Circular bioeconomy-driven carotenoid production by Agrococcus sp. NP24 using cheese whey byproduct: process optimization and bioactivity assessment
Nehad Noby, Fatma Elsayed, Mahmoud M. Agami, Nadia A. Soliman

TL;DR
A new method uses cheese whey to produce carotenoids with Agrococcus sp. NP24, showing good stability and bioactivity for food coloring.
Contribution
Optimized production of zeaxanthin monoester from cheese whey using Agrococcus sp. NP24 with bioactivity evaluation.
Findings
Maximum pigment yield of 0.0567 mg/mL achieved using 80% whey and optimized nutrients.
Pigment showed potent antioxidant activity (IC₅₀ of 6 μg/mL) and cytotoxicity against cancer cells.
Pigment was stable up to 50°C but lost 74% activity after 1 hour of daylight exposure.
Abstract
Growing interest in the circular economy has promoted the use of agri-food wastes as fermentable and readily available substrates for microbial cultivation, offering a sustainable and cost-effective strategy for natural pigment production. In this study, cheese whey was utilized as a nutritional substrate for pigment synthesis by an isolated strain identified as Agrococcus sp. NP24 (PQ097720.1). The work further aimed to characterize the produced pigment and evaluate its bioactivity. The culture medium was optimized using a Box–Behnken design (BBD). The carotenoid profile of the extracted pigment was analyzed by HPLC-DAD and LC–MS. Pigment stability was assessed across a range of pH values and temperatures, and its antimicrobial, antioxidant, and anticancer activities were examined. The pigment was identified as zeaxanthin monoester (C14:0). The maximum pigment yield (0.0567 mg/mL…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
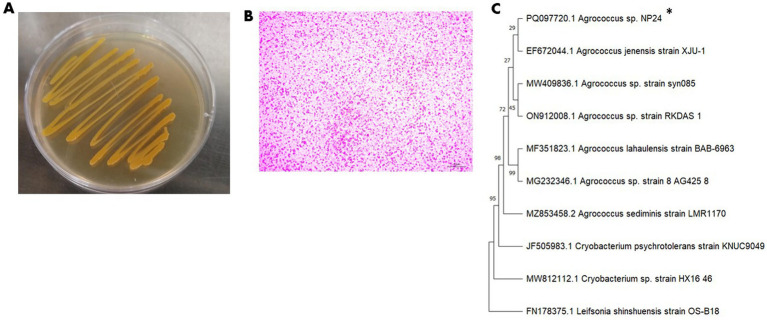 Figure 1
Figure 1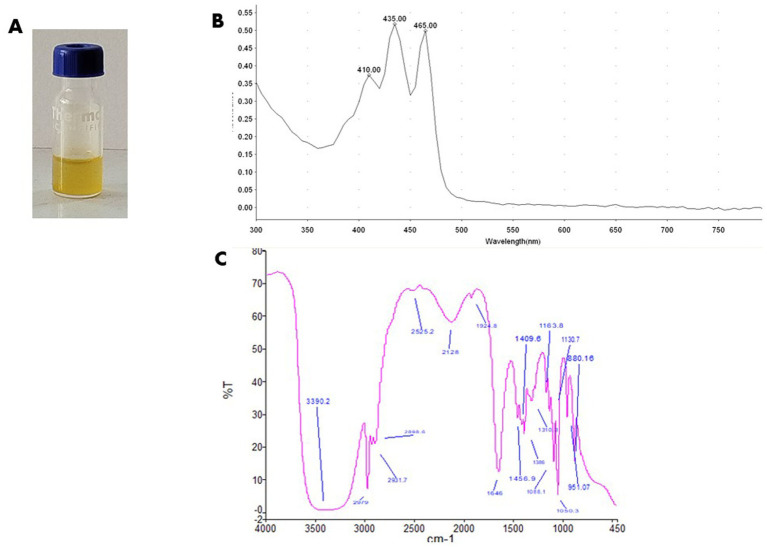 Figure 2
Figure 2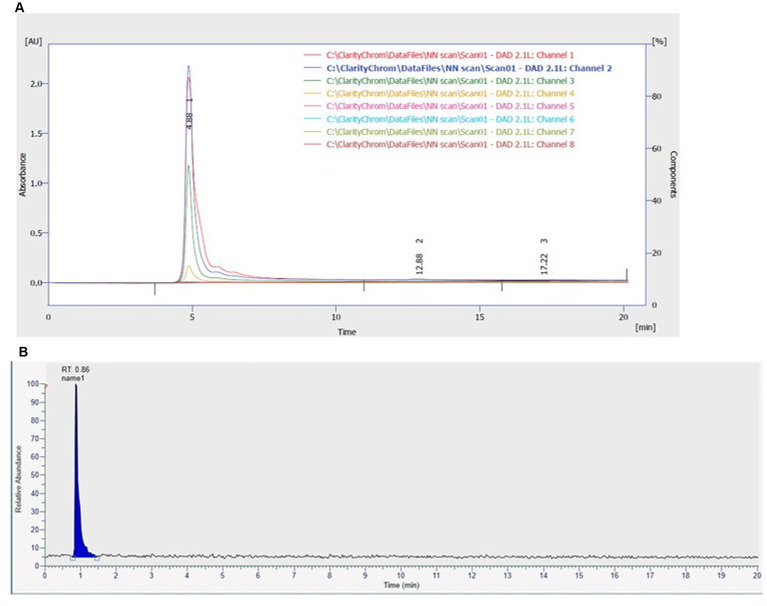 Figure 3
Figure 3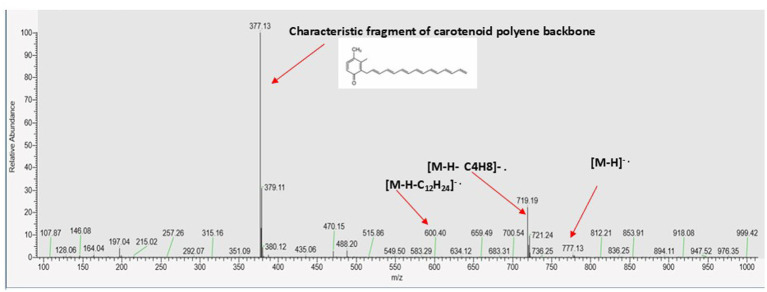 Figure 4
Figure 4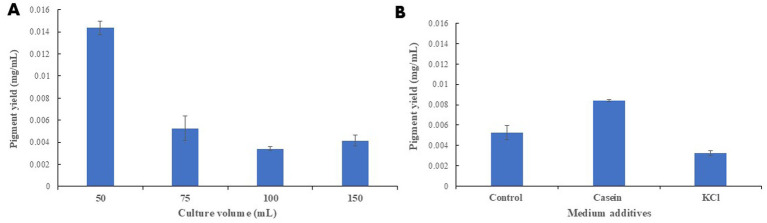 Figure 5
Figure 5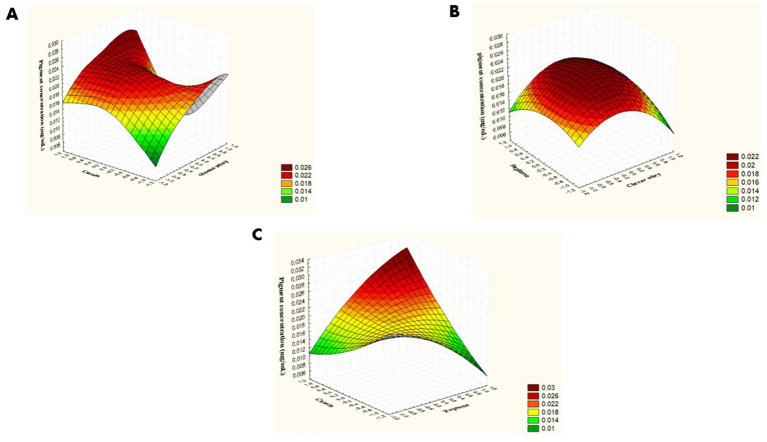 Figure 6
Figure 6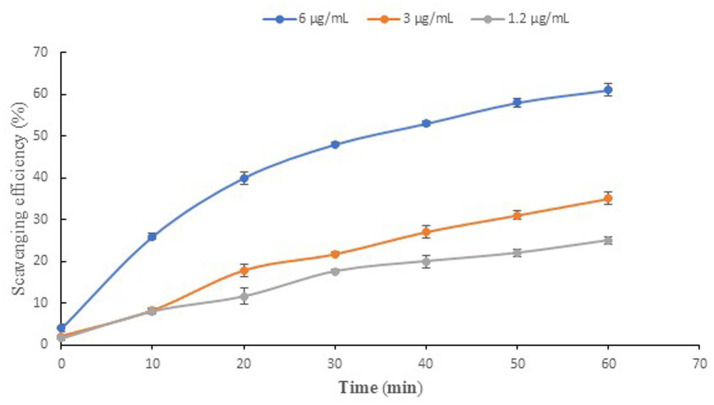 Figure 7
Figure 7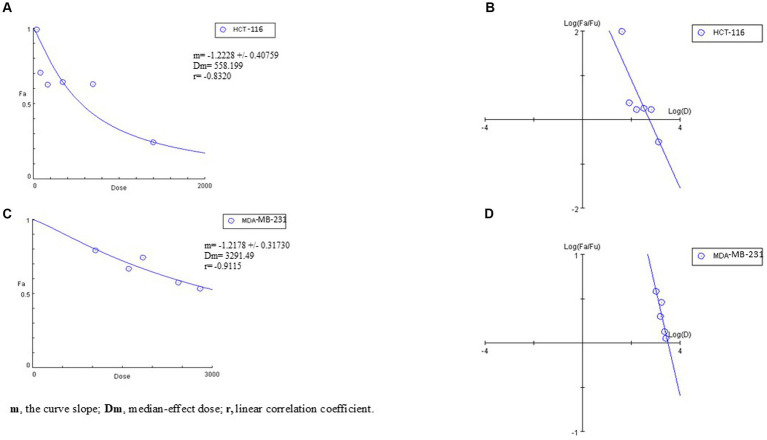 Figure 8
Figure 8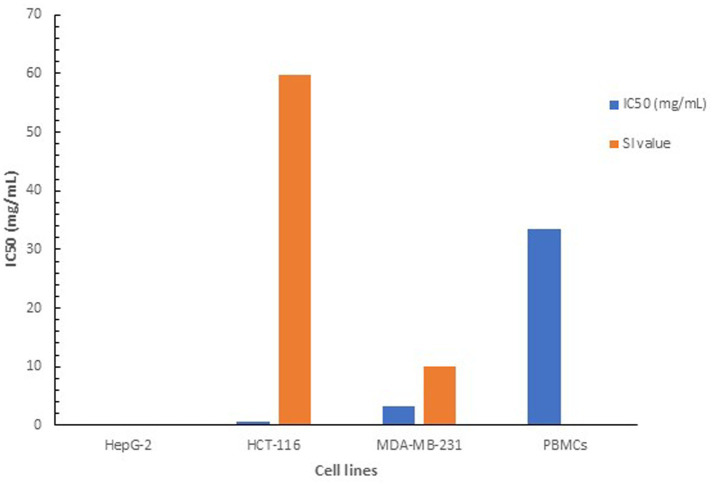 Figure 9
Figure 9| Peak number |
| [M-H].- | Carotenoid identification | Formula | |
|---|---|---|---|---|---|
| 1 | 4.23 | 435 | 777 | Zeaxanthin monoester | M-C14:0 |
| Fragment ions (m/z) | Formula | Reference |
|---|---|---|
| 719 [M-H-58].- | [M-H-C4H8] | |
| 600.4 [M-H-174].-_ | [M-H-C12H24] | |
| 377 [M-H-399].- | Characteristic fragment of polyene structure |
|
| pH | *Stability % | Temperature | *Stability % | Light effect | *Stability % |
|---|---|---|---|---|---|
| 3 | 310.5 ± 2.47 | 20 °C | 99.0 ± 0.98 | Day light | 26% ± 2.5 |
