Morphological Distinctiveness and Phylogenetic Analysis of Strobilanthes malvifolia (Acanthaceae), a New Species From Dry‐Hot Valley in Yunnan, Southwest China
Huai‐Yu Chen, Feng Yang, Shao‐Yun Liu, Ting‐Ting Wang, Huan‐Chong Wang

TL;DR
A new plant species, Strobilanthes malvifolia, was discovered in a dry-hot valley in Yunnan, China, and is classified as endangered due to its limited range and small population.
Contribution
The discovery and description of a new Strobilanthes species with distinct morphology and phylogenetic placement in the Chamiponella group.
Findings
Strobilanthes malvifolia is a new species with cordate leaves and long petioles found in the Jinsha River Basin.
Phylogenetic analysis places it as sister to S. japonica within the Chamiponella group.
The species is classified as Endangered (EN) due to its restricted distribution and small population.
Abstract
The genus Strobilanthes (Acanthaceae) is known for its high morphological diversity and taxonomically challenging. During field investigations in Yunnan, southwest China, we discovered a remarkable new species, Strobilanthes malvifolia Huan C. Wang & Huai Yu Chen, which is described and illustrated here. This species exhibits distinct morphological characteristics, including cordate leaves with relatively long petioles (up to 8.7 cm long), sparsely and irregularly rounded serrations along the leaf margin. The complete chloroplast genome of this new species was also sequenced, assembled, and annotated. It is endemic to the Jinsha River Basin in western Yunnan, and was discovered in the dry‐hot valley characterized by extreme aridity and high temperatures. Phylogenetic analyses based on nrITS sequences support its placement as sister to S. japonica within the Chamiponella group.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
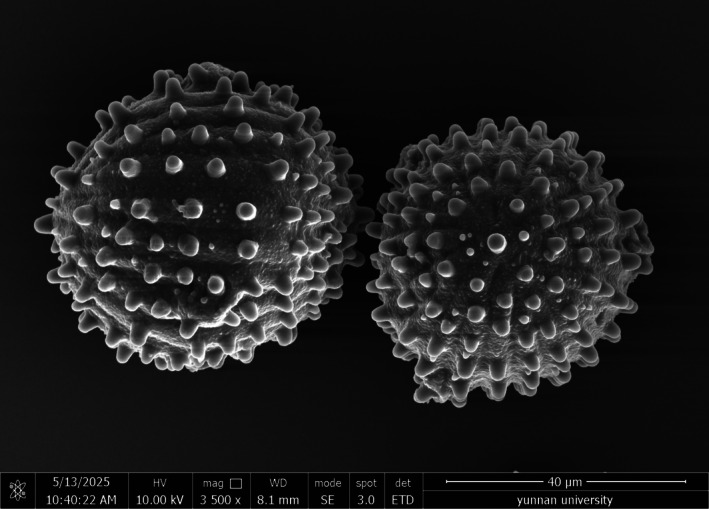 FIGURE 1
FIGURE 1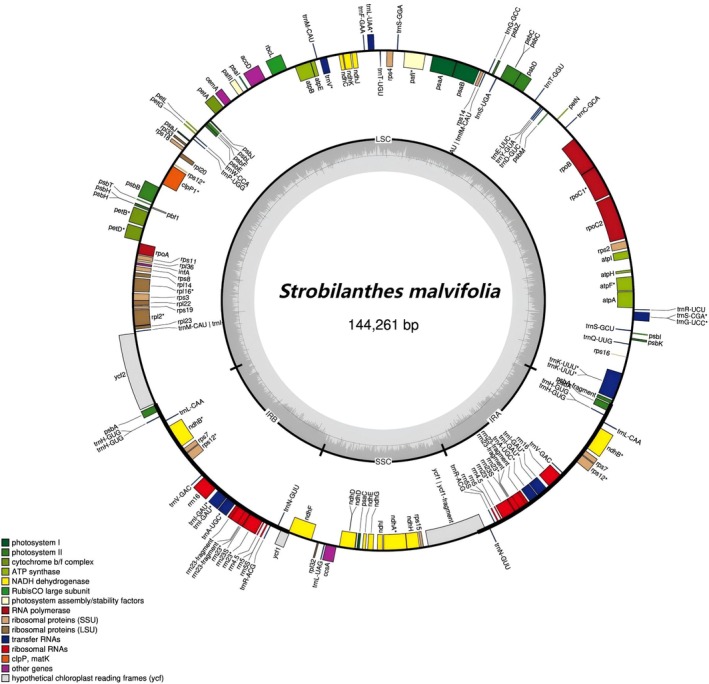 FIGURE 2
FIGURE 2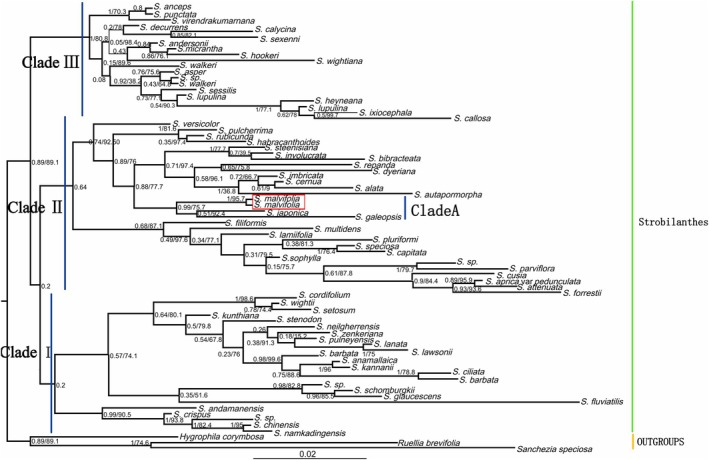 FIGURE 3
FIGURE 3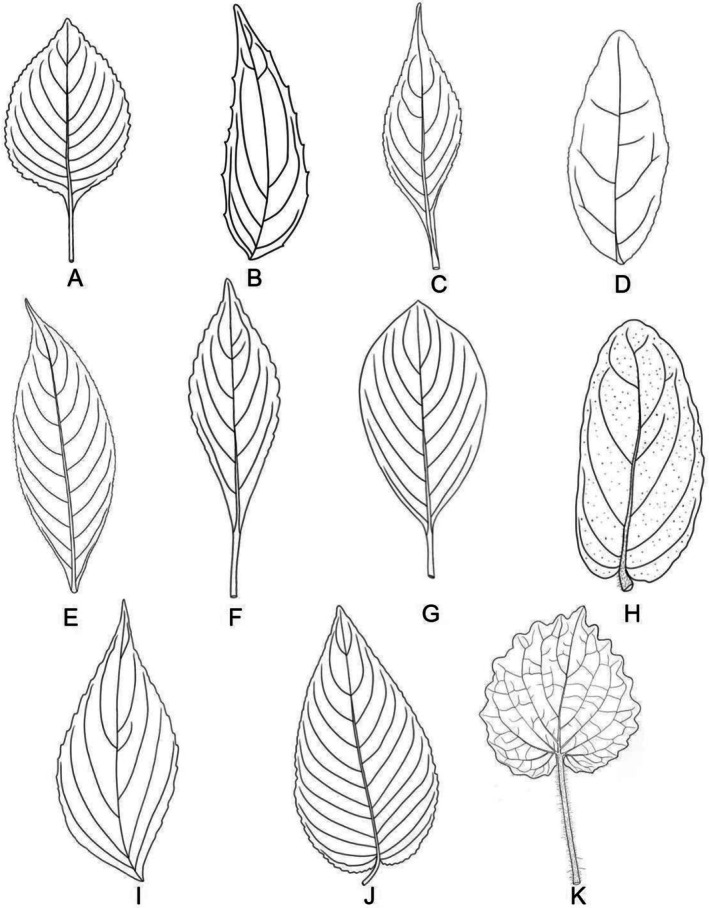 FIGURE 4
FIGURE 4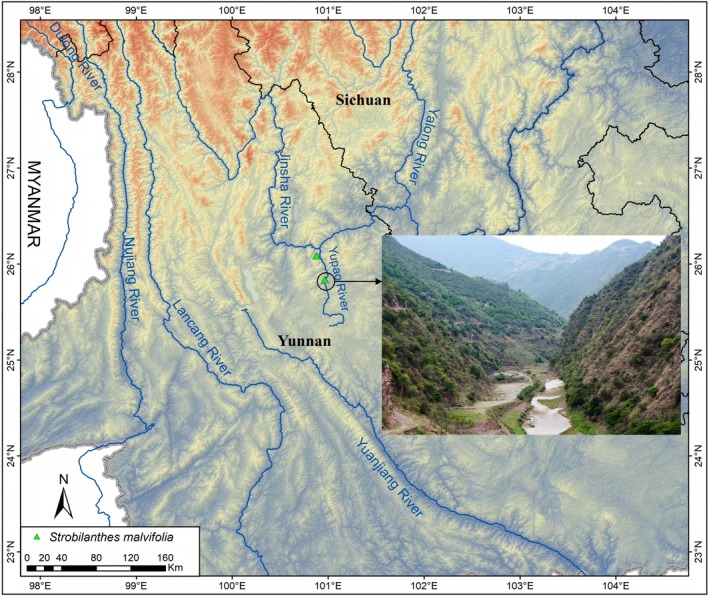 FIGURE 5
FIGURE 5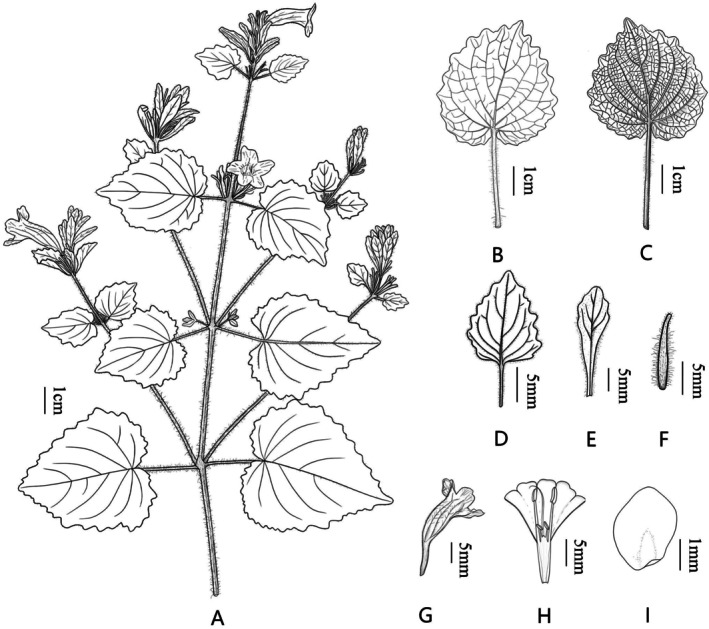 FIGURE 6
FIGURE 6 FIGURE 7
FIGURE 7 FIGURE 8
FIGURE 8| Characters | Species | |||
|---|---|---|---|---|
|
|
|
|
| |
| Habit | Herbaceous | Herbaceus | Herbaceous | Herbaceous |
| Stems | Erect, somewhat flexuose, crinite and pilose | Erect, nearly glabrous | Erect fulvo‐tomentellus | Erect, hirsute, setae |
| Petioles | 1–8.7 cm | 0.3–1.5 cm | 2.5–3 cm | 2.5–8.5 cm |
| Leaves dimension | 4–11 × 3.5–10 cm | 2–5 × 0.5–2 cm | 6–9 × 3–6 cm | 4.5–8 × 8–16 cm |
| Leaf shapes | Cordate | Elliptico‐lanceolata | Ovate | Ovate |
| Leaf bases | Cordate | Attenuata | Rotundata or brevissime contracta | Cordate |
| Leaf margins | Irregularly crenate | Entire or slightly repand | Dentato‐crenata | Serrata, serraturis incurvis |
| Leaf apices | Acuminate | Slightly obtuse and attenuate | Caudato‐acuminata | Caudato‐acuminata |
| Veins | Crinite, 4–7 pairs | Glabrous, 3–5 parirs | Setulosa, 7–8 pairs | Boary‐pubesceut, 4–8 pairs |
| Bracteses | Lanceolato, pubescent | Imbricate, spatulate‐obovate, ciliate | Lanceolato‐oblongae, setosae | Lanceolato‐linearis, glanduloso‐hirsutae |
