Discovery of two new Phaeoclavulina species (Basidiomycota, Gomphaceae) from high-elevation habitats in the Qinghai–Xizang Plateau

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
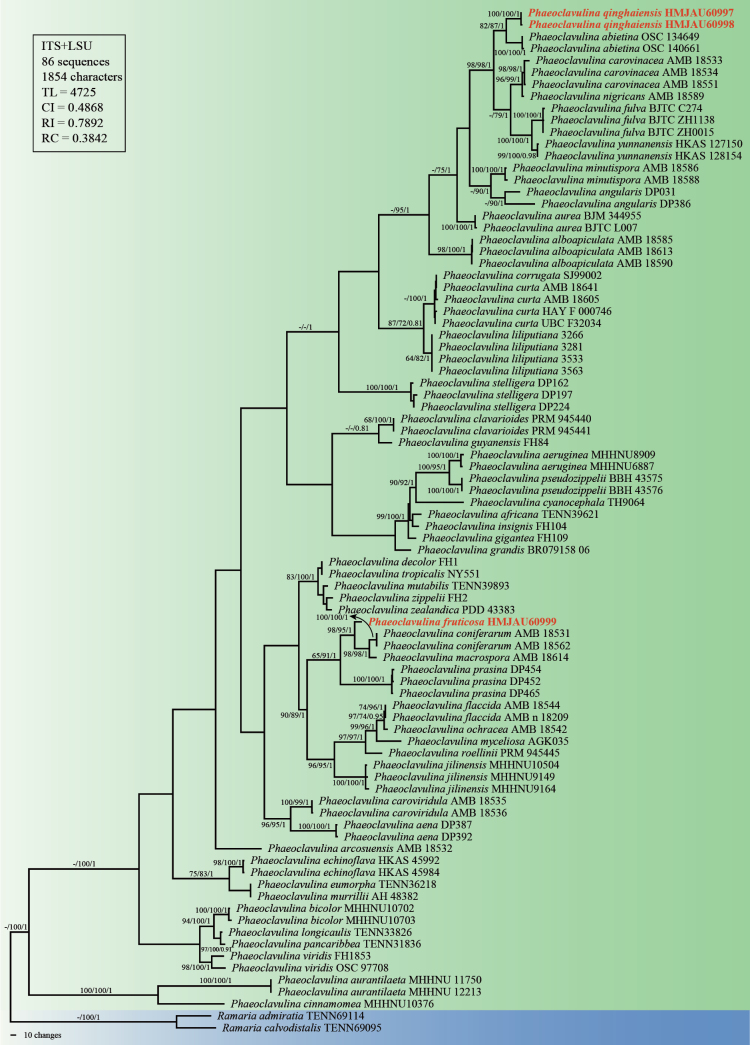 Figure 1
Figure 1 Figure 2
Figure 2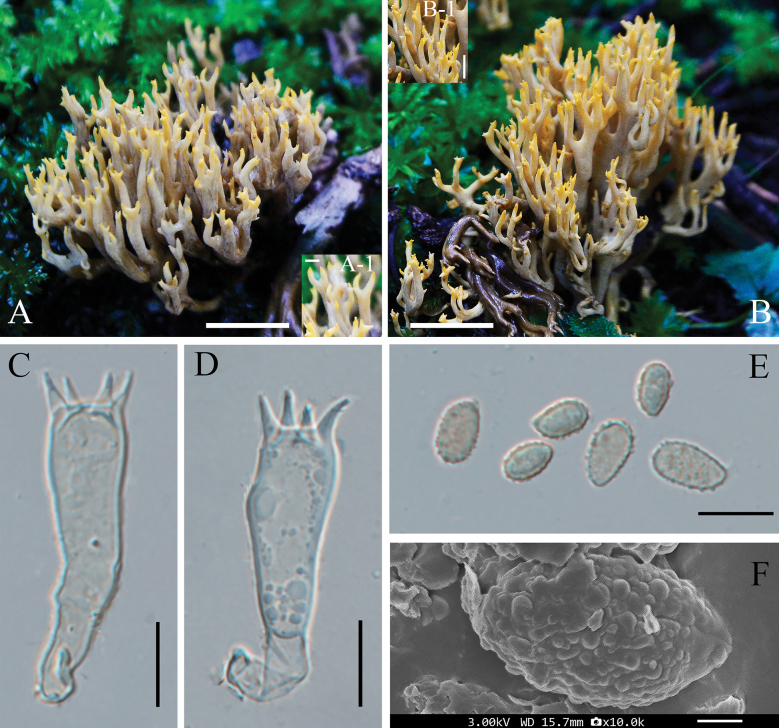 Figure 3
Figure 3| Taxon | Voucher | GenBank accession numbers | Geographical origin | References | |
|---|---|---|---|---|---|
|
|
| ||||
|
| OSC 134649 |
|
| USA | Unpublished |
|
| OSC 140661 |
|
| USA | Unpublished |
|
| DP387 |
|
| Argentina |
|
|
| DP392 |
|
| Argentina |
|
|
| MHHNU8909 |
|
| China |
|
|
| MHHNU6887 |
|
| China |
|
|
| TENN39621 | – |
| USA |
|
|
| AMB 18590 |
|
| Italy |
|
|
| AMB 18585 |
| – | Italy |
|
|
| AMB 18613 |
| – | Italy |
|
|
| DP031 |
|
| Argentina |
|
|
| DP386 |
|
| Argentina |
|
|
| AMB 18532 |
|
| Italy | Unpublished |
|
| MHHNU 11750 |
|
| China |
|
|
| MHHNU 12213 |
|
| China |
|
|
| BJM 344955 |
|
| China |
|
|
| BJTC L007 |
|
| China |
|
|
| MHHNU10702 |
|
| China |
|
|
| MHHNU10703 |
|
| China |
|
|
| AMB 18533 |
| – | Italy |
|
|
| AMB 18551 |
| – | Italy |
|
|
| AMB 18534 |
| – | Italy |
|
|
| AMB 18535 |
|
| Italy |
|
|
| AMB 18536 |
|
| Italy | Unpublished |
|
| MHHNU10376 |
|
| China |
|
|
| PRM:945441 |
| – | Czech |
|
|
| PRM:945440 |
|
| Czech |
|
|
| AMB 18562 |
| – | Italy | Unpublished |
|
| AMB 18531 |
|
| Italy | Unpublished |
|
| SJ99002 | – |
| Sweden |
|
|
| AMB 18641 |
|
| Italy |
|
|
| AMB 18605 |
| – | Italy |
|
|
| UBC |
| – | Canada | Unpublished |
|
| HAY-F-000746 |
| – | USA | Unpublished |
|
| TH9064 |
|
| Guyana | Unpublished |
|
| FH1 | – |
| USA | Unpublished |
|
| HKAS 45984 |
|
| China |
|
|
| HKAS 45992 |
|
| China |
|
|
| TENN36218 | – |
| USA |
|
|
| AMB n.18209 |
|
| Italy | Unpublished |
|
| AMB 18544 |
|
| Italy | Unpublished |
|
|
|
|
|
|
|
|
| BJTC C274 |
|
| China |
|
|
| BJTC ZH0015 |
| – | China |
|
|
| BJTC ZH1138 |
|
| China |
|
|
| FH109 | – |
| USA |
|
|
| BR079158-06 | – |
| USA |
|
|
| FH84 | – |
| USA |
|
|
| FH104 | – |
| USA |
|
|
| MHHNU9149 |
|
| China |
|
|
| MHHNU9164 |
|
| China |
|
|
| MHHNU10504 |
|
| China |
|
|
| 3281 | – |
| Mexico |
|
|
| 3533 | – |
| Mexico |
|
|
| 3266 | – |
| Mexico |
|
|
| 3563 | – |
| Mexico |
|
|
| TENN33826 | – |
| USA |
|
|
| AMB 18614 |
| – | Italy |
|
|
| AMB 18588 |
|
| Italy |
|
|
| AMB 18586 |
|
| Italy |
|
|
| AH:48382 |
| – | Spain |
|
|
| TENN39893 | – |
| USA | Unpublished |
|
| AGK 035 |
| – | USA | Unpublished |
|
| AMB 18589 |
|
| Italy |
|
|
| AMB 18542 |
|
| Italy | Unpublished |
|
| TENN31836 | – |
| USA |
