Assay of Ds2 Transposition Activity in the Maize W22
Harpreet Kaur, Matthew Bacchus, Lilly Horowitz, Caroline Larow, Vedant Rawat, Dafang Wang

TL;DR
The study examines the transposition activity of Ds2 elements in the maize W22 line, finding that most are immobile and only one shows possible somatic transposition.
Contribution
The study identifies structural integrity and DNA methylation as insufficient predictors of Ds2 transposition variation in W22.
Findings
Most Ds2 elements in W22 are immobile, with only Ds2-23 showing possible somatic transposition.
Ac transposase reduces methylation but does not fully explain transposition activity variation.
Structural integrity and DNA methylation alone cannot predict Ds2 transposition in W22.
Abstract
Ds elements are non-autonomous DNA transposons requiring Ac transposase for mobility. From the 39 Ds2 elements reported in B73, we identified 38 Ds2 homologous elements in the W22, 30 of which retained intact terminal inverted repeats and target site duplications. PCR-based excision assays on 19 loci in embryos and leaves revealed the majority are immobile; only Ds2-23 showed suggestive somatic transposition. Targeted bisulfite sequencing revealed that while Ac reduces methylation, absolute levels remain stable and do not correspond to the varied transposition activities. This suggests structural integrity or DNA methylation alone does not predict Ds2 transposition variation in W22.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
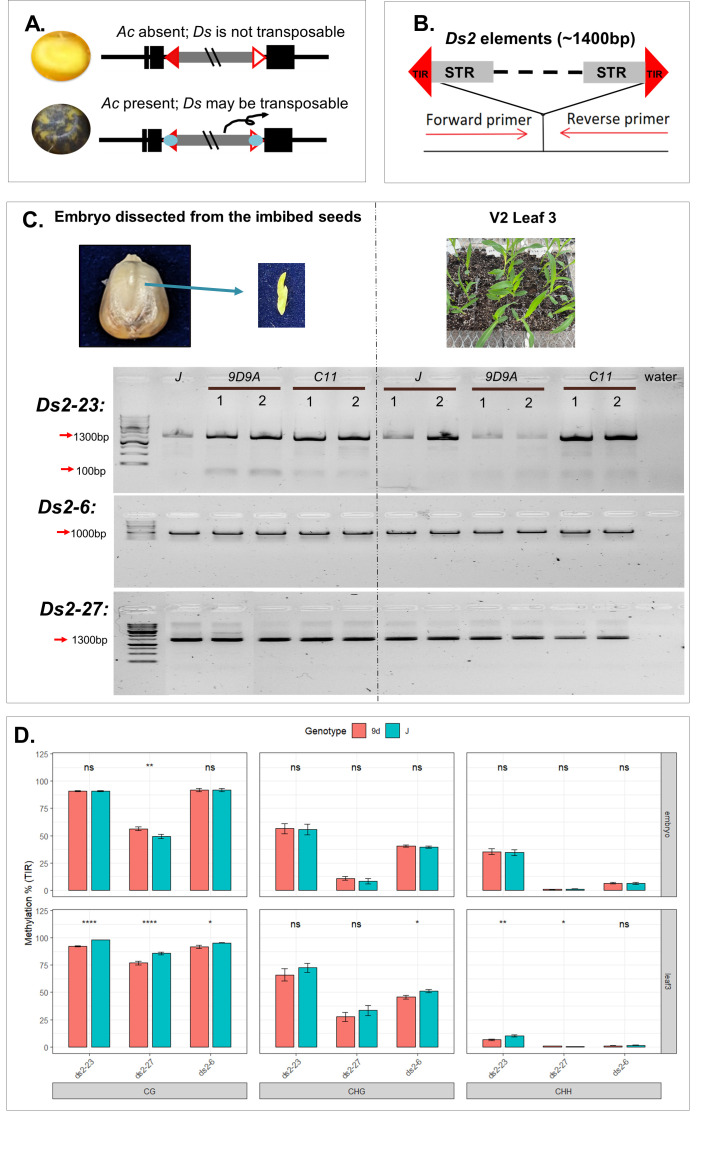 Figure 1
Figure 1|
| ||
|
Ds2-6_473-720 (Ds2-6 subterminal 1-220bp) |
AAATGTTGTTTAGTATTATTTGTTATTTTTAGATTTAGG |
ATTTCATAATATAATTTTACCRAACAAAAATACC |
|
(Ds2-23 subterminal 1-207bp) |
GAGGAGTTATGGGTTGTGGAGGTTGT |
RCCTACTCTCTCCCTATCTCTCAACC |
|
(Ds2-27 subterminal 1-270bp) |
TGTATAATTAGGGATGAAAGTAGGATG |
ATCTAAATTACAACAAACTATATTAAAATAATCCC |
- —U.S. National Science Foundation (United States)https://ror.org/021nxhr62
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChromosomal and Genetic Variations · Plant Molecular Biology Research · Genomics and Phylogenetic Studies
Description
Transposable elements (TEs) are mobile genetic sequences first identified in maize by Barbara McClintock (McClintock, 1950; 1951; 1956). In maize, TEs account for approximately 85% of the genomic DNA (Schnable et al, 2012). The Activator/Dissociation ( Ac/Ds ) family includes the autonomous Ac element, which encodes a transposase that catalyzes the excision and reintegration of non-autonomous Ds elements. Ds elements arise from Ac through internal deletions or substitutions (Fedoroff et al, 1983; Conrad et al, 2007; Vollbrecht et al, 2010; Lazarow et al, 2013). Genome-wide analysis of the B73 reference genome previously identified nearly 900 Ds and Ds -like elements, classified into four subfamilies: Ds1 , Ds2 , Ds3L , and *Ds4L * (Du et al, 2011).
