and endotypes. Further progress is expected as technological advances improve the identification of CRS subtypes. In parallel, ongoing investigation of biologic factors, including thymic stromal lymphopoietin, IL-33, and IL-25, may further distinguish known CRS endotypes and help identify patients who do or do not respond to specific agents. Personalized management of upper airway disease, including CRSwNP, should be a priority in the coming decade, particularly for patients with severe CRS. Biologic therapies may offer additional treatment options once patients are categorized by phenotype and endotype. However, broader implementation requires improved understanding of which monoclonal antibody is most appropriate for each CRS endotype and phenotype. Earlier identification of these subgroups at disease onset may support earlier treatment, reduce the need for repeated surgical procedures in subsequent years, and potentially lower the risk of progression to lower airway disease.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1European position paper on rhinosinusitis and nasal polyps 2020 Rhinology Fokkens WJ Lund VJ Hopkins C 146458202010.4193/Rhin 20.60032077450 · doi ↗ · pubmed ↗
- 2Clinical practice guideline: adult sinusitis update Otolaryngol Head Neck Surg Payne SC Mc Kenna M Buckley J 0173202510.1002/ohn.134440742114 · doi ↗ · pubmed ↗
- 3Symptom importance, patient expectations, and satisfaction in chronic rhinosinusitis Int Forum Allergy Rhinol Mattos JL Rudmik L Schlosser RJ Smith TL Mace JC Alt J Soler ZM 593600920193074810110.1002/alr.22309 PMC 6555643 · doi ↗ · pubmed ↗
- 4Pathogenesis of chronic rhinosinusitis: inflammation J Allergy Clin Immunol Van Crombruggen K Zhang N Gevaert P Tomassen P Bachert C 72873212820112186807610.1016/j.jaci.2011.07.049 · doi ↗ · pubmed ↗
- 5Inflammatory endotypes of chronic rhinosinusitis based on cluster analysis of biomarkers J Allergy Clin Immunol Tomassen P Vandeplas G Van Zele T 1449145613720162694905810.1016/j.jaci.2015.12.1324 · doi ↗ · pubmed ↗
- 6Rhinosinusitis: establishing definitions for clinical research and patient care J Allergy Clin Immunol Meltzer EO Hamilos DL Hadley JA 15521211420041557786510.1016/j.jaci.2004.09.029PMC 7119142 · doi ↗ · pubmed ↗
- 7Phenotypes of chronic rhinosinusitis J Allergy Clin Immunol Pract Cho SH Hamilos DL Han DH Laidlaw TM 15051511820203238927510.1016/j.jaip.2019.12.021PMC 7696652 · doi ↗ · pubmed ↗
- 8Precision medicine in chronic rhinosinusitis with nasal polyps Curr Allergy Asthma Rep Avdeeva K Fokkens W 251820182957458610.1007/s 11882-018-0776-8PMC 5866836 · doi ↗ · pubmed ↗