| 4 | 112 ± 0.24 | 30 °C | 115 ± 2.4 | ||
| 5 | 108 ± 1.48 | 40 °C | 104 ± 1.97 | ||
| 6 | 63.7 ± 1.69 | 50 °C | 102.5 ± 1.48 | ||
| 7 | 41.5 ± 1.48 | 60 °C | 62.5 ± 2.33 | ||
| 8 | ND | 70 °C | 52.0 ± 2.89 | ||
| 9 | ND |
| Variables | Pigment concentration (mg/mL) | |||||
|---|---|---|---|---|---|---|
| Trial number | X1 | X2 | X3 | Experimental | Predicted | Residual |
| 1 | 0 | 0 | 0 | 0.0220 | 0.0220 | -3.5E-18 |
| 2 | 0 | 0 | 0 | 0.0220 | 0.0220 | -3.5E-18 |
| 3 | 0 | -1 | 1 | 0.0176 | 0.0229 | −0.00539 |
| 4 | 0 | 1 | −1 | 0.0102 | 0.0048 | 0.00538 |
| 5 | −1 | −1 | 0 | 0.0175 | 0.0159 | 0.00152 |
| 6 | 0 | 0 | 0 | 0.0220 | 0.0220 | 3.5E−18 |
| 7 | 1 | 0 | -1 | 0.0192 | 0.0230 | −0.00386 |
| 8 | 0 | 1 | 1 | 0.0280 | 0.0228 | 0.00513 |
| 9 | 0 | −1 | −1 | 0.0260 | 0.0311 | −0.00514 |
| 10 | −1 | 0 | 1 | 0.0181 | 0.0142 | 0.00386 |
| 11 | −1 | 0 | −1 | 0.0116 | 0.0079 | 0.00361 |
| 12 | 1 | 0 | 1 | 0.0231 | 0.0267 | −0.00361 |
| 13 | 1 | 1 | 0 | 0.0150 | 0.0165 | −0.00153 |
| 14 | 1 | −1 | 0 | 0.0650 | 0.0560 | 0.009 |
| 15 | −1 | 1 | 0 | 0.020 | 0.029 | −0.009 |
| Term | Coefficients | Standard error | Upper 95.0% | ||
|---|---|---|---|---|---|
| Intercept | 0.022 | 0.004715 | 4.666246 | 0.005501 | 0.03412 |
| X1 | 0.006888 | 0.002887 | 2.385563 | 0.062731 | 0.014309 |
| X2 | −0.00661 | 0.002887 | −2.29031 | 0.070622 | 0.000809 |
| X3 | 0.002475 | 0.002887 | 0.857244 | 0.430472 | 0.009897 |
| X1X2 | −0.01313 | 0.004083 | −3.2145 | 0.02361 | −0.00263 |
| X1X3 | −0.00065 | 0.004083 | −0.15919 | 0.879746 | 0.009846 |
| X2X3 | 0.00655 | 0.004083 | 1.604189 | 0.169578 | 0.017046 |
| X1X1 | 0.002463 | 0.00425 | 0.579441 | 0.587417 | 0.013387 |
| X2X2 | 0.004913 | 0.00425 | 1.155941 | 0.299935 | 0.015837 |
| X3X3 | −0.00646 | 0.00425 | −1.52067 | 0.188823 | 0.004462 |
| Source | df | SS | MS |
| Significance |
|---|---|---|---|---|---|
| Regression | 9 | 0.001928 | 0.000214 | 3.2129 | 0.1058 |
| Residual | 5 | 0.000333 | 6.67E-05 | ||
| Total | 14 | 0.002262 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrobial Metabolism and Applications · Antioxidant Activity and Oxidative Stress · Algal biology and biofuel production
Introduction
1
Rising the world concern of environmental and health impacts of synthetic dyes has urged the need of natural, safe and ecofriendly alternatives. Traditionally, synthetic dyes have provided the sustained demand of colorants in food, pharmaceutical and textile industries. Their availability, low cost and stability have underpinned their preference for bright and stable coloration (Negi, 2025). However, recent studies have proved an association between synthetic food colorants and many health concerns (Barciela et al., 2023; Vieira Rubio et al., 2025; Alnuqaydan, 2024).