Ds transposition is driven by the Ac transposase. The terminal inverted repeats (TIRs) and subterminal repeats (STRs) of Ds are conserved sequences essential for transposase recognition and binding (Kunze et al, 1989). Cytosine methylation, particularly within GC-rich STRs, has been implicated in regulating Ds activity (Len et al 1992; Ott et al, 1992; Brutnell, et al, 1994; Wang et al, 1996; 1998). Among the subfamilies, Ds1 and Ds2 are predicted to retain mobility due to conserved STR motifs harboring Ac -binding sites (Du et al, 2011).
Here, we focused on 39 Ds2 elements annotated in B73 that retain intact STR sequences and possess the potential to transpose. We examined these elements in the W22 maize background, a line historically used for studying Ac/Ds and Mutator ( Mu ) families (Lisch, 2009; Jiang, et al, 2019). The objectives of this study were to: (i) identify W22 homologous elements of B73 Ds2 elements, (ii) assess their transposition capacity in embryos and young leaves, and (iii) evaluate whether variation in DNA methylation within STR regions is associated with the presence of Ac and transposition activity.
We performed BLAST searches to identify homologous elements in the W22 inbred line corresponding to the 39 Ds2 elements previously annotated in the B73 reference genome (Schnable et al, 2012; Springer, et al, 2018). Of these, 38 homologous elements were recovered. Thirty of these elements retained the characteristic TIRs and were flanked by TSDs (Supplemental Data 1). These 30 characteristic elements in W22 shared high sequence similarity with their B73 counterparts (>90%), with length differences ranging from 0% to 45% (Supplemental Data 1). These elements represent only the W22 homologous elements of B73 Ds2 ; W22-specific Ds2 elements were not included, as a genome-wide annotation was not conducted in this study.
To assess Ds2 mobility, we utilized two classes of genotypes: Ac -present lines ( p1-vv9D9A and p1-wwC11 ) and Ac -absent controls (J and W22). All lines except W22 (specifically p1-vv9D9A , p1-wwC11 , and J) carry the r1-m3 allele, a specialized "tester" for Ac activity containing a Ds insertion at the R1 locus (Kermicle, 1970; Federoff et al., 1983). To ensure genomic consistency, these tester stocks were backcrossed to the W22 over multiple generations. Despite the predominantly W22 background, residual donor DNA from the tester lines likely persists, explaining the observed sequence heterogeneity in specific genomic regions. In the Ac -present lines, the Ds element was excised from the R1 gene by the Ac transposase, restoring anthocyanin pigmentation and producing purple spots in the aleurone layer of the endosperm (Figure A), which is to confirm that the Ac transposase was active and competent to catalyze Ds excision at independent loci within the W22 background.
For each Ds2 element tested in W22, primers were designed from the ±500 bp flanking sequences (Figure B). In Ac
- backgrounds (W22, J), where Ds2 elements are immobile, PCR amplification produced only the expected large insertion bands. In Ac
- lines, transposition was expected to generate smaller bands following excision. We tested germinating embryos and young leaves (V2 stage), representing early developmental stages where Ac/Ds transposition is typically active.