|
|
| DP452 |
|
| Argentina |
|
|
| DP454 |
|
| Argentina |
|
|
| DP465 |
|
| Argentina |
|
|
| BBH 43575 |
|
| Thailand |
|
|
| BBH 43576 |
|
| Thailand |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| PRM:945445 |
| – | Czech |
|
|
| DP162 |
|
| Argentina |
|
|
| DP197 |
|
| Argentina |
|
|
| DP224 | – |
| Argentina |
|
|
| NY551 | – |
| USA | Unpublished |
|
| FH1853 | – |
| USA |
|
|
| OSC 97708 | – |
| USA |
|
|
| HKAS 127150 |
|
| China |
|
|
| HKAS 128154 |
|
| China |
|
|
| PDD:43383 | – |
| New Zealand | Unpublished |
|
| FH2 | – |
| USA | Unpublished |
|
| TENN69114 |
|
| USA |
|
|
| TENN69095 |
|
| USA |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Lichen and fungal ecology · Fungal Biology and Applications
Introduction
The genus Phaeoclavulina Brinkmann was coined by Brinkmann (1897) to accommodate coralloid fungi characterized by fleshy, often fragile basidiomata with varying degrees of branching and ochraceous to light brown basidiospores, with Phaeoclavulina macrospora Brinkmann from Germany as the type species. Brinkmann (1897) also suggested that Phaeoclavulina exhibits the closest morphological affinities to Clavaria testaceoflava Bres., Clavaria spinulosa Pers., and Clavaria abietina Pers. Based on Brinkmann’s original circumscription, Van Overeem (1923) adopted the concept of Phaeoclavulina, transferred Clavaria zippelii Lév. to this genus as Phaeoclavulina zippelii (Lév.) Overeem. Subsequently, Corner (1970) accommodated species of Phaeoclavulina within the subgenus Echinoramaria Corner of Ramaria Fr. ex Bonord., a taxonomic treatment that was subsequently followed by Petersen (1981) in his comprehensive revision of Ramaria. The taxonomic concept of the genus underwent a major revision when Giachini (2004), based on an integrative analysis of morphological characters and molecular phylogenetic data, recognized Phaeoclavulina as a distinct genus within Gomphus sensu lato. This revised classification was further supported by subsequent molecular phylogenetic analyses (Giachini et al. 2010), which provided robust evidence for the recognition of Phaeoclavulina as an independent lineage within the Gomphaceae.
Taxonomically, the genus Phaeoclavulina is placed in the family Gomphaceae, (Gomphales, Agaricomycetes, Basidiomycota) (He et al. 2019). Species of Phaeoclavulina typically produce branched to merismatoid basidiomata, with coloration ranging from white, yellowish brown, and orange to green, olivaceous, bluish green, violet, brown, reddish cinnamon, brick red, and gray and are micromorphologically characterized by