| Braeteoles | Linear, glandular and pubescent | Linear, obtuse | Linear, calyce paulobreviores | Linear, glanduloso‐hirsutae |
| Calyces | Linear, glandular and Pubescent, segmenta subzequalia | Linear, scabrid‐puberulent, upper lobe longer | Lanceolato‐linearia, hirsuta and setulosa, segmenta subzequalia | Lineares, hirsutis‐glandulosis, laciniisupera longiore |
| Corollas | 1.7 cm, light purplish‐red, funnelform, short pubescence, glandular‐pilose pubescence | 1.5 cm, favescens, parum obliqua, campanulate‐cylindric | 2.6 cm, albus to violaceus, extus parce pilosulus | 2 cm, purpurea, glabra |
| Styles | Pilose | Hirtello‐puberus | Pilosula | Pubescent |
| Filamentums | Pubescent | Sparsely pilosulous | Pilosula | Sparsely pubescent |
| Seeds | 4, obliquely ovoid | 4, ovate | 4, ovato‐orbieularia | 4, linear‐oblong |
| References | / | Miquel ( | Stapf ( | Jacq. ex Nees ( |
- —the Second Tibetan Plateau Scientific Expedition and Research (STEP) program
- —the National Natural Science Foundation of China10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Diversity and Evolution · Genomics and Phylogenetic Studies · Plant and Fungal Species Descriptions
Introduction
1
Strobilanthes Blume (Ruellieae: Strobilanthinae) is one of the most diverse genera in the family Acanthaceae (Moylan et al. 2004; Tripp et al. 2022). It comprises approximately 450 species, mainly distributed in the tropical and subtropical regions of Asia, with some species extending to the Pacific Islands (Tripp et al. 2013; Deng 2019; Mabberley 2017). Strobilanthes is unique in having rugulae and trichomes that retain the style on the inner surface of the posterior corolla tube, and stamens monadelphous (by a sheath) at the base (Hu et al. 2011; Kladwong and Chantaranothai 2024). Some species in the genus are important medicinal plants, such as S. cusia (Nees) O. Kuntze, S. crispa (L.) Blume, S. ciliata Wall. ex Nees (e.g., George et al. 2017; Hu et al. 2011; Mabberley 2017; Ng et al. 2021; Zhu et al. 2022). In addition, some taxa also have significant value in ornamental use, dye production, and as beverages (Kumar et al. 1998; Zhang et al. 2021; Hua et al. 2022; Wood and Adhikari 2014).
Due to the highly variable morphological characteristics and rich species diversity, there has long been significant divergence in the circumscription of the genus since its establishment by Blume in 1826 (McDade et al. 2008). This has led to debate over whether the genus should be adopted with a broad or narrow circumscription. Anderson was the first to use ovule number, an anatomical feature, to separate morphologically similar species of this genus (Anderson 1867). Lindau (1895) applied pollen characteristics to the classification of the Acanthaceae and defined the genus Strobilanthes as having the “Rippenpollen” type, characterized by ellipsoidal pollen grains with ridges. Since then, pollen morphology has become an important basis for classification and has played a crucial role in distinguishing genera. Bremekamp (1944) divided Strobilanthes sensu Anderson (1867) into 54 smaller genera using combinations of macromorphological, anatomical, and pollen characters. However, many researchers have avoided Bremekamp's