Numerous environmental fears have been also evolved regarding the extensive use of synthetic dyes. In addition to contaminating the human food chain, the leakage of dye effluents into water bodies reduces light penetration, thereby inhibiting photosynthetic activity in aquatic plants, disrupting the microbial balance and affecting soil fertility (Taha and Gouda, 2025; Ali, 2010). Furthermore, several synthetic colorants and their degradation products are known to be mutagenic, carcinogenic, or resistant to biodegradation, posing long-term ecological and health risks (Ali, 2010; Fried et al., 2022).
Although plant dyes are used as colorants in old human civilizations (Fried et al., 2022), their large-scale use is restricted by their low pigment yield, seasonal variations, and the competition for arable lands and water resources. Moreover, many plant dyes exhibit poor stability under light, temperature and pH variations (Lu et al., 2023), which limits their applicability in cosmetic and food products. The extraction of plant pigments is often inefficient and relies on a combination of organic solvents, thereby raising both economic and environmental concerns (Pati and Sarkar, 2022). These limitations highlight the necessity for potent alternative natural colorants.
Microorganisms have emerged as a potent sustainable source for natural biodegradable pigments. Several species of bacteria, yeast and fungi produce pigment to serve certain biological roles in microorganisms, such as the control of defense mechanisms and microbial interactions, which might explain their antimicrobial activity (Kirti et al., 2014). In other cases, they contribute the survival of the producer strains under harsh condition such as photooxidative damage caused by solar radiation due to their significant antioxidant qualities (Reis-Mansur et al., 2019). Microbial pigments can also be secreted conditionally in a stress-dependent manner, particularly under conditions such as high salinity or low temperature, to maintain the optimal fluidity and permeability of cellular membranes (Flegler and Lipski, 2021; Li et al., 2023).
Microorganisms produce pigments in different elegant shades with broad stability at different pH and temperatures (Saubenova et al., 2024). Various classes of microbial pigments have been identified, including flavins, carotenoids, monascin, anthraquinone and melanin, each characterized by distinct biosynthetic pathways and functional properties (Afroz Toma et al., 2023; Saubenova et al., 2024). Beyond their natural color, most of the tested pigments have demonstrated wide bifunctionalities like antioxidant and anti-inflammatory, making them appealing candidates as functional food additives and active ingredients in pharmaceutical products (Saubenova et al., 2024; Barreto et al., 2023).
Carotenoids represent the most widely distributed pigments in nature, produced in yellow, orange and red hues. They are produced in several microorganism-like bacteria, fungi, archaea and algae, in addition to their synthesis in some plants that appear in bright red-yellow colors. On the other side, animals like insects, crustaceans cannot synthesis them de novo while they mainly gain them through their diet (Maoka, 2020).
Structurally, most carotenoids harbor a polyene chain with nine conjugated double bonds arranged in a C40 skeleton produced from the condensation of two C20 geranyl-geranyl diphosphate chain in a head to tail manner (Siziya et al., 2022). Other less common carotenoids, synthesized in shorter (C30) or longer (C50) skeleton are produced in rare types of microorganisms. For instance, Staphylococcus aureus synthesizes a C30 golden-yellow pigment, known as staphyloxanthin, that helps in oxidative stress relief. On the other hand, C50 carotenoid known as bacterioruberin can be produced in some halophilic and psychrotolerant strains to maintain membrane functionalities (Flegler and Lipski, 2021; Pelz et al., 2005; Noby et al., 2023).
Despite the continuously expanded popularity of microbial pigments because of their safety, biodegradability and natural aesthetic colors, the sustainable production on the commercial scale has not been achieved yet. The nutritional medium cost is one of the major obstacles hindering the economic scale production of microbial pigments. To overcome this challenge, considerable efforts have been considered to employing low-cost substrates as nutrient sources for microbial growth (Grewal et al., 2022). Agricultural and food processing residues are rich in proteins, carbohydrates, vitamins, and minerals. However, the improper disposal of agricultural and food residues poses serious environmental challenges such as greenhouse gas emission, soil and water pollution and proliferation of pathogens (Yaashikaa et al., 2022). Thus, the valorization of these residues into bio-based compounds, particularly natural pigments, provides an environmentally sustainable approach to addressing the growing need for bio-colorants and promoting the circular economy framework (Banu et al., 2021).
The circular economy concept seeks to reduce waste and enhance resource efficiency by prolonging the lifecycle of materials inside production processes. Unlike the classic model of “take–make–dispose,” the circular approach highlights recycling, reuse, and bioconversion of waste into valuable products. The conventional linear production systems rely on refined substrates and energy-intensive waste management; however, circular bioeconomy-based fermentation integrates waste streams directly into the production cycle, thereby reducing capital investment associated with raw material purification, effluent treatment and disposal (Zaky et al., 2022). In addition, the use of minimally processed by-products such as whey or other nutrient-rich residues lowers the overall energy demand by eliminating upstream processing steps and enabling biological conversions into valuable products under mild operating conditions (Pescuma et al., 2015). Through direct waste reutilization, and energy recovery from residual biomass, circular bioeconomy strategies significantly decrease both capital expenditure and energy consumption per unit product, enhancing the economic feasibility and sustainability of fermentation-based bioprocesses. Moreover, its low carbon economy concept supports the potential of more sustainable and greener environment (Khanna et al., 2024; Mohan et al., 2016).
In the context of microbial pigment production, agriculture and food residues can be used as regenerative feedstocks for microbial fermentation (Vural Gursel et al., 2022). These solutions not only mitigate the environmental impact of waste disposal but also enhance economic sustainability by transforming waste streams into economically viable bioresources.
Many agri-food byproducts and wastes such as, corn steep liquor, bran, whey, molasses fruit pomace, seeds, peels, etc. have been utilized as possible fermentable substrates to produce bio-pigments using bacteria, yeast and fungi. For instance, Monascus ruber Tieghem IOC 2225 had produced red pigment using corn steep liquor in a higher titer compared to glucose based- medium under the same conditions (Hilares et al., 2018), supporting the high nutritional value of complex medium compared to synthetic one. In another study, glycerol by-product was applied as a carbon source by R. glutinis TISTR 5159 for carotenoids production (Saenge et al., 2011). Potato peels were used in solo as nutritional medium for bio-pigments production using Streptomyces genus train SO6 with a yield of 1.75 mg/g (Schalchli et al., 2021).