echinulate or verrucose basidiospores, basidia bearing two to four sterigmata, and a monomitic hyphal system with frequent clamp connections and gloeoplerous hyphae (Krishnapriya 2023). In 2025, several new species were described, including Phaeoclavulina aurantilaeta P.T. Deng & P. Zhang (Deng et al. 2025), Phaeoclavulina aena Pelissero, Kuhar & Nouhra, Phaeoclavulina angularis Pelissero, Kuhar & Nouhra, Phaeoclavulina prasina Pelissero, Kuhar & Nouhra and Phaeoclavulina stelligera Pelissero, Kuhar & Nouhra (Pelissero et al. 2025), as well as Phaeoclavulina aurea Y. Gao, X. Tong & C.L. Hou and Phaeoclavulina fulva Y. Gao, X. Tong & C.L. Hou (Tong et al. 2025). These newly described species provide further evidence supporting the current circumscription of the genus.
At present, 68 species of Phaeoclavulina have been recorded globally (Species Fungorum 2025, https://www.speciesfungorum.org/Names/Names.asp), with 25 species reported from China (Teng 1963; Zhang et al. 2005; Li 2008; Liu et al. 2022; Deng et al. 2024, 2025; Zheng et al. 2024; Tong et al. 2025). Members of the genus Phaeoclavulina primarily inhabit soil or decaying wood substrates and are predominantly distributed in tropical and subtropical regions, although occurrences in temperate zones have also been recorded (Giachini 2004; Deng et al. 2024).
The Qinghai–Xizang Plateau (QXP) is the highest plateau in the world and is characterized by unique geographical features and highly diverse climatic conditions, which have given rise to exceptionally rich and highly endemic biodiversity (Cao et al. 2025; Wang et al. 2025b). In recent years, the QXP has become a hotspot for the discovery of new macrofungal taxa, highlighting its remarkable but still underexplored fungal diversity. In the year 2025 alone, multiple novel fungal species discovered on the QXP have been reported. Liu et al. (2025) described Entoloma griseopileum M.Q. He & X. Liu from Qinghai Province; Han et al. (2025) described four new Conocybe species from Xizang; Lei et al. (2025) introduced two new species of Pseudohydnum from Xizang; Wang et al. (2025a) reported 15 new species belonging to six orders, 11 families, and 12 genera of Agaricomycetes (Basidiomycota) from the southern border of Xizang; and another study by Wang et al. (2025b) revealed 64 novel taxa of macrofungi from the QXP and its surrounding areas. These findings collectively underscore the exceptional fungal richness and endemism of the QXP, emphasizing the urgent need for continued taxonomic and ecological investigations in this unique high-altitude ecosystem.