classification because the generic boundaries within subtribe Strobilanthinae remained ambiguous (Terao 1983; Wood 1994, 1995, 1998; Wood and Scotland 2003; Wood et al. 2003; Carine and Scotland 2002). Molecular studies (Moylan et al. 2004) have shown that most of these segregate genera in Bremekamp's classification are non‐monophyletic, and a single expanded genus, Strobilanthes sensu lato, is proposed at the level of the well‐supported monophyletic subtribe Strobilanthinae. Thereafter, the broad sense Strobilanthes has been generally accepted (Deng et al. 2006; Wood and Scotland 2009; Hu et al. 2011; Wood 2014; Deng 2019).
The genus Strobilanthes exhibits high diversity in China, which is one of the major distribution areas and centers of diversification for the genus. Hu and Cui (2006) adopted Bremekamp's classification for the systematics and taxonomy of taxa occurring in China, recording 114 species within the subtribe Strobilanthinae, belonging to 23 genera in Flora Reipublicae Popularis Sinicae. In the recent treatment of Flora of China, Hu et al. (2011) adopted the Strobilanthes sensu lato approach and recognized 128 species in China, including 57 endemics. Most of these species are distributed in the southwestern and southeastern China, with Yunnan Province having the highest species richness. As early as 1984, Wu adopted Bremekamp's classification in The catalog of seed plants in Yunnan Province, where he recorded 61 species in Yunnan, placed within 12 genera, and proposed new taxonomic combinations for three species (Wu 1984). Hu and Cui (2006) followed this classification framework in Flora Yunnanica, recording 72 species within 21 genera of the subtribe Strobilanthinae in Yunnan. In recent years, several new species have been reported (e.g., Chen et al. 2019, 2020; Deng, Wood, and Fu 2010; Deng, Wood, and Li 2010; Thanh et al. 2018; Thomas et al. 2018, 2019, 2020; Wood et al. 2019, 2022), leading to ongoing updates to the species checklist. According to the Catalogue of Life China 2025 Annual Checklist, a total of 136 species, 4 varieties, and 1 subspecies of Strobilanthes have been recorded from China to date (CBCAS 2025).
During investigations and research on plant diversity in Yunnan, southwestern China, we discovered a plant that differs markedly from known species of Strobilanthes in its leaf morphology. Through detailed observations of living individuals in the field, comparative analysis of herbarium specimens, morphological measurements in the laboratory, and thorough review of relevant literature (Wood 1994; Hu and Tsui 2002; Hu and Cui 2006; Karthikeyan et al. 2009; Hu et al. 2011; Adhikari 2018; Wood et al. 2022; Roy et al. 2023; Kladwong and Chantaranothai 2024), we have preliminarily determined that this plant represents a previously undescribed new species. In this study, we provide a comprehensive morphological description of the new species and discuss its systematic position within the genus based on molecular phylogenetic analyses and palynological evidence. Additionally, we present preliminary data on its geographic distribution, habitat preferences, and conservation status, offering new insights for future taxonomic and evolutionary studies of the genus.