The increasing emphasis on the circular economy framework in natural pigment production has driven efforts to expand agri-food wastes employment as renewable and fermentable substrates for microbial cultivation, offering a sustainable and cost-effective route for bio-pigment synthesis. In this respect, the present study demonstrates the successful application of the circular economy concept through the optimization of carotenoid production by a psychrotolerant Agrococcus strain using cheese whey as a readily available, low-cost substrate.
Materials and methods
2
Materials
2.1
Peptone, yeast extract, MgSO_4,_ casein, glycerol and agar were obtained from Biolife Co., Italy. Universal primers for 16S rRNA were synthesized by Willowfort, UK. Standard Master Mix 2X was purchased from Promega, USA. Methanol, absolute ethanol, N-methyl-dimethyl sulphoxide (DMSO) and sodium hydroxide (NaOH), were obtained from Piochem (Piochem Laboratory Chemicals), Egypt. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) was obtained from Sigma-Aldrich Co., USA. Cheese whey was kindly provided from dairy processing department, faculty of Agriculture, Alexandria University.
Cell lines
2.2
Colorectal carcinoma HCT-116 cell line (RRID: CVCL_0291, ATCC^®^ CCL-247™), human hepatoblastoma HepG-2 cell line (RRID: CVCL_0027, ATCC^®^ Cat. No. HB-8065™), and human triple negative breast carcinoma (TNBC) MDA-MB-231 cell line (RRID: CVCL0062, ATCC HTB-26™). The three tested cell lines were obtained from Nawah Scientific, Egypt.
Growth and production media
2.3
Peptone–yeast (PY) medium was used for the isolation and routine activation of the bacterial strains. The medium consisted of 10 g L^−1^ peptone, 5 g L^−1^ yeast extract, and 5 g L^−1^ NaCl. Peptone–yeast agar (PA) medium was prepared by supplementing PY medium with 1.5% (w/v) agar.
A partially optimized medium obtained from the OVAT experiment consisted of 50% (v/v) cheese whey, 10 g L^−1^ peptone, 5 g L^−1^ casein, 5 g L^−1^ yeast extract, and 5 g L^−1^ MgSO₄.
The fully optimized medium consisted of 80% (v/v) cheese whey, 5 g L^−1^ peptone, 0.979 g L^−1^ casein, 5 g L^−1^ yeast extract, and 5 g L^−1^ anhydrous MgSO₄.
Methods
2.4
Isolation and identification of carotenoid producing strain
2.4.1
Isolation of pigmented bacterial strains was performed from soil samples. The collected soil samples were activated in 50 mL peptone yeast medium (PY) for 18 h., the activated culture was then serially diluted on Petone yeast agar medium (PA), followed by incubation for 48 h. at 20 °C. The developed pigmented colonies were picked up, purified and preserved at −20 °C on 40% glycerol. The selected bacterial strain was examined morphologically and further identified by molecular characterization using 16S rRNA gene sequencing (Weisburg et al., 1991). Genomic DNA was extracted from a pure culture of the selected strain using DNA miniprep kit (D3024, ZYMO), according to manufacturer’s instructions. The primer pair s F27/R1429 was used to amplify the 16 S rRNA universal gene (Weisburg et al., 1991). The resulting nucleotide sequence was analyzed via BLASTn tool provided by the NCBI. Finally, phylogenetic analysis was carried out using MGEA 9 software with closely related taxa to estimate the taxonomic affiliation of the isolate.
Pigment production, extraction and quantification
2.4.2
The selected bacterial strain was activated on PA plate. The grown colonies were activated in 20 mL PY medium and incubated at 20 °C for 18 h. A volume of 5% of the grown culture was inoculated into 100 mL fresh PY medium prepared in 500 mL Erlenmeyer flask, the culture was incubated at 150 rpm, 20 °C for 3 days.
At the end of the incubation period, the culture was harvested via centrifugation at 1,677 × g for 10 min. Cell pellets were then suspended and soaked in methanol until completed bleaching. The extract was clarified by centrifugation at 5,662 × g for 10 min. to remove cell debris. The methanolic extract was further stored in dark conditions at −20 °C for subsequent analysis. The extracted pigment was quantified using the percentage extinction coefficient (E₁%^1^cm) method as reported previously (Liaaen-Jensen and Jensen, 1971).
Structural characterization of the produced pigment
2.4.3
The spectral analysis of the extracted pigment was carried out through ultraviolet–visible (UV–Vis) spectrometry and Fourier Transform Infra-red (FT-IR) spectrometry. The UV–Vis spectrum of pigment methanolic extract was recorded using a Thermo Fisher Scientific spectrophotometer (Madison, WI, United States) over the range of 200–800 nm, with methanol as the blank. In FT-IR analysis, pigment methanolic extract was placed in a salt cuvette and analyzed using an FT-IR spectrometer (Perkin Elmer, Germany) over the range of 4,000–450 cm^−1^ to estimate the functional groups of pigment structure.
The extracted pigment was further analyzed using high-performance liquid chromatography (HPLC), coupled with diode array detection (LC-DAD) (AZURA Analytical (U) HPLC 862 bar System) and tandem mass spectrometry (LC–MS/MS) (Thermo Fischer Vanquish Horizon UHPLC system combined with TSQ Fortis™ Plus Triple Quadrupole Mass Spectrometer), a volume of 2 μL of the prefiltered pigment extract was injected into C_18_ reversed phase column (150 mm x 4 mm, 5 μm, Hypersil GOLD™, Lithuania) maintained at 30 °C. Separation of carotenoids was achieved using isocratic elution of 60% Acetonitrile in 0.1% Formic acid for 30 min.
Stability test of pigment extract
2.4.4
The stability of the extracted pigment was evaluated under various physical conditions, including temperature, pH, and exposure to sunlight. A known concentration of pigment methanolic extract was incubated under each tested condition for 1 h. The residual pigment concentration was determined spectrophotometrically at the maximum absorption wavelength (λ_max = 435 nm), and the stability percentage was calculated according to the following Equation 1:
Where, C1 and C0 are the concentrations of teste sample and control sample, respectively, the statistical significance was set at p < 0.05.
For temperature stability, the pigment extract was incubated at different temperatures, ranging from 20 °C to 70 °C for 1 h. A non-incubated sample was used as the control.
For pH stability, equal volumes (1:1, (v/v)) of the pigment methanolic extract and 100 mM buffer solutions were mixed and incubated at 4 °C for 1 h. The following buffers were used: acetate buffer (pH 3.0–5.0), phosphate buffer (pH 6.0–7.0), and Tris–HCl buffer (pH 8.0). The buffer volume was replaced with methanol in the control sample.
For light stability, the test sample was exposed to natural daylight for 1 h., while a non-exposed sample served as the control.
Optimization of whey-based medium for pigment production
2.4.5
A cheese whey–based medium was used as a cost-effective substrate for carotenoid production. The medium was formulated to contain 50% (v/v) cheese whey, 10 g L^−1^ peptone, 5 g L^−1^ yeast extract, and 5 g L^−1^ anhydrous MgSO₄. The culture medium (75 mL working volume) was prepared in 250 mL Erlenmeyer flasks. The initial pH was adjusted to 7.0, and incubation was carried out at 20 °C with agitation at 85 rpm for 3 days.
Pigment production was initially optimized using the one-variable-at-a-time (OVAT) approach, in which individual factors were varied while keeping all others constant. First, the effect of culture working volume on pigment yield was evaluated using different medium volumes (50, 75, 100, and 150 mL) prepared in 250 mL Erlenmeyer flasks. All cultures were incubated at 20 °C with agitation at 85 rpm for 3 days (statistical significance was set at p < 0.05).