Although taxonomic research on the genus Phaeoclavulina has increased in recent years, its species diversity on the QXP remains poorly studied, and many unknown taxa are likely to exist in this region. Two previously undocumented species of Phaeoclavulina were collected during the field investigations in Qinghai Province in 2024 and are described as two new species based on morphology and phylogenetic analysis.
Materials and methods
Specimen collection
In September 2024, three specimens of Phaeoclavulina were collected by the authors from QXP, China. Field photographs were taken in situ prior to collection, capturing the shape, branching structure, and color characteristics of the basidiomata under natural light. Close-up images were also obtained to document diagnostic features such as branching patterns, surface textures, and color changes upon bruising. All photographs were taken using a digital camera with standardized settings to ensure consistent color representation. Fresh specimens were dried at 45–50 °C on the same day of collection, sealed in plastic bags containing silica gel desiccant. Voucher specimens are deposited in the Herbarium of Mycology of Jilin Agricultural University (HMJAU), China.
Morphological observation
Morphological identification was conducted based on both macroscopic and microscopic characteristics. Macroscopic features were primarily assessed using detailed field notes and photographs of fresh basidiomata taken at the time of collection. Color descriptions followed the standard color codes of Kornerup and Wanscher (1978). Sections of dried basidiomata were mounted in 5% KOH solution for the observation of internal tissue structures. Microscopic features, including basidiospores, basidia, and hyphae, were examined from dried material under a light microscope (Zeiss Axio Scope. A1, Germany).
To observe the surface ornamentation of basidiospores, dried spores were mounted on aluminum stubs with conductive carbon adhesive and coated with an approximately 10 nm layer of gold using a sputter coater (JEC-3000FC, JEOL Ltd., Japan), and then were observed with a field-emission scanning electron microscope (SEM; JSM-IT800, JEOL Ltd., Japan).
For each specimen, at least 30 mature basidiospores were randomly selected and measured. Spore dimensions were presented as (a)b–c(d), where b–c represents the 90% confidence interval (mean ± 1.64 × standard error), and a and d denote the extreme minimum and maximum values, respectively, and Qm represents the average Q value of all measured basidiospores ± the sample standard deviation (Liu et al. 2022).
DNA extraction, PCR amplification, and sequencing
Genomic DNA was extracted from 20 mg of dried fungal tissue, which was ground into a fine powder in liquid nitrogen, using the Hi-DNAsecure Plant Kit (Tiangen Biotech, Beijing, China) according to the manufacturer’s instructions. The internal transcribed spacer (ITS) and the large subunit (LSU) regions of the nuclear rDNA were amplified using the primer pairs ITS4/ITS5 (White et al. 1990; Gardes and Bruns 1993) and LR5/LR0R (Vilgalys and Hester 1990). PCR amplification was performed in a final reaction volume of 25 µL, containing 12.5 µL of Premix Taq [TaKaRa Taq (1.25 U/25 µL), 2 × dNTP mixture (0.4 mM each), and 2 × Taq buffer with 3 mM Mg^2+^] (TaKaRa, Tokyo, Japan), 9.5 µL of double-distilled water (ddH_2_O), 1 µL of each primer, and 1 µL of genomic DNA. The PCR program consisted of an initial denaturation at 94 °C for 4 min, followed by 32 cycles of denaturation at 94 °C for 40 s, annealing at 55 °C (for ITS) or 52 °C (for LSU) for 40 s, and extension at 72 °C for 1 min. A final extension was performed at 72 °C for 8 min, followed by storage at 4 °C (Deng et al. 2024). PCR products were visualized by electrophoresis on a 1.2% agarose gel, and high-quality amplicons were purified and sequenced by Sangon Biotech Co., Ltd. (Shanghai, China). Forward and reverse sequences were assembled into consensus sequences using DNAMAN version 9.0.1.116 (Lynnon Corporation), and all assembled sequences were manually checked to ensure accuracy. BLASTn searches were conducted in the NCBI nucleotide database to confirm sequence identity and assess similarity to previously published Phaeoclavulina species.
Alignment and phylogenetic analyses
The newly generated and validated sequences in this study were deposited in the GenBank database (Table 1). Based on previous studies (Deng et al. 2024, 2025; Zheng et al. 2024; Pelissero et al. 2025; Tong et al. 2025), available ITS and LSU sequences of 49 Phaeoclavulina species were retrieved from GenBank. Ramaria admiratia R.H. Petersen and Ramaria calvodistalis R.H. Petersen were selected as outgroup taxa based on their phylogenetic position (Zheng et al. 2024). The final ITS+LSU dataset, comprising 86 taxa (65 ITS and 71 LSU; Table 1), was used for multigene phylogenetic analyses. Multiple sequences were aligned using MUSCLE implemented in MEGA X (Kumar et al. 2018) and manually adjusted in BioEdit version 7.0.5.3 (Hall 1999).