Materials and Methods
2
Morphological Analyses
2.1
The morphology of the new species was studied based on observation of living plants and specimens housed at YUKU. Digital images of the genus available at online databases, including the JSTOR Global Plants (http://plants.jstor.org/), the Global Biodiversity Information Facility (GBIF, https://www.gbif.org/), the Indian Virtual Herbarium (https://ivh.bsi.gov.in/), and the Chinese Virtual Herbarium (CVH, https://www.cvh.ac.cn/), as well as collections housed at CDBI, IBSC, KUN, PE and YUKU, were extensively examined and compared with the new species. The herbarium abbreviations follow the Index Herbariorum (Thiers 2025, continuously updated). Pertinent taxonomic literature was extensively consulted (Wood 2014). Field trips were conducted in Dayao County, Yunnan to sample the new species. Images of the plants and flowers of the new species were obtained in the field using a D2X digital camera (Nikon, Tokyo, Japan), and all parts of the flowers were detached and photographed. Specimens with flowers were collected and preserved using both dry and wet methods (Jain and Rao 1977). For the morphological measurements and observations of this new species, the materials used in this study were primarily derived from three independent field gatherings (S. Y. Liu et al. DY23106; S. G. Li et al. DY23333; W. M. Zhu et al. 02557), comprising a total of 11 specimen sheets. The observations covered all morphological characters, including roots, stems, leaves, flowers, fruits, and seeds. The measured traits included plant height, leaf length, leaf width, petiole length, inflorescence length, pedicel length, bract length and width, bracteole length, calyx length, corolla length, and fruit length. All morphological observations and measurements were conducted using a stereomicroscope (Olympus SZX2, Tokyo, Japan), with a ruler (precision: 1 mm) or a metric vernier caliper (precision: 0.02 mm).
Palynological Studies
2.2
Mature pollen grains of the new species were sampled from the type materials and attached directly to carbon adhesive tape. They were coated with gold–palladium using a BAL‐TEC SCD 005 cool sputter coater (BAL‐TEC AG, Liechtenstein) at Yunnan University, Kunming, China. Observations were conducted using a Nova NanoSEM450 scanning electron microscope (SEM) (FEI Co., USA) at 10 kV at a magnification of 3500×. Descriptive terminology for the pollen grains follows Punt et al. (2007). Pollen dimensions, including polar axis (P) and equatorial axis (E), were measured using the updated software tool MATO (Liu et al. 2023).
Material Sampling, DNA Extraction, Sequencing, nrDNA and Plastome Assembly, and Annotation
2.3
Samples of the new species for molecular analysis were collected from Sanchahe Town, Dayao County, Yunnan Province, China. Samples and vouchers were deposited in the herbarium at Yunnan University (YUKU). The genomic DNA was extracted from silica gel‐dried leaves using the modified CTAB method (Doyle and Doyle 1987). The short‐insertion library (350 bp) was constructed and then sequenced to obtain 2 × 150 bp paired‐end data by using the Illumina NovaSeq 6000 platform (Illumina, San Diego, CA, USA) at Novogene (Beijing, China). The 3G of raw data for the new species was filtered through Trimmomatic v. 0.39 (Bolger et al. 2014) to obtain clean data, and then the clean data was quality‐controlled using FastQC v. 0.11.9 (Simon 2010). The nrDNA data and plastid genome were assembled using GetOrganelle v. 1.7.5 (Jin et al. 2020). The plastid genome was annotated using CPGAVAS2 (Shi et al. 2019) and Geseq (Tillich et al. 2017). The plastome maps were drawn using the Organellar Genome DRAW tool (Greiner et al. 2019).
Phylogenetic Analysis
2.4
To investigate the phylogenetic position of this species, two DNA sequences, including ITS and trnL‐trnF, were selected as phylogenetic markers in this study based on Fernandes et al. (2025). The ITS region was extracted from assembled nrDNA, and other ITS sequences are those used by Fernandes et al. (2025) and from NCBI database. There are a total of 74 taxa of Strobilanthes to be used, representing most major clades within the genus. In addition, Ruellia brevifolia (Pohl) C. Ezcurra, Hygrophila corymbosa (Blume) Lindau, and Sanchezia speciosa Leonard were selected as outgroups. A total of 70 taxa with 3 outgroups were included in the trnL‐trnF dataset. The trnL‐trnF plastid marker was extracted either from the annotated plastid genome or downloaded from GenBank. The voucher specimens and GenBank accession information of the molecular markers used in this study are listed in Table A1.
Due to the limited number of identical species between the two datasets, and most of the identical species are not from the same individual, a combined dataset analysis was not conducted. All sequences were aligned using MAFFT (Katoh and Standley 2013), and the phylogenetic tree was constructed using Phylosuite v.1.2.1 (Zhang et al. 2020; Xiang et al. 2023). Maximum Likelihood (ML) and Bayesian Inference (BI) methods were used to reconstruct phylogenetic trees. The ML analyses were conducted using IQ‐TREE (Nguyen et al. 2015; Minh et al. 2013; Guindon et al. 2010), ML bootstrap values (ML_BS_) were calculated applying 1000 bootstrap replicates with the substitution model chosen according to AIC. BI analysis and posterior probability (BI_PP_) calculation were also conducted in PhyloSuite v.1.2.1 using MrBayes (Ronquist et al. 2012). The best‐fit model for BI analysis was chosen according to BIC using ModelFinder (Kalyaanamoorthy et al. 2017).
Results
3
Morphological Characters
3.1
Strobilanthes malvifolia possesses distinctive leaf morphology within the genus. Its leaves are cordate with a length‐to‐width ratio close to 1:1, opposite and nearly equal in size, and with relatively long petioles, reaching up to 8.7 cm. The leaf margins are sparsely and irregularly crenate (Figure 4K). These characteristics clearly distinguish it from other species within the genus. A comparative analysis was conducted between the new species and the phylogenetically closest taxa ( S. japonica and S. galeopsis), as well as the morphologically more similar taxon ( S. attenuata ), as shown in Table 1. For a detailed morphological description, see Section 5.3 Taxonomic Treatment.