The effect of medium composition was also examined through OVAT experiments. Specifically, potassium chloride (KCl) was supplemented into the production medium, and peptone was replaced with casein to qualitatively evaluate the influence of nitrogen source on pigment biosynthesis (statistical significance was set at p < 0.05).
Although the OVAT approach is useful for identifying significant individual factors, it does not account for possible interactions among variables. Therefore, the statistical optimization represented in response surface methodology will help in estimating the optimal concentration of the tested variables as well as the interactions among them (Reji and Kumar, 2022). In this context, three independent variables cheese whey (X1), peptone (X2) and casein (X3) were studied through Box Behnken Design (BBD) matrix to maximize carotenoid yield.
Each independent variable was represented in three coded levels, designated as −1 (low level), 0 (center level), and + 1 (high level), in a full design matrix of 15 experimental trials. The dependent variable (response) was expressed as carotenoids yield in (mg/mL). Each trial was prepared in 250 mL Erlenmeyer flasks with 50 mL medium, inoculated with 5% (v/v) of a 48 h preculture (~3 × 10^7^ CFU), and incubated at 20 °C with shaking at 85 rpm for 72 h.
The three independent variables were displayed by Equation 2:
Where Y represents the response, β0 is model intercept, β1, β2, and β3 are linear coefficients, β12, β13, and β23 are cross coefficients, while β11, β22, and β33 are quadratic coefficients.
For model validation and accuracy evaluation, the maximum yield predicted from the model was verified experimentally and the deviation percentage between predicted and achieved values was calculated.
Biological activity of pigment extract
2.4.6
Antimicrobial activity
2.4.6.1
The antimicrobial effect was assessed with 5 mg/mL of NP24 carotenoid dissolved in dimethyl sulfoxide (DMSO) against 0.5 McFarland of E. coli ATCC 25922, S. aureus ATCC 29213, and C. albicans ATCC 90028 using the agar well diffusion method (Perez, 1990). In this technique, the microbial strains were distributed separately on solidified medium plates, the wells are cut into solidified agar using a circular cork borer with 6 mm diameter. The wells were then filled with carotenoid solution (100 μL). Pure DMSO solvent served as the control test. The plates were first incubated at 4 °C for 1 h. to guarantee equal distribution of the extract, then incubated overnight at 37 °C. The zone of inhibition around the wells were compared with the control well.
Antioxidant activity
2.4.6.2
The antioxidant efficiency of Agrococcus sp. NP24 was assessed spectrophotometrically at 517 nm through free radical scavenging activity (RSA) using 2,2 Di Phenyl Picryl Hydrazil (DPPH) (Marinova and Batchvarov, 2011). Different concentrations of the extracted carotenoid were tested to estimate the IC50 value (6, 3, and 1.2 μg/mL). The total samples volume was brought into 250 μL with ethanol and mixed with 1 mL of 70 μM DPPH solution, the reaction was incubated at 20 °C in dark. The scavenging efficiency was measured at intervals along 1 h. incubation time. The sample volume was replaced with ethanol in the control sample. The scavenging efficiency was calculated according to Equation 3:
Anticancer activity
2.4.6.3
In vitro culture conditions
2.4.6.3.1
The three tested cell lines; HCT-116, HepG-2 cell line and MDA-MB-231 were cultured according to the American Type Culture Collection (ATCC) guidelines. Cultures were maintained using DMEM (Dulbecco’s modified Eagle’s medium), fortified with glucose (4.5 g/L) and L-glutamine (4 mM) (Lonza, United Kingdom). Complete culture medium preparation included the addition of fetal bovine serum (FBS) (MSE, France) at a concentration of 10% (v/v). Human peripheral blood mononuclear cells (PBMCs), as reference normal cells, were isolated from 5 mL of heparinized human blood, as described by Boyum (1968), using Histopaque^®^-1077 (Sigma–Aldrich, United States) density gradient separation medium, with the culture maintenance performed in complete Roswell Park Memorial Institute-1640 (RPMI-1640) medium (Lonza, United Kingdom). Incubation was carried out at 5 ± 1% CO2, 95 ± 5% humidity, and 37 ± 1 °C in a CO_2_ incubator (Nuaire, Germany).
Evaluation of the
in vitro anticancer effects of pigment extract
2.4.6.3.2
An MTT assay was adopted for this purpose as detailed by Vijayarathna and Sasidharan (2012). Cells were dispensed into 96-well plates (1 × 10^4^ cells/well) and further incubated under the previously described in vitro culture conditions for 24 h. Cells were treated with serial twofold dilutions of the carotenoid extract (5.6–0.043 mg/mL in complete culture medium) for 48 h after which the treatment medium was discarded followed by washing with saline solution (0.89%). Afterward, 100 μL of filtered MTT solution [0.5 mg/mL in phosphate buffer saline (pH 7.3–7.5) (Lonza BioWittaker, United Kingdom) was added]. Following a 2-h incubation, supernatants were carefully discarded, and 100 μL of dimethyl sulfoxide was used for solubilizing the precipitated formazan crystals before measuring the UV–Vis absorbance utilizing 96-well microplate spectrophotometer (Bio-Rad, Japan) at 490 nm.
An MTT assay was also employed to evaluate the extract’s effect on the cellular viability of normal PBMCs, following the method of Molaae et al. (2017). PBMNCs were dispensed as 100 μL (2 × 10^5^ cells) aliquots into a 96-well plate wells and treated with serial dilutions of the extract (33.5–4.2 mg/mL in RPMI-1640 complete culture medium). The assay was performed as described earlier under the same conditions adjusted to accommodate the suspended state of the PBMCs. Trials were in duplicate, and the anticancer activity was conveyed as the half-maximal inhibitory concentration (IC50) determined through regression analysis of dose–response curves using CompuSyn software (Chou and Martin, 2005).
Carotenoid extract selectivity
2.5
To assess the selectivity of the anticancer attribute of the carotenoid extract from Agrococcus sp., the selectivity index (SI) value was determined as the ratio of the extract IC50 value recorded for the PBMCs to the extract IC50 value recorded for each cell line, separately, as established by Nogueira and do Rosário (2010).
Results
3
Isolation and identification of carotenoid producing bacterial strain
3.1
Microbial isolation yielded a yellow-pigmented bacterial strain. On PA medium, the colonies appeared smooth, yellow, and rounded (Figure 1A), with a noticeable darkening of color after storage at 4 °C. The cells were Gram-positive and appeared in short irregular rods under the light microscope (Figure 1B).
Phenotypic and molecular identification of the isolated bacterial strain. (A) Colony morphology of Agrococcus sp. NP24 strain on PY medium. (B) Cell morphology under light microscope showing irregular rod shape cells. (C) Phylogenetic relatedness between NP24 strain sequence and closely related bacteria using neighbor joining method, Leifsonia shinshuensis is used as an outgroup. Accession number is indicated for each genus. Bootstrap values are represented as a percentage of 500 replicates.
The BLASTn analysis of 16S rRNA sequence showed high similarity to several closely related species of genus Agrococcus genus. The highest sequence identities (≥99%/) were observed with species Agrococcus jenensis (accession no. EF672044.1) A. lahaulensis (accession no. MF351823.1) and A. sediminis (accession no. MZ853458.2). Phylogenetic analysis of the 16S rRNA gene sequence and comparison with related sequences revealed that the isolate belongs to the genus Agrococcus within order actinomycetales. The strain was designated Agrococcus sp. NP24 (Figure 1C), and its 16S rRNA sequence was deposited in GenBank under the accession number PQ097720.1.
Identification and structural characterization of the produced pigment
3.2
NP24 pigment methanolic extract appeared as clear yellow liquid (Figure 2A). Pigment extract demonstrated a characteristic three fingered absorption spectrum distinctive to carotenoids class, with maximum absorbance at 435 nm (Figure 2B).