Phylogenetic relationships were inferred using three complementary approaches: maximum parsimony (MP), maximum likelihood (ML), and Bayesian inference (BI). MP analyses were performed in PAUP* v4.0 (Swofford 2002) using a heuristic search with 100 random sequence additions. Tree statistics, including consistency index (CI), retention index (RI), and rescaled consistency index (RC), were calculated. ML analyses were performed using RAxML, implemented in raxmlGUI 2.0 (Edler et al. 2020), using the GTRGAMMA model with 1000 bootstrap replicates (Felsenstein 1985). BI analyses were conducted in MrBayes v3.2.7 (Ronquist et al. 2012), with the best-fitting nucleotide substitution model selected using MrModeltest v2.0 (Nylander 2004). Two parallel Markov chain Monte Carlo (MCMC) runs were executed for 1 million generations, sampling every 1000 generations and discarding the first 25% as burn-in. Convergence was assumed when the average standard deviation of split frequencies fell below 0.01. The topologies obtained from MP, ML, and BI analyses were congruent. The MP tree was selected for presentation, with branch lengths representing the number of inferred nucleotide substitutions. Bootstrap support values derived from MP, ML, and BI analyses were simultaneously mapped onto the corresponding nodes of the MP topology.
Results
Phylogenetic analyses
The concatenated ITS+LSU alignment matrix consisted of 1854 characters, of which 1102 positions (59.4%) were parsimony-informative, and 110 positions (5.9%) were parsimony-uninformative. Ramaria admiratia and R. calvodistalis were designated as outgroup taxa to root the phylogenetic tree.
Phylogenetic analyses using MP, ML, and BI methods consistently formed a monophyletic clade comprising all Phaeoclavulina taxa (Fig. 1). Phaeoclavulina fruticosa (HMJAU60999) was formed as sister to a subclade comprising Phaeoclavulina coniferarum Franchi & M. Marchetti and P. macrospora, with nodal support values (MP/ML/BI = 98%/ 95%/ 1.00). This phylogenetic placement indicates that P. fruticosa is not nested within any known species complexes and supports its genetic distinctiveness.
Phylogenetic tree of Phaeoclavulina species based on concatenated ITS and LSU sequences. Branches show bootstrap support values > 60% from maximum parsimony (MP) and maximum likelihood (ML) analyses, followed by posterior probabilities ≥ 0.70 from Bayesian inference (BI) analyses; “-” indicates support values below the respective threshold. Newly described species in this study are shown in red bold.
The two newly collected specimens of Phaeoclavulina qinghaiensis (HMJAU60997 and HMJAU60998) formed a distinct and well supported lineage (MP/ML/BI = 100%/100%/1.00). This lineage was formed as sister to Phaeoclavulina abietina (Pers.) Giachini, and the clade comprising P. qinghaiensis and P. abietina was in turn resolved as sister to a clade including Phaeoclavulina carovinacea Franchi & M. Marchetti, Phaeoclavulina nigricans E. Campo, Franchi & M. Marchetti, P. fulva, Phaeoclavulina yunnanensis W.H. Lu, D.G. Zheng, Karun. & Tibpromma. The relatively long branch length separating this lineage from its closest relatives indicates its phylogenetic distinctiveness.
Taxonomy
Phaeoclavulina
fruticosa
Taxon classificationFungiGomphalesGomphaceae
Y. Shan & S.Y. Liu sp. nov.
E16641DD-CAE2-5CE8-8276-1D9AB55C1E3B
860375
Etymology.
From the Latin “fruticosus” refers to the densely branched, shrub-like appearance of the basidiomata.
Diagnosis.
Basidiomata compact, densely branched, caespitose, turning blue when injured. Basidiospores dacryoid to ellipsoid with patched to subconical to verrucose ornamentations.
Type.
China • Qinghai Province, Xining City, Datong County, 37°04'16"N, 101°48'01"E, alt. 2870 m, 11 September 2024, Yong Shan, Shu-Yan Liu, Xue-Lian Wu, Sheng-Bang Zhang, Wan-Lin Zhao and De-Ning Zhang, HMJAU60999, holotype. GenBank: nuc rDNA ITS1-5.8S-ITS2 = PV821750; nuc rDNA 28S = PV822043.
Basidiomata.