Of the 38 homologous elements identified, 31 were selected for testing, including 27 with intact TIRs/TSDs and four lacking one or both features. PCR amplification yielded the expected insertion bands for 19 of these elements. Among them, only Ds2-23 generated smaller fragments consistent with somatic excision. These putative excision products were consistently observed across biological replicates in Ac+ backgrounds but were absent in Ac– controls in both embryonic and leaf tissues (Figure C). While these results suggest somatic excision of Ds2-23 , low band intensity prevented the recovery of sufficient PCR product for Sanger sequencing. Consequently, the "active" status of Ds2-23 remains tentative pending future cloning experiments to confirm the excision footprint sequences. The remaining 18 elements showed no observable bands corresponding to excision footprints; therefore, we conclude they are either immobile or possess negligible transposition activity within the tested tissues.
To assess whether cytosine methylation correlates with transposition potential, we performed targeted bisulfite sequencing of three Ds2 elements: the potentially active Ds2-23 and two inactive elements ( Ds2-6 and Ds2-27 ) (Supplementary Data 2). Methylation in CG, CHG, and CHH contexts was quantified across the first 250 bp of each element, including sub-terminal regions (STRs) expected to be methylation targets (Kunze et al., 1989; Len et al 1992; Ott et al, 1992; Brutnell, et al, 1994; Wang et al, 1996; 1998). This region includes a varying number of cytosine sites (e.g., 23 CG sites for all the tested Ds2; 8 CHG sites in Ds2-23 ; 12 in Ds2-27 ; and 4 in Ds2-6; 9 CHH sites in Ds2-23 ; 7 in Ds2-27 ; and 6 in Ds2-6 ).
The average DNA methylation levels and standard errors are presented in Figure D. In the embryo, the presence of Ac did not significantly alter cytosine methylation levels, except for Ds2-27 . In leaves, methylation significantly decreased in the presence of Ac in the CG and CHH contexts for Ds2-23 and Ds2-27 , and in the CG and CHG contexts for Ds2-6 . However, despite these statistically significant reductions, the absolute DNA methylation levels remained notably stable across all genotypes and do not induce transposition activities in Ds2-27 and Ds2-6 . This observation mirrors the behavior of the cryptic Ac -homologous sequence, cAc-11 , which shares over 90% homology with known Ac sequences and possesses the characteristic 11-bp terminal inverted repeats and 8-bp target site duplications of the Ac/Ds family (Len et al, 1992). In that system, the presence of Ac failed to eliminate hypermethylation or recover the activity of the cryptic element. However, that study lacked quantitative analysis to determine whether methylation was also significantly reduced yet remained above a functional threshold, as observed in our results. Interestingly, while the potentially mobile Ds2-23 also showed a decrease in DNA methylation, its absolute methylation levels remained higher than those of the inactive Ds2-27 and Ds2-6 . This suggests that factors beyond DNA methylation, such as local chromatin environment or position effects, may be involved in the immobilization of Ds2-27 and Ds2-6 or the activation of Ds2-23 .
Our results demonstrate that most Ds2 elements in B73 have homologous elements in W22 with intact terminal inverted repeats (TIRs) and target site duplications (TSDs). Despite this structural integrity, the W22 genome maintains stable silencing across most Ds2 loci, likely through DNA methylation. While the presence of Ac induces statistically significant reductions in methylation, these shifts appear biologically insufficient to restore mobility. Of 19 loci tested, only Ds2-23 showed evidence of somatic transposition. Notably, this activity did not correlate with lower methylation levels compared to other lines. Consequently, neither structural integrity nor DNA methylation levels serve as reliable predictors of Ds2 transposition, suggesting that regulation is likely enforced by additional mechanisms such as the local chromatin environment or position effects.
Methods
Genome Searches Sequences and genomic locations of all Ds2 elements were obtained from Du et al. using the B73 reference genome. Because the Ac lines used in this study were originally derived from the W22 background, we performed BLAST searches of Ds2 elements in the W22 genome to obtain their sequences along with 500 bp of upstream and downstream flanking regions. Primers were designed from these flanking regions using the NCBI Primer-BLAST tool. Three primer pairs were designed and tested for each Ds2 element.