Spectral analysis of carotenoid extract from Agrococcus sp. NP24. (A) Methanolic extract of NP24 carotenoids. (B) UV/Vis scanning at 200–800 nm showing a characteristic three fingered chromatogram with maximum absorbance at 435 nm, FTIR spectrum showing the main functional group in the pigment.
FTIR spectral analysis (Figure 2C) revealed a characteristic absorption band at 1,650 cm^−1^, attributed to C=C stretching vibrations of alkenes, indicating the presence of aliphatic groups in the carotenoid extract. Additionally, the band at 1,456 cm^−1^ corresponds to CH₂ stretching, whereas the peaks observed at 1,386 cm^−1^ and 1,159 cm^−1^ are assigned to C–H and C–OH stretching vibrations, respectively.
NP24 carotenoid eluted as a single major component in HPLC-DAD, giving one sharp peak at RT = 4.89 min across the DAD channels (0–20 min run) with no additional detectable peaks. Consistently, MS analysis produced a single chromatographic peak (RT shown in the MS trace = 0.86 min) and a mass spectrum at the peak apex consistent with a carotenoid, supporting both identity and pigment purity (Figures 3A,B).
HPLC-DAD and MS chromatographic profiles of NP24 carotenoid. (A) HPLC-DAD chromatogram recorded across DAD channels showing a single dominant peak at RT = 4.89 min with no additional peaks detected across the 0–20 min separation. (B) UHPLC–MS chromatogram of NP24 showing a single peak at RT = 0.86 min.
The spectrum of NP24 carotenoid showed single peak on both HPLC-DAD and MS. The fragmentation pattern in negative ion mode identified the carotenoid as zeaxanthin monoester (M-C14:0) (Figure 4 and Tables 1, 2).
LC/ESI-MS–MS of NP 24 carotenoid extract in negative ionization mode.
Pigment stability assessment
3.3
Pigment stability was evaluated under different pH conditions, temperatures, and exposure to sunlight (Table 3). The pigment remained fully stable up to 50 °C but showed a noticeable decline at higher temperatures, with approximately 50% loss after 1 h. of incubation at 70 °C. The effect of pH revealed greater stability in acidic conditions (pH 3–5) compared to neutral and alkaline ones (pH 6–9). Under alkaline conditions, the pigment was totally instable with visible precipitation under short incubation time. Exposure to sunlight adversely affected the pigment extract, leading to severe degradation after 1 h. of daylight exposure, leaving a residual concentration of 26%.
Optimization of carotenoid production on whey-based medium
3.4
The effect of culture volume on pigment accumulation was tested via OVAT approach using different culture volumes. Reducing the working volume to 50 mL had significantly improved pigment yield with more than 3 times compared to higher volumes (statistical significance was set at p ≤ 0.05) (Figure 5A).
Optimizing pigment production via OVAT approach. (A) Testing different culture volumes on pigment yield. All trials were carried out in 250 mL Erlenmeyer flask. Seventy-five mL culture volume was set as the control trial. (B) Studying the effect of casein and KCl addition on pigment yield. The control trial consists of 80% (v/v) cheese whey, 1% (w/v) peptone, 0.5 g % (w/v), 0.5% (w/v) MgSO4. All trials were carried out in 250 mL Erlenmeyer flasks incubated at 20 °C for 3 days at 85 rpm. All trials were carried out in triplicates ± SE.
The effect of modifying the medium composition on carotenoids yield revealed a positive effect of casein while the addition of KCl had significantly reduced carotenoids production (Figure 5B). In addition, the effect of omitting MgSO_4_ and yeast extract additives had significantly lowered the total yield (data not shown). However, higher concentrations of MgSO_4_ and yeast extract negatively affected pigment accumulation (data not shown).
In this respect, a fixed concentration of MgSO4 and yeast extract was applied, while the concentrations of cheese whey (X1), peptone (X2) and casein (X3) were estimated via BBD in a 15-trial matrix at three encoded levels −1, 0, 1. Table 4 displays the designed matrix, the coded and actual level of each factor, and the response of each trial in terms of pigment concentration (mg/mL) quantified at 435 nm. Table 5 shows the coefficients, p-value and t stat of the tested variables.
Model significance was inferred from p-value (0.105) and F value (3.21) displayed by ANOVA analysis (Table 6). The small regression value (p = 0.056) proves the accuracy of the model in elucidating the relationship between the response and the involved independent variables. The value of R^2^ was 0.85, indicating a high degree of association between the predicted and experimental values.
The interactions among the variables are illustrated via Surface plots (Figure 6). The interactive effect between cheese whey and peptone is represented in Figure 6A. A significant antagonistic interaction was observed between whey and peptone (X1 × X2), as evidenced by a negative interaction coefficient (−0.013125) and a significant p-value (p = 0.02). In contrast, the interactions between whey and casein (X1 × X3) (Figure 6B) and between peptone and casein (Figure 6C) (X2 × X3) were not statistically significant (p = 0.879 and p = 0.169, respectively), indicating that pigment production in these cases was governed mainly by the individual effects of whey and peptone rather than by their interactions with casein.
Three-dimensional surface plots showing the interactive effect between the three independent variables on pigment concentration expressed in (mg/mL). (A) Whey and peptone; (B) whey and casein; (C) peptone and casein.
The exact concentrations of each variable and the maximum response were inferred from integrating the obtained results in a second order polynomial (Equation 4), expressing all forms of interactions.
Diffracting the polynomial equation revealed the optimal value for each variable to be 80% (v/v) cheese whey, 0.5 g % (w/v) peptone and 0.97 g % (w/v) casein to achieve a maximum pigment yield of 0.0565 mg/mL.
A verification experiment was carried out with the optimal values of the three tested value to assess the model accuracy. The model achieved an accuracy of 80.1% with an experimental value of 0.0453 mg/mL compared to a predicted value of 0.056 mg/mL.
Biological effect of Agrococcus sp. NP24 carotenoid extract
3.5
Antimicrobial effect
3.5.1
Upon comparing with control well (DMSO solvent), the NP24 carotenoid extract showed weak inhibitory activity with an inhibition zone of −8 mm (including 6 mm of agar well) against E. coli, while no detectable activity was observed against S. aureus and C. albicans.
Antioxidant effect
3.5.2
The three tested concentrations of NP24 carotenoids exhibited a positive correlation between carotenoid concentration and RSA%, showing increased radical scavenging activity with higher concentrations (Figure 7). The IC_50_ value was reached at a concentration of 6 μg/mL after 30 min of incubation.
Antioxidant effect of NP24 carotenoid extract using DPPH reagents. Three concentrations of NP24 carotenoid extract were tested along 1 h. Six μg/mL achieved IC50 after 30 min incubation.
In vitro cytotoxic/antiproliferative effects
3.5.3
The anticancer activity of the carotenoid pigment produced by Agrococcus sp. NP24 strain was evaluated using the MTT assay after 48 h of incubation. The dose–response curves and median effect plots (Figures 8A–D) illustrate the effects of serial dilutions of the methanolic pigment extract on the viability of the tested cell lines. The IC50 values of the carotenoid extract against the TNBC cell line MDA-MB-231 and the colorectal carcinoma cell line HCT-116 were 3.3 mg/mL and 0.56 mg/mL, respectively, whereas no cytotoxic effect was observed against the hepatoblastoma cell line HepG-2. Notably, the carotenoid extract exhibited an IC50 value of approximately 33.5 mg/mL against normal PBMCs, suggesting its potential selectivity and safety for anticancer applications.