Medium-sized, (20–)30–100(–105) × (6–)10–20(–24) mm, erect, compact, overall shrub-like to distinctly coralloid, typically caespitose, arising from soil on moss-covered coniferous forest floor. Branches with up to 4–5 levels of division, predominantly dichotomous; cylindrical to slightly compressed, ascending, mostly parallel or slightly divergent, closely spaced, and gradually tapering toward the apices; apices 1–2 mm long, usually blunt and commonly bifurcate. Stipe short to moderately developed, 10–50 mm high, distinctly widening toward the branching point. Texture fleshy, slightly viscid; tissues turning blue when injured. Fresh basidiomata grayish-yellow to olive-gray overall, with paler branch apices, yellow to yellowish-brown; with age, the overall coloration becomes progressively darker, turning olive-brown to brown. Surface dry to slightly moist, smooth to minutely pruinose. Context firm and fleshy; odor indistinct. Upon drying, basidiomata moderately shrink yet retain overall morphology and branching pattern; color darkens to olive-brown, with apices often remaining pale yellowish-brown; surface becomes hardened and brittle.
Basidiomata and microscopic features of Phaeoclavulina fruticosa sp. nov. A. Basidiomata; A-1. Enlarged view of the branch apex; B, C. Basidium; D. Basidiospores; E. Scanning micrograph of basidiospore. Scale bars: 2 cm (A); 2 mm (A-1); 10 μm (B–D); 1 μm (E).
Micromorphology.
Basidia 44.3–55.6 × 6.2–10.3 μm, slightly granular at maturity, hyaline to faintly yellowish, bearing four sterigmata. Hyphal system monomitic; hyphae hyaline, septate, with clamp connections, distinctly branched, (1.5–)2–6 μm wide, densely arranged, forming fasciculate or interwoven patterns.
Basidiospores (6.3–)7.4–8.0(–9.5) × (3.3–)4.1–4.5(–5.5) μm [Q = 1.45–2.16, Qm = 1.82 ± 0.2], dacryoid to ellipsoid, often slightly inequilateral, more or less rounded at one end and prolonged and sharp-pointed at the other; mature spores occasionally guttulate; surface roughened. Under SEM, basidiospores are distinctly ornamented, with surfaces sparsely covered by irregularly patched to subconical to verrucose ornamentations. The ornamentation elements are moderately sized, mostly discrete, and sparsely distributed across the spore surface, measuring approximately 0.15–0.35 µm in diameter. The ornamentations do not coalesce into ridges or form reticulate patterns.
Habit and distribution.
Gregarious on the soil around Picea crassifolia Kom. Known only from the type locality in Qinghai Province, China.
Notes.
Phaeoclavulina fruticosa forms a well-supported sister lineage to the P. coniferarum and P. macrospora clade. Despite this close phylogenetic relationship, P. fruticosa is readily distinguished from both species by a combination of macromorphological and micromorphological characters.
Phaeoclavulina fruticosa differs from P. coniferarum in having larger basidiomata (30–100 × 10–20 mm vs. 25–50 × 15–30 mm). With respect to fresh coloration and bruising reactions, basidiomata of P. fruticosa are grayish-yellow to olive-gray and turn blue when injured, while those of P. coniferarum are yellow to yellow-ochraceous and rapidly become green to cyan-green upon handling or exposure. Micromorphologically, P. fruticosa has larger basidia (44.3–55.6 × 6.2–10.3 μm) than P. coniferarum (35–45 × 5.5–7.5 μm). Furthermore, basidiospores of P. fruticosa are sparsely covered with irregularly patched to subconical to verrucose ornamentations, whereas P. coniferarum is characterized by densely spinose spores with spines reaching up to 0.5 μm in length.
Phaeoclavulina fruticosa differs from P. macrospora by its small basidiospores (7.4–8.0 × 4.1–4.5 μm vs. 14–20 × 4.5–8 μm). The basidiospore ornamentation also shows differences between the two species, spores of P. macrospora are prominently spinose, with spines reaching up to 2.5 μm in height, whereas spores of P. fruticosa are distinctly ornamented, with surfaces sparsely covered by irregularly patched to subconical to verrucose ornamentations (ca. 0.15–0.35 μm in diameter). The discoloration reactions upon injury are also different, with P. macrospora turning brown soon after bruising, whereas P. fruticosa turns blue.
Phaeoclavulina
qinghaiensis
Taxon classificationFungiGomphalesGomphaceae
Y. Shan & S.Y. Liu sp. nov.
992554EE-06D4-525C-B3A3-B7E2EE6719D4
860374
Etymology.
Qinghaiensis (Latin) refers to the type location, Qinghai Province, China.
Diagnosis.