TABLE 1: Morphological comparison among Strobilanthes malvifolia, S. japonica, S. galeopsis, and S. attenuata .
Pollen Morphology
3.2
The pollen grains of Strobilanthes malvifolia are globose in shape (Figure 1), 57.65 (57.1–58.2) × 52.3 (51.2–53.4) μm, P/E = 0.79. They are circular in polar view, with radial symmetry, possessing three colpi alternating with pseudocolpi; the exine surface bears wart‐like verrucae of varying sizes, arranged in distinct or indistinct rows.
Pollen morphology of Strobilanthes malvifolia sp. nov. observed under SEM. The left one shows the equatorial view, and the right one shows the polar view.
Characteristics of the Complete Plastid Genome and nrDNA
3.3
The complete plastid genome sequence of Strobilanthes malvifolia is 144,261 bp in length. It features a circular and typical quadripartite structure, and contains a large single‐copy region (LSC, 52,574 bp), a small single‐copy region (SSC, 17,738 bp), and two reverse sequence repeat regions (IR, 36,975 bp each) (Figure 2). The percentage of GC in the whole genome is 38.3%, and the corresponding values in LSC, SSC, and IR regions are 34.7%, 30.2%, and 42.9%. We recovered a total of 129 functional genes, including 84 protein‐coding genes, 37 tRNA genes, and 8 rRNA genes. The trnL‐trnF region is located in the large single‐copy region of the chloroplast genome and includes the tRNA‐Leu (trnL) gene (partial sequence), the trnL‐trnF intergenic spacer (complete sequence), and the tRNA‐Phe (trnF) gene (partial sequence). The trnL‐trnF sequence of S. malvifolia is 876 bp in length, with a GC content of 36.9%. The entire nrDNA repeats, including ETS, 18S, ITS1, 5.8S, ITS2, and 26S regions, were 2199 bp in length. The GC content is 52.0%. The newly sequenced and newly assembled plastid genome, trnL‐trnF, and nrDNA sequences have been deposited in GenBank (accession numbers: PX570735, PX380933, and PX372082), and are listed in Table A1.
Chloroplast genome of Strobilanthes malvifolia sp. nov. Genes on the outside and inside of the circle are transcribed in the clockwise and counterclockwise directions, respectively. The dark and light gray bars in the inner circle denote G + C and A + T contents, respectively. The IRA and IRB (two inverted repeating regions); LSC (large single‐copy region); and SSC (small single‐copy region) are indicated outside of GC content.
Molecular Phylogenetic Studies
3.4
The alignment of ITS sequences resulted in a matrix of 473 total characters, 277 of which are constant; 82 of the variable characters are parsimony‐uninformative, and 114 characters are parsimony‐informative. The alignment of trnL‐trnF sequences resulted in a matrix of 891 total characters, 701 of which are constant; 264 of the variable characters are parsimony‐uninformative, and 155 characters are parsimony‐informative. The best substitution models of the ITS and trnL‐trnF matrix were TIM2 + F + I + G4 and HKY + F + G4, respectively.
The present study reconstructed the phylogenetic relationships of the genus Strobilanthes using two methods (ML and BI) based on ITS and trnL‐trnF datasets, which were similar to the results of an earlier study (Moylan et al. 2004; Fernandes et al. 2025). The phylogenetic trees generated from the ITS and trnL‐trnF datasets showed that the topology of the Bayesian inference (BI) tree was largely congruent with that of the maximum likelihood (ML) strict consensus tree. Therefore, only the Bayesian strict consensus trees are presented in Figure 3 and Figure A1, with Bayesian posterior probabilities and maximum likelihood bootstrap values indicated.
Phylogenetic tree of Strobilanthes Blume obtained by Mrbayes based on ITS sequences. Bootstrap percentages and Bayesian posterior probabilities are shown near the nodes (BIPP/BPML). The position of S. malvifolia sp. nov. is marked by a red box.
All sampled species sampled of Strobilanthes formed a well‐supported monophyletic clade in all analyses, and the majority of the subclades within the genus had strong support (Figure 3, BI_PP_ = 0.89/ML_BS_ = 89.1%; Figure A1, BI_PP_ = 1/ML_BS_ = 99.6%; all support values follow this order hereafter). The phylogenetic tree based on nuclear ITS sequences supported the division of Strobilanthes into three major clades (Clades I–III), with the relationship structured as ((Clade I, Clade II), Clade III) (Figure 3). Most species in Clade I are distributed in the tropical lowlands of the Indochina Peninsula—including Vietnam, Laos, Cambodia, Thailand, and Myanmar—with only a few exceptions such as S. fluviatilis (C. B. Clark ex W. W. Sm.) Moylan & Y. F. Deng and S. chinensis (Nees) J. R. I. Wood & Y. F. Deng occurring in southern China. Clade II contains numerous species distributed across a wide geographical range, including China, Japan, India, Malaysia, Thailand, and Java. As for Clade III, this clade comprises species with a relatively narrow distribution, confined to the Indian subcontinent (India and Sri Lanka), and it has not been recorded in China. Within Clade II, the newly discovered species S. malvifolia is recovered in a distinct sublineage (Clade A) and forms a strongly supported sister group with S. japonica (Thunb.) Miq. and S. galeopsis Stapf (Figure 3, BI_PP_ = 0.99/ML_BS_ = 75.7%). A plastid phylogeny constructed from trnL‐trnF sequences also supported the division of Strobilanthes into three major monophyletic clades (Figure A1, Clades I–III), in general agreement with the nuclear ITS phylogeny. However, discrepancies in the placement of certain taxa between the two analyses were observed, likely due to limited sampling and potential cytonuclear discordance. For example, S. cusia was placed in Clade II in the ITS phylogeny but grouped into Clade III in the trnL‐trnF phylogeny, suggesting possible cytonuclear incongruence or a complex evolutionary history. In the trnL‐trnF phylogeny, S. malvifolia is placed within Clade B, a relatively distinct lineage under Clade II, together with S. japonica , S. oligantha Miquel, and S. wakasana Wakas. & Naruh. in the same sublineage (Figure A1). Furthermore, although the phylogenetic tree constructed based on chloroplast DNA (cpDNA) has limited reference value for assessing the systematic position of this species due to the small number of available sequences, we have nevertheless included it in the appendix for reference (Figure A2).