Dose–response curves (A,C) and median-effect plots (B,D) generated by CompuSyn software to estimate the IC50 values of the carotenoid extract against HCT-116 and MDA-MB-231 cell lines.
The safety and selectivity of the NP24 carotenoid extract were further assessed by calculating the selectivity index (SI). The SI value measures the selective cytotoxicity of the investigated compound toward cancer cells relative to normal cells, and is calculated as the ratio of the IC50 value of the investigated compound, recorded for the PBMCs to the IC50 value recorded for the respective cell line. Accordingly, a higher SI value promises a more selective and hence safer therapeutic potential. The SI values for the MDA-MB-231 and HCT-116 cell lines were 10.2 and 59.8, respectively, indicating the high safety and selectivity of the carotenoid extract (Figure 9). According to Nogueira and do Rosário (2010), an SI value greater than 2 is generally required to ensure the safe application of a natural bioactive compound. Whereas Famuyide et al. (2019) suggested that an SI value above 1 can be considered satisfactory. The observed selectivity of the pigment extract remains valid even under more stringent selectivity criteria, including those proposed by Weerapreeyakul et al. (2012), who suggested an SI value ≥ 3, as well as more rigorous threshold of SI value ≥ 10 reported by Peña-Morán et al. (2016) for considering product safety.
IC50 and SI values of the carotenoid extract of Agrococcus sp. NP24 determined by the MTT assay on the three tested cell lines.
Discussion
4
The transition toward a circular bioeconomy has become a global priority to minimize waste generation and emphasizing the sustainable use of biological resources. Production of natural pigments from microorganisms using waste substrates, align well with the principles of circular bioeconomy since many agri-food wastes can be used as nutritional substrates for cultivating pigment producing strains. This work frame will not only guarantee a sustainable production of natural pigment in a cost wise approach, but will also reduce the environmental burden caused by the miss handling of these wastes.
The present study aligns well with the circular bioeconomy approach, as it addresses the production of microbial carotenoids using an isolated pigmented bacterial strain identified as Agrococcus sp. NP24, cultivated on a whey-based medium. Cheese whey is a major byproduct of dairy processing industries, representing a substantial volume in the dairy sector, with global estimates ranging between 160 and 190 million tons per year (Arshad et al., 2022). However, despite this large volume, more than 40% of whey remains under-utilized or disposed, leading to both environmental and economic burdens (Ahmed et al., 2023). Because cheese whey is rich in nutrients, such as soluble proteins, lactose, minerals, and vitamins, its direct disposal poses a significant environmental challenge. This is primarily due to its high organic load, which can deplete dissolved oxygen, disrupt aquatic ecosystems, and cause eutrophication. In this context, utilizing whey as a fermentation substrate for microbial pigment production offers a promising strategy for valorizing this byproduct while simultaneously mitigating its ecological impact.
Agrococcus genus is classified as Actinobacteria, order Micrococcales, family Microbacteriaceae (Whitman et al., 2012). A recent study carried out by Ding et al. (2022) showed novel bioactive metabolites from an isolated marine Agrococcus strain such as furan fatty acid and linear tetradepsipeptide, which highlights the importance of Agrococcus bacteria as a potential source of new bioactive compounds. Although previous studies reported the presence of carotenoid biosynthetic genes in Agrococcus pavilionensis RW1 and Agrococcus lahaulensis K22-21 genomes, including phytoene synthase, β-carotene ketolase (White et al., 2018), the research on Agrococcus carotenoids in terms of production optimization bioactivity and evaluation is quite limited.
Different species of Agrococcus genus produce a number of enzymes, enabling them to metabolize a wide range of carbon sources. Although β-galactosidase activity is not common among several spp. of Agrococcus genus (Behrendt et al., 2008), the isolated NP24strain was able to effectively use lactose sugar as the main carbon source (data not shown), which favored the use of whey, a lactose-rich byproduct, as a suitable and sustainable substrate for microbial pigment production.
Pigment production, like most fermentation processes, is strongly influenced by the composition of the growth medium as well as the cultivation parameters such as temperature and aeration. Therefore, optimization of these conditions is important for maximizing production efficiency. Aeration represents a crucial factor in pigment synthesis. High aeration ratio can enhance the activity of oxygen-dependent enzymes involved in pigment biosynthesis (e.g., desaturases, oxidases) (Musaalbakri et al., 2006). However, in certain microorganisms limited aeration can act as a stress factor, favoring alternative pathways involved in pigment accumulation (Sarian et al., 2016). There are two factors affecting oxygen transfer; the gas–liquid interfacial area (working volume) and the agitation speed. Aeration factor had significantly affected carotenoid synthesis in NP24 strain. Where a working volume of 50 mL incubated at low rpm (85 rpm) achieved the maximum yield.
High performance liquid chromatography is the main analytical technique applied for carotenoid identification. Reversed-phase high performance liquid chromatography (RP-HPLC) is the preferred method for separating carotenoids due to its excellent selectivity and simplicity of usage (Bijttebier et al., 2013). The spectral characteristics of carotenoid can be anticipated via photodiode array detector (PDA), while molecular weight and fragmentation pattern can be identified from mass spectrometric (MS) detectors. In the HPLC-DAD chromatogram, NP24 eluted as a single dominant, symmetrical peak at RT = 4.89 min, consistently observed across the DAD channels (200, 210, 230, 254, 320, 400, 450, and 500 nm), with no additional detectable peaks over the 0–20 min run. In the UHPLC–MS chromatogram, NP24 likewise produced a single chromatographic peak (RT = 0.86 min in the MS trace shown), and the mass spectrum acquired at the peak apex corresponded to the expected carotenoid molecular ion with characteristic fragment ions (Table 2), supporting compound identity. Together, the presence of a single peak in both DAD and MS (and the absence of secondary peaks) indicates that the isolated pigment is of high purity under the applied chromatographic conditions.
Esterified carotenoids are derivatives of xanthophyll that have one or both hydroxyl groups bound to fatty acids by ester bonds. This modification boosts the hydrophobicity and stability of carotenoids; it also contributes in protecting the polyene chain from oxidation and isomerization. Carotenoid esterification usually found in natural sources such as certain microorganisms, fruits and flowers, functioning as stabilized variant and storage form of free xanthophylls like lutein, zeaxanthin and violaxanthin. In mass spectrometric analysis, esterified xanthophylls usually show the molecular ion of the intact ester in addition to characteristic fragment ions corresponding to the cleavage of the fatty acid moiety (e.g., neutral loss of an acyl group) and/or the carotenoid backbone (Mercadante et al., 2017; Frassanito et al., 2008). The molecular ion peak of the separated peak [M-H]-^.^ was 777 corresponding to zeaxanthin monoester (C14:0). A fragment at m/z 719 [M-H-58]^-.^, represents a loss of small fraction, plausibly a short alkyl group (C_4_H_8_). The ester degradation sequence was confirmed by a fragment ion at m/z 600, which represents a larger cleavage of the alkyl group (C_12_H_24_). In addition, a characteristic ion peak of carotenoids polyene chain degradation was detected at m/z 377 (Rivera et al., 2014).