Basidiomata yellow to orange-yellow, not changing color when bruised. Basidiospores are broadly ellipsoid to oblong and exhibit a dense covering of spherical to irregular warts.
Type.
China • Qinghai Province, Xining City, Datong County, 37°04'21"N, 101°49'04"E, alt. 2865 m, 11 September 2024, Yong Shan, Shu-Yan Liu, Xue-Lian Wu, Sheng-Bang Zhang, Wan-Lin Zhao and De-Ning Zhang, HMJAU60997, holotype. GenBank: nuc rDNA ITS1-5.8S-ITS2 = PV821748; nuc rDNA 28S = PV822041.
Basidiomata.
Arising from soil or moss-covered coniferous forest floor, typically gregarious or caespitose, erect, and distinctly coralloid in form, measuring (26–)30–60(–63) × (6–)10–20(–21) mm. Main branches thick, originating from a short stipe-like base and successively divided dichotomously or polychotomously, typically 4–5 times, with apices subacute, commonly forked or slightly recurved, measuring approximately 1.5–4 mm in length. Fresh basidiomata yellow to orange-yellow overall, paler at the base and slightly darker at the tips; color unchanging when bruised or injured. Surface dry, without gelatinous or tomentose covering; texture fleshy to firm. Stipe short and inconspicuous, central or slightly eccentric, often merging with the basal branches. In some specimens, basal stipe structures laterally connate, with distinct longitudinal grooves at the points of fusion, forming deeply embedded junctions. Basal parts frequently associated with rhizomorphic strands, sometimes bearing remnants of moss, pine needles, or cone scales. Upon drying, basidiomata dark brown to yellowish brown, with slight surface shrinkage but well-preserved branching structure; base pale yellowish white to light brown, with slightly fibrous texture.
Basidiomata and microscopic features of Phaeoclavulina qinghaiensis sp. nov. A, B. Basidiomata; A-1, B-1. Enlarged view of the branch apex; C, D. Basidium; E. Basidiospores; F. Scanning micrograph of basidiospore. Scale bars: 2 cm (A, B); 2 mm (A-1, B-1); 10 μm (C–E); 1 μm (F).
Micromorphology.
Basidia 39.1–50.9 × 5.9–9.1 μm, slightly granular upon maturation, hyaline to faintly yellowish, bearing four sterigmata. Hyphal system monomitic; hyphae septate, with clamp connections; contextual hyphae (1.8–)2–5 μm wide, with variable wall thickness, branched, densely arranged.
Basidiospores (6.5–)8.4–8.9(–11.0) × (3.5–)4.8–5.3(–6.7) μm [Q = 1.19–2.31, Qm = 1.74 ± 0.27], broadly ellipsoid to oblong, slightly inequilateral, more or less rounded at one end and narrower and slightly prolonged at the other; surface distinctly roughened. Some spores contain one to several guttules. Under SEM, spores exhibit a dense covering of warty (verrucose) ornamentation. These ornamentations are spherical to irregular in shape, moderately sized (0.1–0.5 µm in diameter), evenly distributed, and do not coalesce into ridges or reticulate structures.
Notes.
Phaeoclavulina qinghaiensis formed as sister to P. abietina in the phylogenetic analyses, but the two species can be distinguished by several morphological characters. Basidiomata of P. qinghaiensis remain unchanged in color when bruised or injured, whereas the flesh of P. abietina becomes slightly ochraceous when cut. Basidiospores of P. qinghaiensis are broadly ellipsoid to oblong and often slightly inequilateral, while those of P. abietina are elongate and pip-shaped. In addition, differences are observed in basidiospore ornamentation: basidiospores of Phaeoclavulina qinghaiensis are verrucose, with spherical to irregular ornamentation that are evenly distributed and of moderate size (0.1–0.5 μm in diameter), whereas basidiospores of P. abietina bear closely echinulate ornamentation composed of short, acute, hyaline spines 0.5 μm long, or 0.5–1 μm long in other collections.
Additional materials examined.
China • Qinghai Province, Xining City, Datong County, 37°04'21"N, 101°49'04"E, alt. 2860 m, 11 September 2024, Yong Shan, Shu-Yan Liu, Xue-Lian Wu, Sheng-Bang Zhang, Wan-Lin Zhao and De-Ning Zhang, HMJAU60998, paratype. GenBank: nuc rDNA ITS1-5.8S-ITS2 = PV821749; nuc rDNA 28S = PV822042.
Habit and distribution.
Gregarious on the soil around Picea crassifolia. Known only from the type locality in Qinghai Province, China.