Plant Growth Maize lines p1-vv9D9A, p1-wwC11, and the wild-type line J were obtained from Dr. Thomas Peterson’s laboratory at Iowa State University. The W22 line was obtained from the Maize Genetics Cooperation Stock Center. Seeds were germinated in Petri dishes containing two sheets of paper towel moistened with 3 mL of water, followed by 500 µL of water every 24 h. Germination was carried out at 28 °C in an incubator, and embryos were dissected from the imbibed seeds after 41 h of incubation. For whole-plant growth, seeds were germinated in small pots containing Premier B10281RG ProMix in a greenhouse maintained at 25 °C. The entire third leaf was collected at the V2 stage.
DNA Extraction Genomic DNA was extracted from 41-hour imbibed embryos and from third leaves at the V2 stage. Two to three independent plants of the same genotype were processed using the Quick-DNA™ Plant/Seed Miniprep Kit (Zymo Research, D6020) following the manufacturer’s protocol. DNA quality was assessed using ZmTUB (Zm00001d010275) primers, and all samples were confirmed to be PCR-amplifiable.
PCR Somatic transpositions in lines carrying active Ac were detected by PCR using PCR Master Mix (Sydlabs, MB067-EQ2B). Each reaction contained primers specific to the Ds2 flanking sequence (primer sequences listed in “Reagents”), with nuclease-free water serving as a negative control. PCR amplification conditions were: initial denaturation at 95 °C for 30 s; 35 cycles of 95 °C for 30 s, 55 °C for 30 s, 72 °C for 30 s; and a final extension at 72 °C for 5 min. Amplified products were separated on 1% agarose gels and visualized using the GelDoc Go Imaging System (Bio-Rad).
Targeted Bisulfite Sequencing For each genotype and tissue type, genomic DNA from two independent biological replicates was sent to CD Genomics (Shirley, NY) for library preparation and sequencing (primer sequences listed in “Reagents”). Primers targeting the subterminal regions (1–250 bp) of the TIR and STR sequences were used. Methylation data were averaged across replicates for presentation in the figure.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brutnell TP Dellaporta SL 199491 Somatic inactivation and reactivation of Ac associated with changes in cytosine methylation and transposase expression.Genetics 13810016-673121322510.1093/genetics/138.1.2138001788 PMC 1206132 · doi ↗ · pubmed ↗
- 2Du Chunguang Hoffman Andrew He Limei Caronna Jason Dooner Hugo K 2011121 The complete Ac/Ds transposon family of maize BMC Genomics 1211471-216410.1186/1471-2164-12-588PMC 326021022132901 · doi ↗ · pubmed ↗
- 3Fedoroff N. Wessler S. Shure M. 1983111 Isolation of the transposable maize controlling elements Ac and Ds Cell 3510092-867423524210.1016/0092-8674(83)90226-x 6313225 · doi ↗ · pubmed ↗
- 4Kermicle J L 197021 SOMATIC AND MEIOTIC INSTABILITY OF R -STIPPLED, AN ALEURONE SPOTTING FACTOR IN MAIZE Genetics 6421943-263124725810.1093/genetics/64.2.24717248480 PMC 1212400 · doi ↗ · pubmed ↗
- 5Kunze R. Stochaj U. Laufs J. Starlinger P. 198761 Transcription of transposable element Activator (Ac) of Zea mays L The EMBO Journal 660261-41891555156310.1002/j.1460-2075.1987.tb 02400.x 16453772 PMC 553524 · doi ↗ · pubmed ↗
- 6Kunze R. Starlinger P. 1989111 The putative transposase of transposable element Ac from Zea mays L. interacts with subterminal sequences of Ac.The EMBO Journal 8110261-41893177318510.1002/j.1460-2075.1989.tb 08476.x 2555157 PMC 401433 · doi ↗ · pubmed ↗
- 7Len Jun-Yi Henry Sun Y. Lai Yiu-Kay Chen Jychian 199261 A maize cryptic Ac-homologous sequence derived from an Activator transposable element does not transpose Molecular and General Genetics MGG 23330026-892541141810.1007/bf 002654381320187 · doi ↗ · pubmed ↗
- 8Mc Clintock Barbara 195061 The origin and behavior of mutable loci in maize Proceedings of the National Academy of Sciences 3660027-842434435510.1073/pnas.36.6.344PMC 106319715430309 · doi ↗ · pubmed ↗