Discussion
4
Morphological Distinctiveness of Strobilanthes malvifolia
4.1
Leaf shape in Strobilanthes is highly variable, including lanceolate, ovate, elliptic, orbicular, and rhomboid forms, and serves as an important diagnostic feature for distinguishing species and infraspecific taxa (Figure 4) (Deng et al. 2006; Hu et al. 2011). However, among previously documented Strobilanthes species from China and adjacent regions, truly cordate leaves have rarely been reported. Only a few species, such as S. attenuata Jacq. ex Nees and S. forrestii Diels, have cordate leaf bases. In the strict sense, S. malvifolia represents the first species in the genus with cordate leaves (Figure 4). Its leaf blade shows a nearly 1:1 length‐to‐width ratio, and the opposite leaves are nearly equal in size. The petiole is relatively long, reaching up to 8.7 cm. In addition, the leaf margin of S. malvifolia is highly distinctive bearing sparsely distributed, irregularly crenate teeth, a trait that is also rarely seen in other Strobilanthes species.
Leaf shape diversity of Strobilanthes (Drawn by Ting T. Wang). (A) S. kingdonii J. R. I. Wood. (B) S. mastersii T. Anderson. (C) S. atropurpurea Nees. (D) S. japonica (Thunb.) Miq. (E) S. auriculata Nees. (F) S. henryi Hemsl. (G) S. aprica (Hance) T. Anderson. (H) S. reptans (G. Forst.) Moylan ex Y. F. Deng & J. R. I. Wood. (I) S. oligantha Miquel. (J) S. tomentosa (Nees) J. R. I. Wood. (K) S. malvifolia sp. nov.
We conducted a comprehensive comparative analysis of the major vegetative and reproductive characters of S. malvifolia and its phylogenetically related species ( S. japonica and S. galeopsis) as well as the morphologically more similar species ( S. attenuata ). The results are presented in Table 1, which clearly highlights the diagnostic traits that distinguish S. malvifolia from these related taxa. In terms of reproductive structures, S. malvifolia exhibits a series of stable and pronounced diagnostic characters. In addition to its markedly distinctive leaf characteristics, S. malvifolia also exhibits clear differences in stem morphology. Its stems are densely covered with long hairs and bear crinite and pilose indumentum, which sharply contrasts with the nearly glabrous stems of S. japonica , the fulvo‐tomentose stems of S. galeopsis, and the hirsute and setose stems of S. attenuata . In terms of reproductive structures, S. malvifolia likewise possesses stable and pronounced diagnostic features. The bracts are lanceolate and densely pubescent, the calyx lobes bear glandular and pubescent hairs and are nearly equal in length, and the corolla is light purplish‐red.
Phylogenetic Analysis of Strobilanthes malvifolia
4.2
Currently, there is no widely accepted classification system for the genus Strobilanthes. Bremekamp (1944) mainly based on pollen and seed morphology, divided the subtribe Strobilanthinae into 54 genera and 27 informal groups. Pollen morphology holds significant value in the phylogenetic studies of the family Acanthaceae, particularly in comparative studies among different genera and species (Bremekamp 1944; Deng et al. 2006; Lindau 1895). Compared to other members of Acanthaceae, the genus Strobilanthes exhibits a greater diversity in pollen morphology, which has been extensively applied in species delimitation (Carine and Scotland 2002; Wang and Blackmore 2003; Hu et al. 2011). Scanning electron microscopy (SEM) observations show that the pollen grains of S. malvifolia are globose, with wart‐like protrusions of various sizes on the exine, arranged in clear or indistinct rows. A comparison of pollen traits reveals that the pollen of S. malvifolia closely resembles the type 10 identified by Hu et al. (2011), whose representative species is S. tetrasperma (Champ. ex Benth.) Druce. S. tetrasperma was classified in the Chamiponella group (Bremekamp 1944; Deng et al. 2006; Deng, Wood, and Fu 2010; Deng, Wood, and Li 2010). The group currently comprises 11 species: S. austrosinensis Y. F. Deng & J. R. I. Wood, S. japonica , S. labordei H. Lév., S. lihengiae Y. F. Deng & J. R. I. Wood, S. longiflora Benoist, S. oligantha , S. sarcorrhiza (C. Ling) C. Z. Cheng ex Y. F. Deng & N. H. Xia, S. tetrasperma, S. wakasana, S. wilsonii J. R. I. Wood & Y. F. Deng, and S. yunnanensis Diels (Li and Deng 2024). Through examining the pollen morphology of other species in the Chamiponella group (Deng et al. 2006; Gao et al. 2011), we found that S. malvifolia shares the same pollen type with members of this group. This similarity in pollen morphology is largely congruent with results of the phylogenetic analyses based on ITS and trnL‐trnF sequences.
Phylogenetic analysis based on ITS sequences revealed that Strobilanthes malvifolia and S. japonica form a clade (Figure 3, Clade A). Similarly, S. malvifolia and S. japonica continue to appear as sister taxa, and together form a clade (Clade B) with Strobilanthes oligantha Miq. and Strobilanthes wakasana Wakas. & Naruh. in the phylogenetic analysis based on the plastid trnL‐trnF sequence (Figure A1). Therefore, both phylogenetic evidence and palynological data support the placement of S. malvifolia within the Chamiponella group, indicating a close relationship with species such as S. japonica . Morphologically, S. malvifolia also matches key diagnostic characteristics of the Chamiponella group, including isomorphic petiolate leaves, terminal or axillary inflorescences, persistent bracts, nearly five‐lobed calyxes to the base, funnel‐shaped corollas, two strong fertile stamens, two ovules per ovary locule, and capsules containing four seeds. However, S. malvifolia is distinguished within the Chamiponella group by its unique leaf shape (cordate with sparsely and irregularly crenate margins) and habitat (dry‐hot river valleys), supporting its status as a distinct species within the group. Based on a comprehensive analysis of palynological, phylogenetic, and morphological data, S. malvifolia is most closely related to the Chamiponella group and is thus provisionally placed within this group of the genus Strobilanthes.