Discussion
Phaeoclavulina was proposed by Brinkmann (1897) but remained controversial because many coralloid taxa with echinulate to verrucose basidiospores were traditionally placed in Ramaria, especially Ramaria subgenus Echinoramaria (Corner 1970). Subsequent integrative work has consistently recovered Phaeoclavulina as a distinct lineage, supporting its recognition as an independent genus (Giachini 2004; Giachini et al. 2010; Giachini and Castellano 2011). Morphologically, Phaeoclavulina species typically form coralloid (rarely subpileate) basidiomata with complex branching and variable pigmentation, and produce basidiospores that are usually echinulate to verrucose (Pelissero et al. 2025). The two new species in this study, P. fruticosa and P. qinghaiensis, conform to this concept and are placed in Phaeoclavulina by ITS–LSU phylogenetic analyses.
Species delimitation in Phaeoclavulina is supported by both molecular and morphological evidence. Two new species are formed as distinct, well-supported lineages in ITS and LSU analyses and can be separated from described species by stable morphological traits. A total of 68 species have been described in the genus Phaeoclavulina, bruise-induced discoloration and basidiospore ornamentation are essential characters for taxonomy. Of these, 16 species are reported to discolor after injury (Corner 1950; Giachini 2004; Franchi and Marchetti 2020; González-Ávila et al. 2020; Liu et al. 2022), and only eight species are characterized by predominantly verrucose spores (Giachini 2004; Zheng et al. 2024; Tong et al. 2025). Phaeoclavulina fruticosa is characterized by turning blue when bruised and irregularly patched to subconical to verrucose ornamentations of basidiospores. While Phaeoclavulina qinghaiensis is characterized by non-discoloring basidiomata and uniformly dense verrucae.
The QXP represents a habitat with distinctive environmental conditions and supports rich biodiversity, but Phaeoclavulina has been rarely studied there. Although Phaeoclavulina is widespread globally, records from the plateau are limited. To date, only 25 species have been reported from China. The discovery of P. fruticosa and P. qinghaiensis in high-elevation coniferous forests at approximately 2870 m in Qinghai Province further supports the view that the QXP harbors substantial but still poorly sampled diversity of Phaeoclavulina, and provides important insights into the presence of previously hidden species diversity within the genus. While the phylogenetic relationships of Phaeoclavulina at the genus level are well resolved, ecological differentiation and utilitarian traits among species remain incompletely known: P. flaccida is reported as toxic and inedible (Bau et al. 2024), whereas some ramarioid species currently placed in Phaeoclavulina are locally consumed in China (Li 2008; Dai et al. 2010), highlighting the need for more systematic studies of functional traits within the genus.
Supplementary Material
XML Treatment for Phaeoclavulina fruticosa
XML Treatment for Phaeoclavulina qinghaiensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bau T, Li HJ, Bao HY, Li Y (2024) A revised checklist of poisonous mushrooms in China. Journal of Fungal Research 22: 301–321. 10.13341/j.jfr.2024.0010 · doi ↗
- 2Cao GL, Li XQ, Zhang J, Peng HW, Li HL, Erst AS, Jabbour F, Ortiz Rdel C, Soltis PS, Soltis DE, Wang W (2025) Cenozoic geoclimatic changes drove the evolutionary dynamics of floristic endemism on the Qinghai–Tibet Plateau. Proceedings of the National Academy of Sciences of the United States of America 122: e 2426017122. 10.1073/pnas.2426017122 PMC 1223261040549922 · doi ↗ · pubmed ↗
- 3Corner EJH (1950) A monograph of Clavaria and allied genera. Oxford University Press, London, 1–740.
- 4Corner EJH (1970) Supplement to “A monograph of Clavaria and allied genera”. Beihefte zur Nova Hedwigia 33: 1–299.
- 5Deng PT, Liu WH, Ge ZW, Zhang P (2024) Three new ramarioid species of Phaeoclavulina (Gomphaceae, Gomphales) from China. Myco Keys 108: 1–14. 10.3897/mycokeys.108.128716 PMC 1135861539220354 · doi ↗ · pubmed ↗
- 6Deng PT, Huang ZH, He ZM, Zhang P (2025) Phaeoclavulina aurantilaeta (Gomphaceae, Gomphales), a new species of vivid fruiting body from China. Phytotaxa 696: 266–275. 10.11646/phytotaxa.696.4.1 · doi ↗
- 7Edler D, Klein J, Antonelli A, Silvestro D (2020) raxml GUI 2.0: A graphical interface and toolkit for phylogenetic analyses using R Ax ML. Methods in Ecology and Evolution 12: 373–377. 10.1111/2041-210X.13512 · doi ↗
- 8Felsenstein J (1985) Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39: 783–791. 10.2307/240867828561359 · doi ↗ · pubmed ↗