Carotenoids generally exhibit varied stability under different physical conditions, mainly due to their long polyene chains, which are highly susceptible to degradation by factors such as pH, temperature, and light. In general, carotenes display a higher temperature stability profile than xanthophylls, owing to their lack of oxygen-containing functional groups and their purely hydrocarbon backbones. However, the complete stability of the NP24 pigment up to 50 °C is consistent with the previously reported behavior of esterified xanthophylls, which typically exhibit greater structural stability and enhanced resistance to thermal degradation compared to their free counterparts (Moura et al., 2023). The thermal stability of carotenoids is an advantageous property for food applications, as it helps preserve their nutritional and functional value during processing and storage (Meléndez-Martínez et al., 2023). Like most carotenoids, NP24 pigment showed better stability at acidic conditions compared to neutral one by increasing the absorbance by about 310, 112, 108% at pH 3, 4 and 5, respectively (Cheng et al., 2023). The enhancement in absorbance can be explained by increased pigment solubility and absence of hydroxide ions. However, the pigment completely precipitated at alkaline conditions which can be attributed to the hydroxide-induced oxidation and decreased solubility (Cheng et al., 2023).
The low photostability of the tested carotenoid matches with the well-known photosensitivity of carotenoids. Where, the conjugated double bond system makes them highly reactive toward free radicals generated by light, causing isomerization and photooxidation under light exposure (Aslam et al., 2021).
The biological activity of natural pigments such as antioxidant, anticancer, antimicrobial usually helps in understanding their possible applications in pharmaceutical, nutraceutical, or food industries. NP24 zeaxanthin monoester extract showed anticancer activity against two cancer cell lines with variable potency. The production of carotenoids with anticancer properties by various bacterial taxa has been documented in several studies. A previous study by Choi et al. (2014) reported the synthesis of deinoxanthin by the radioresistant actinomycete Deinococcus radiodurans, with IC₅₀ values of 59, 61, and 77 μM against the HepG-2, HT-29, and PC − 3 cell lines, respectively. Similarly, (Afra et al., 2017) reported the anticancer activity of red carotenoid-related pigments from a marine Arthrobacter sp. against the oesophageal cancer cell line KYSE30, with an IC₅₀ value of 23.5 μg/mL. The same pigment also exhibited antioxidant potential in the DPPH assay, with an IC₅₀ value of 4.5 mg/mL. Later, Tapia et al. (2021) demonstrated that the extremophilic actinomycete Deinococcus sp. UDEC-P1, isolated from Patagonia, produced deinoxanthin with selective anticancer activity against the human osteosarcoma cell line Saos-2. In contrast, Arthrobacter sp. UDEC-P1, isolated from Antarctica, synthesized other carotenoids that exhibited selective antiproliferative effects against the cancer cell lines MCF-7 (human breast adenocarcinoma), Saos-2 (human osteosarcoma), and Neuro-2a (mouse neuroblastoma).
In this work, human cancer cell lines were selected to represent clinically distinct carcinoma models to evaluate the broad-spectrum anticancer potential of the bacterial carotenoids across diverse types of carcinomas. Considering that the anti-colorectal activity previously reported in the literature for bacterial carotenoids was directed mostly to HT-29 human colorectal carcinoma cells (Choi et al., 2014); however, more cell lines modeling distinct genetic backgrounds of colorectal carcinoma remain unexplored; hence, in the current study it was aimed to investigate the anticancer activity of Agrococcus sp. carotenoids against HCT-116 cells known to be a highly proliferative, tumorigenic, and poorly differentiated epithelial subtype of colorectal carcinoma, harboring a key mutation in codon 13 of KRAS proto-oncogene (Rajput et al., 2008) rendering HCT-116 cell line commonly used model in drug screening studies. Similarly, MDA-MB-231 cells were selected to model a highly dedifferentiated, mesenchymal, basal subtype of human TNBC which lacks the expression of estrogen, progesterone, as well as HER-2 receptors; thus, is unresponsive to hormonal and HER-2 targeted therapies; consequently, MDA-MB-231 cells represents a highly aggressive, metastatic human breast adenocarcinoma associated with the high recurrence rates and poor prognosis (Chavez et al., 2011). In the same context, HepG-2 cells, earlier described as hepatocellular carcinoma, were selected to represent a well-differentiated human hepatoblastoma model (López-Terrada et al., 2009). Interestingly, the anticancer activity reported in the literature for carotenoids against the colorectal carcinoma HCT-116 cell line is mostly related to plant and algal carotenoids not bacterial (Baeza-Morales et al., 2024; Hormozi and Baharvand, 2024). Contextually, the current work is among the few reports accentuating the anti-HCT-116 activity of bacterial carotenoids which have been more commonly reported for their anti-colorectal activity against HT-29 cell lines as for the study by Choi et al. (2014) on deinoxanthin from Deinococcus radiodurans against HT-29 cell line.
The NP24 pigment showed selective weak antimicrobial activity only against Escherichia coli. Such species-specific variation has also been reported in other carotenoid studies (Naisi et al., 2023), where antimicrobial activity depended more on cell membrane interactions, pigment composition and solubility than on bacterial Gram type (Saubenova et al., 2024). Carotenoids are generally described as antioxidants and photoprotective agents, not strong antimicrobial agents. A previous study on fucoxanthin carotenoids reported only weak antibacterial effects with a median inhibition zone of 10.5 and 8.5 for Gram negative and Gram positive, respectively at a concentration of 25 μg (Karpiński and Adamczak, 2019). The weak and selective inhibition suggests that the pigment’s primary biological role may not be antimicrobial but rather associated with antioxidative mechanisms.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Afra S. Makhdoumi A. Matin M. Feizy J. (2017). A novel red pigment from marine Arthrobacter sp. G 20 with specific anticancer activity. J. Appl. Microbiol. 123, 1228–1236. doi: 10.1111/jam.13576, 28862783 · doi ↗ · pubmed ↗
- 2Afroz Toma M. Rahman M. H. Rahman M. S. Arif M. Nazir K. N. H. DufosséL. (2023). Fungal pigments: carotenoids, riboflavin, and polyketides with diverse applications. J. Fungi 9:454.10.3390/jof 9040454 PMC 1014160637108908 · doi ↗ · pubmed ↗
- 3Ahmed T. Sabuz A. A. Mohaldar A. Fardows H. M. S. Inbaraj B. S. Sharma M. . (2023). Development of novel whey-mango based mixed beverage: effect of storage on physicochemical, microbiological, and sensory analysis. Foods 12:237. doi: 10.3390/foods 12020237, 36673328 PMC 9858226 · doi ↗ · pubmed ↗
- 4Ali H. (2010). Biodegradation of synthetic dyes—a review. Water Air Soil Pollut. 213, 251–273. doi: 10.1007/s 11270-010-0382-4 · doi ↗
- 5Alnuqaydan A. M. (2024). The dark side of beauty: an in-depth analysis of the health hazards and toxicological impact of synthetic cosmetics and personal care products. Front. Public Health 12:1439027. doi: 10.3389/fpubh.2024.1439027, 39253281 PMC 11381309 · doi ↗ · pubmed ↗
- 6Arshad U.-E.-T. Hassan A. Ahmad T. Naeem M. Chaudhary M. T. Abbas S. Q. . (2022). A recent glance on the valorisation of cheese whey for industrial prerogative: high-value-added products development and integrated reutilising strategies. Int. J. Food Sci. Technol. 58, 2001–2013.
- 7Aslam S. Ahmad M. Riaz M. (2021). “Stability of carotenoids” in Carotenoids: structure and function in the human body. eds. Zia-Ul-Haq M. Dewanjee S. Riaz M. (Cham: Springer International Publishing).
- 8Baeza-Morales A. Medina-Garcia M. Martinez-Peinado P. Pascual-Garcia S. Pujalte-Satorre C. Lopez-Jaen A. B. . (2024). The antitumour mechanisms of carotenoids: a comprehensive review. Antioxidants 13:1060. doi: 10.3390/antiox 13091060, 39334719 PMC 11428676 · doi ↗ · pubmed ↗