Habitat and Conservation Assessment
4.3
Currently, Strobilanthes malvifolia is known from only two locations. One specimen (W. M. Zhu et al. 02557, YUKU) was collected approximately 60 years ago from Pingchuan Town, Binchuan County, Yunnan Province. However, our subsequent field investigations failed to relocate the species at this site. The other known locality is the type locality, located in Sanchahe Town, Dayao County, Chuxiong Yi Autonomous Prefecture, at an elevation of approximately 1500 m. Both distribution sites lie in the Yupao River valley of north‐central Yunnan Province (Figure 5). The Yupao River is a major tributary of the middle reaches of the Jinsha River. This region is characterized by deeply incised valleys and low‐lying terrain, with a typical dry‐hot valley climate (Zhang et al. 2013; Yang et al. 2016). The mean annual temperature ranges from 18°C to 22°C, with extremely hot summers and mild winters. Extreme maximum temperatures can exceed 38°C (Yang et al. 2021). Annual precipitation is 600–800 mm, with most rainfall concentrated between June and September. The area receives more than 2400 h of sunshine annually. Notably, the dry season lasts for 7–8 months, and the annual evaporation is approximately 3–5 times the precipitation. This significant moisture deficit results in long‐term soil stress, posing a severe challenge to plant survival. S. malvifolia mainly grows at elevations of 1400–1800 m in broad‐leaved forests or mixed coniferous and broad‐leaved forests dominated by Pinus yunnanensis Franch. (Pinaceae). It commonly occurs in forest understories and along forest edges, where the soil consists of mountain yellow soil or yellow‐red soil, and the bedrock is primarily limestone.
Geographical distribution of Strobilanthes malvifolia sp. nov. (green triangle).
In southwest China, Strobilanthes exhibits remarkable species diversity (Hu and Cui 2006; Hu et al. 2011). In terms of habitat, most species grow in montane forests or shrublands and prefer warm, humid environments. Except for a few species, such as S. henryi Hemsl. and S. tomentosa (Nees) J. R. I. Wood, Strobilanthes species are rarely found in the dry‐hot valley habitats. A study by Nguyen et al. (2015) on Strobilanthes in Sri Lanka also showed that species diversity within the genus is significantly higher in moist, rainy areas than in arid regions. Other species in the Championella group are also widely distributed along shaded forest edges, valleys, or near streams in the southern to central subtropical regions of Asia, showing a clear preference for humid environments. Therefore, compared with other congeners, S. malvifolia exhibits a relatively unique adaptation to its habitat.
Based on current investigations, Strobilanthes malvifolia is a rare plant species with a very narrow distribution range, known only from its type locality and one nearby site. Fewer than 50 individuals have been recorded at its type locality, indicating a very small population size. The known distribution of this species is not included in any type of nature reserve or protected area, and the population is located near a rural road, making it highly susceptible to disturbances from road expansion and other human activities. According to the IUCN Red List Categories and Criteria (version 16) (IUCN 2024), this species meets the threshold for Criterion D (very small or restricted population), and we recommend it be preliminarily assessed as Endangered (EN).
Taxonomy Treatment
5
Strobilanthes malvifolia Huan C. Wang & Huai Yu Chen, sp. nov. (Figures 6, 7, 8)
5.1
Type. CHINA. Yunnan Province: Dayao County, Sanchahe Town, 25°50′ N, 100°58′ E, alt. 1500 m, 3 October 2023, (S.Y. Liu et al. DY23106) (Holotype YUKU!; isotypes YUKU!, PE!, KUN!).
Strobilanthes malvifolia sp. nov. (Drawn by Ting T. Wang). (A) habit. (B) Adaxial surface of leaf. (C) Abaxial surface of leaf. (D) Lower bract. (E) Floral bract. (F) Bracteole. (G) Flower (side view). (H) Corolla dissected to show both androecium and gynoecium. (I) Seed.
Strobilanthes malvifolia sp. nov. (A, B) Habit. (C) Roots. (D) A portion of stem, with densely villous. (E) Adaxial surface of leaf. (F) Abaxial surface of leaf. (G) Spike inflorescence on branches, forming a large panicle. (H) A portion of spike inflorescence to show bracteoles. (I) A calyx lobe. (J) Flower (side view). (K) Flower (front view). (L) Corolla dissected to show androecium. (M) Oblong anthers and curved filaments with pubescent. (N) Unmature fusiform capsule. (O) Opened capsule to show seeds.
The holotype of Strobilanthes malvifolia sp. nov. (YUKU‐02074897).
Diagnosis
5.2
Strobilanthes malvifolia can be distinguished from other species of Strobilanthes by the following characters: leaves cordate, with a length‐to‐width ratio nearly 1:1, base cordate, margin irregularly and sparsely crenate, with petioles up to 8.7 cm long, densely spreading pilose; bracts variable in shape, including foliaceous, elliptic, ovate, or oblong forms, and persistent.
Description
5.3
Herbs perennial, 0.3–0.8 m tall. Rhizome, slightly elongated, fibrous roots. Stem single, erect, somewhat flexuose, base subterete, usually densely lanate; remainder quadrangular, channeled, densely crinite and pilose. Leaves decussate, cordate, isophyllous, 4–11 cm long, 3.5–10 cm wide, papyraceous; base cordate; apex acuminate; margin irregularly crenate (9–16 pairs); veins 4–7 pairs, adaxially impressed, abaxially prominent; surface green, veins densely crinite, remainder pilose; abaxial surface pale green, veins densely crinite, remainder sparsely pilose; petiole 1–8.7 cm long, densely spreading crinite. Spikes terminal or axillary, paniculate on upper stems, 3–12 cm long, bearing 2–10 flowers, flowers decussate; peduncle quadrangular, 2–12 cm long, glandular crinite. Lower bracts foliaceous, 1.5–3 cm long, 1–3 cm wide, penniveined, margin crenate, both surfaces densely piloglandulose, persistent; floral bracts long‐elliptic, ovate, or oblong, 2–3 cm long, 0.3–0.8 cm wide, persistent; bractlets two, linear or linear‐lanceolate, 0.5–1 cm long, both surfaces densely piloglandulose, margin entire, densely ciliate. Calyx deeply five‐lobed, lobes linear to narrowly lanceolate, nearly equal, apex rounded, 0.7–2 cm long, green, densely glandular‐pilose. Corolla light purplish‐red, funnelform, limb 5‐lobed, lobes suborbicular, nearly equal, apex slightly emarginate. Corolla tube slender‐cylindrical, gibbous upward, densely glandular‐pilose externally, with two rows of short pubescence internally. Stamens 4, included, didynamous, longer pair straight, shorter pair incurved, filaments pubescent, basally connate, anthers elliptic, two‐locular, longitudinally dehiscent, spurless; pistil solitary, stigma slightly exserted, style pilose above. Capsule fusiform, 0.8–1.3 cm long. Seeds 4, obliquely ovoid, flattened, circum‐areolar, villosulous.
Etymology
5.4
The specific epithet “malvifolia” is derived from the generic name Malva L. (Malvaceae Juss.) by adding the suffix “‐folia” (denoting shape or form), specifically chosen to highlight the leaf morphological similarities of this new species to some members of the genus Malva, such as Malva cathayensis M. G. Gilbert, Y. Tang et Dorr particularly. This includes cordate leaf blades with shallow lobes or serrations, palmate venation patterns typical of mallows, and soft, fine pubescence resembling those found in many Malva species.
Vernacular Name
5.5
Simplified Chinese: 锦葵叶马蓝; Chinese pinyin: Jǐn kuí yè mǎ lán.
Phenology
5.6
Flowering period: September—November; Fruiting period: October—November.
Distribution
5.7
Strobilanthes malvifolia is found in western Yunnan Province, southwest China. Based on current surveys, this species is known only from its type locality, Sanchaha Town, Dayao County, Chuxiong Prefecture, and the neighboring Pingchuan Town, Binchuan County, Dali Prefecture, both situated in the dry‐hot valley of the Yupao River, a major tributary of the Jinsha River. It is a rare plant typically restricted to dry‐hot valleys and exhibits a unique adaptation to environments under severe water stress. Morphologically, S. malvifolia is a perennial deciduous herb, with most parts of the plant densely covered in hairs, features that likely reflect its adaptation to arid conditions. The species occurs at elevations of approximately 1400–1800 m. In the type location, the associated plants mainly include Pinus yunnanensis, Castanopsis delavayi Franch. (Fagaceae), Quercus variabilis Blume (Fagacea), Pistacia weinmanniifolia J. Poiss. ex Franch. (Anacardiaceae), Castanopsis orthacantha Franch. (Fagaceae), Dalbergia yunnanensis Franch. (Leguminosae), Bidens pilosa L. (Compositae), Gonostegia hirta (Blume) Miq. (Urticaceae), Ageratina adenophora (Spreng.) R. M. King et H. Rob. (Compositae), Oplismenus undulatifolius (Ard.) Roemer et Schuit. (Gramineae), Shuteria involucrata (Wall.) Wight et Arn. (Leguminosae), Diospyros dumetorum W. W. Sm. (Ebenaceae), Ageratum conyzoides L. (Compositae), Achyranthes bidentata Blume (Amaranthaceae).
Additional Specimens Examined
5.8
Strobilanthes malvifolia. China. Yunnan: Dayao County, Sanchahe town, elev. ca. 1500 m, 26 October 2023, S. G. Li et al. DY23333 (YUKU); Binchuan County, Pingchuan town, elev. ca. 1800 m, August 1965, W. M. Zhu et al. 02557 (YUKU).
Author Contributions
Huai‐Yu Chen: writing – original draft (equal). Feng Yang: writing – review and editing (equal). Shao‐Yun Liu: investigation (equal). Ting‐Ting Wang: visualization (equal). Huan‐Chong Wang: writing – review and editing (equal).
Funding
This work was supported by the National Natural Science Foundation of China (grant no: 31960040) and the Second Tibetan Plateau Scientific Expedition and Research (STEP) program (2019QZKK0502).
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Adhikari, B. 2018. “ Strobilanthes .” In Flora of Nepal Acanthaceae: Web Edition 2, edited by M. F. Watson , S. Akiyama , H. Ikeda , C. Pendry , K. R. Rajbhandari , and K. K. Shrestha , 36–49. Royal Botanic Garden Edinburgh.
- 2Anderson, T. 1867. “An Enumeration of the Indian Species of Acanthaceæ.” Journal of the Linnean Society of London, Botany 9: 455–526.
- 3Bolger, A. M. , M. Lohse , and B. Usadel . 2014. “Trimmomatic: A Flexible Trimmer for Illumina Sequence Data.” Bioinformatics 30, no. 15: 2114–2120.24695404 10.1093/bioinformatics/btu 170PMC 4103590 · doi ↗ · pubmed ↗
- 4Bremekamp, C. E. B. 1944. “Materials for a Monograph of the Strobilanthinae (Acanthaceae).” In Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen, Afdeling Natuurkunde, Reeks 2, vol. 41, 1–305. North‐Holland Publishing Company.
- 5Carine, M. A. , and R. W. Scotland . 2002. “Classification of Strobilanthinae (Acanthaceae): Trying to Classify the Unclassifiable?” Taxon 51: 259–279.
- 6Chen, F. L. , Y. F. Deng , Z. B. Xiong , and J. C. Ran . 2019. “Strobilanthes hongii, a New Species of Acanthaceae From Guizhou, China.” Phytotaxa 388, no. 1: 135.
- 7Chen, J. T. , X. H. Huang , Z. Y. Lu , et al. 2020. “ Strobilanthes sunhangii (Acanthaceae) a New Species From Tibet, China.” Phytokeys 166: 117–127.33239959 10.3897/phytokeys.166.58831 PMC 7679347 · doi ↗ · pubmed ↗
- 8Chinese Biodiversity Committee, Chinese Academy of Sciences (CBCAS) . 2025. Catalogue of Life China 2025 Annual Checklist. Science Press.