Basonuclin-2 promotes fracture repair through NuRD-dependent chromatin remodeling in periosteal stem cells
Zhong Zhang, Lingli Zhang, Bo Jiang, Shuqin Chen, Wenhui Xing, Peilong Wang, Lixiang Lou, Chunxiao Tang, Xuye Hu, Jinlong Suo, Bo O Zhou, Weiguo Zou, Lijun Wang

TL;DR
Basonuclin-2 (BNC2) helps repair bone fractures by regulating stem cell activity and chromatin changes in mice.
Contribution
BNC2 is identified as a novel regulator of periosteal stem cell expansion and fracture repair through NuRD-dependent chromatin remodeling.
Findings
BNC2 marks quiescent periosteal cells and is upregulated during fracture repair.
BNC2 depletion impairs stem cell proliferation and fracture healing in mice.
BNC2 promotes chromatin accessibility and histone acetylation for gene activation.
Abstract
Bone fracture healing remains a significant challenge in orthopedics, as injury-responsive skeletal stem cell (SSC) populations and the regulatory mechanisms governing SSC activation during nonunion fracture repair remain poorly delineated. This study identifies zinc finger transcription factor basonuclin-2 (BNC2) as a skeletal fracture repair control factor in periosteal stem cells. BNC2 marks quiescent periosteal cells during homeostasis and is significantly upregulated upon injury in mice, driving endochondral ossification post-fracture via clonal expansion. Moreover, knockout of Bnc2 in Prx1-cre+ cells (not Ocn-cre+ osteoblasts or LepR-creER+ BMSCs) resulted in impaired fracture healing, suppressing SSC proliferation. Mechanistically, ATAC-seq revealed that BNC2 deletion reduced chromatin accessibility at promoter regions of proliferation genes, hindering transcriptional activation.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
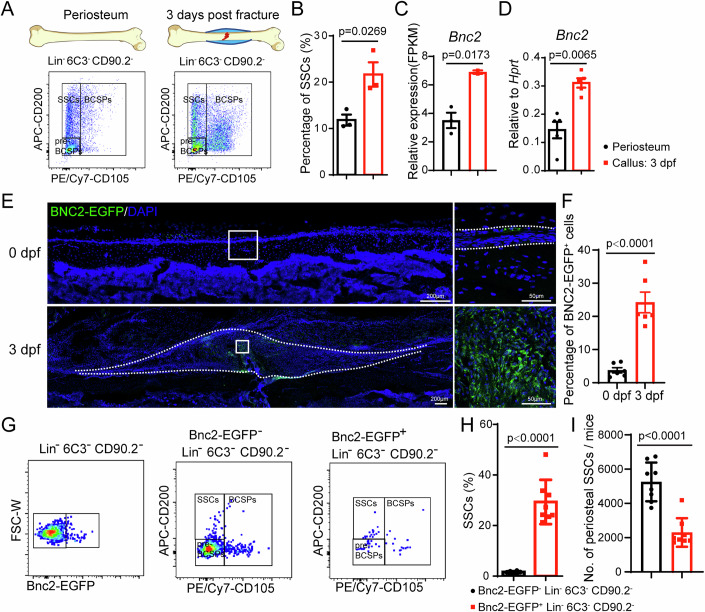 Figure 1
Figure 1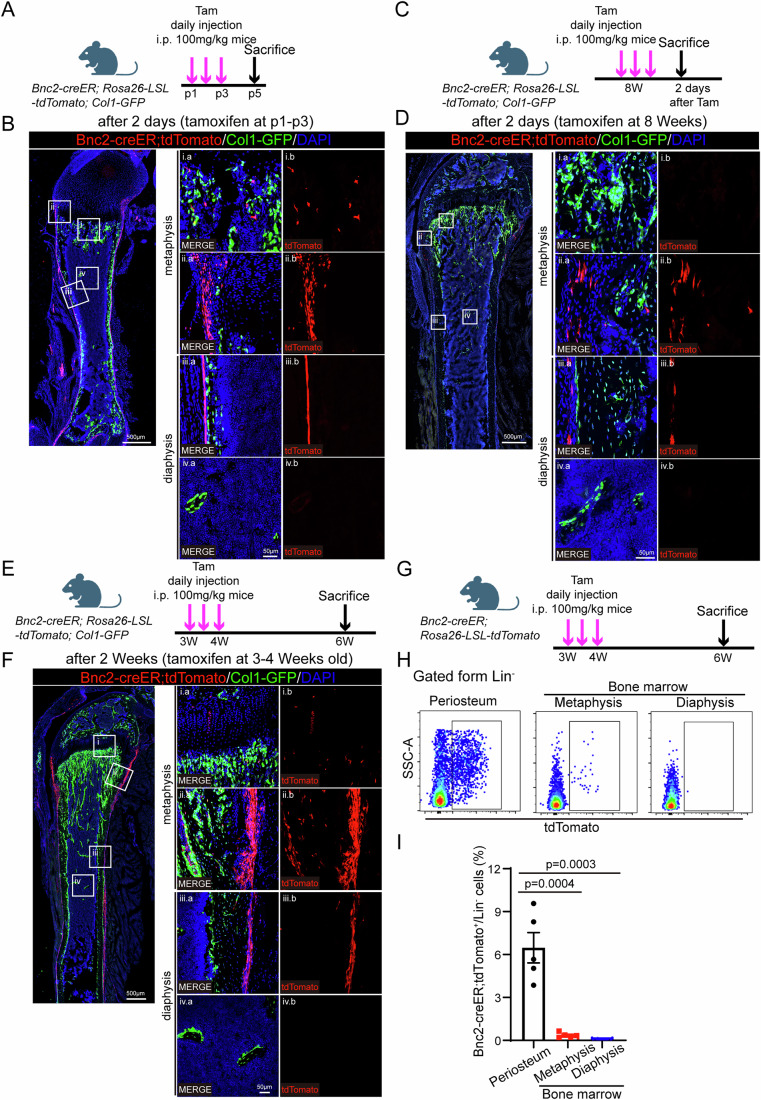 Figure 2
Figure 2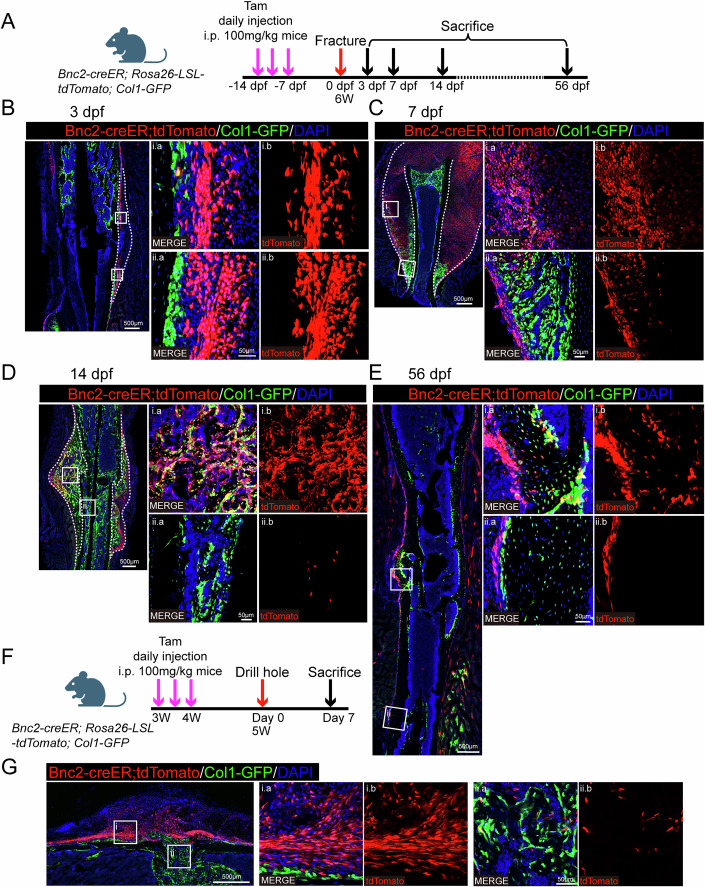 Figure 3
Figure 3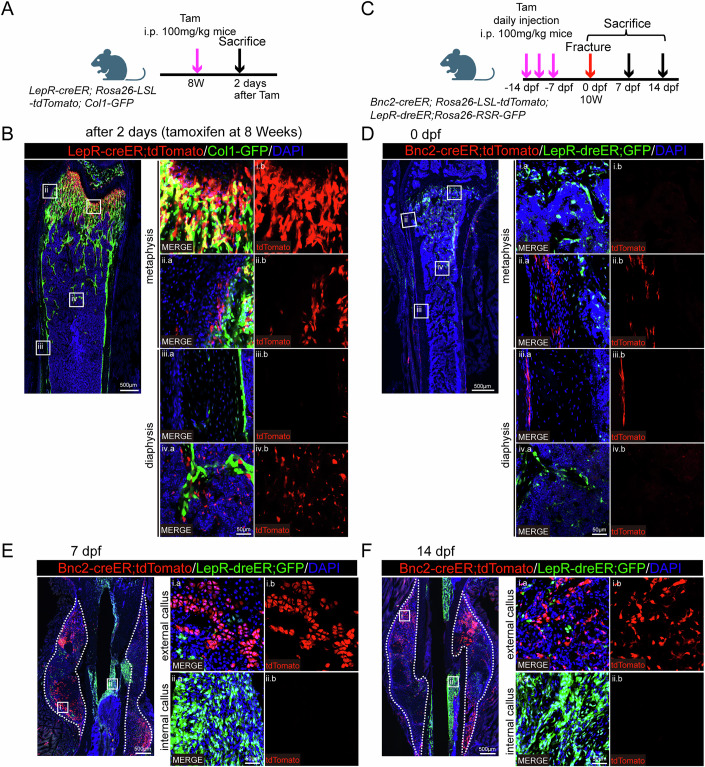 Figure 4
Figure 4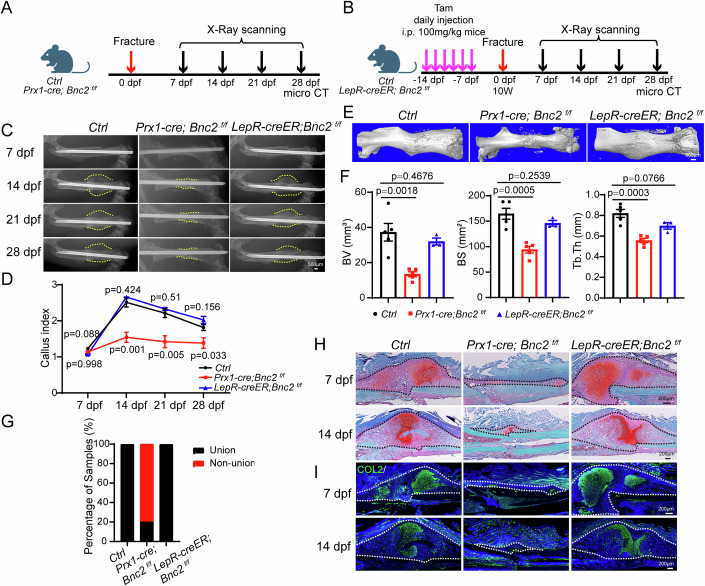 Figure 5
Figure 5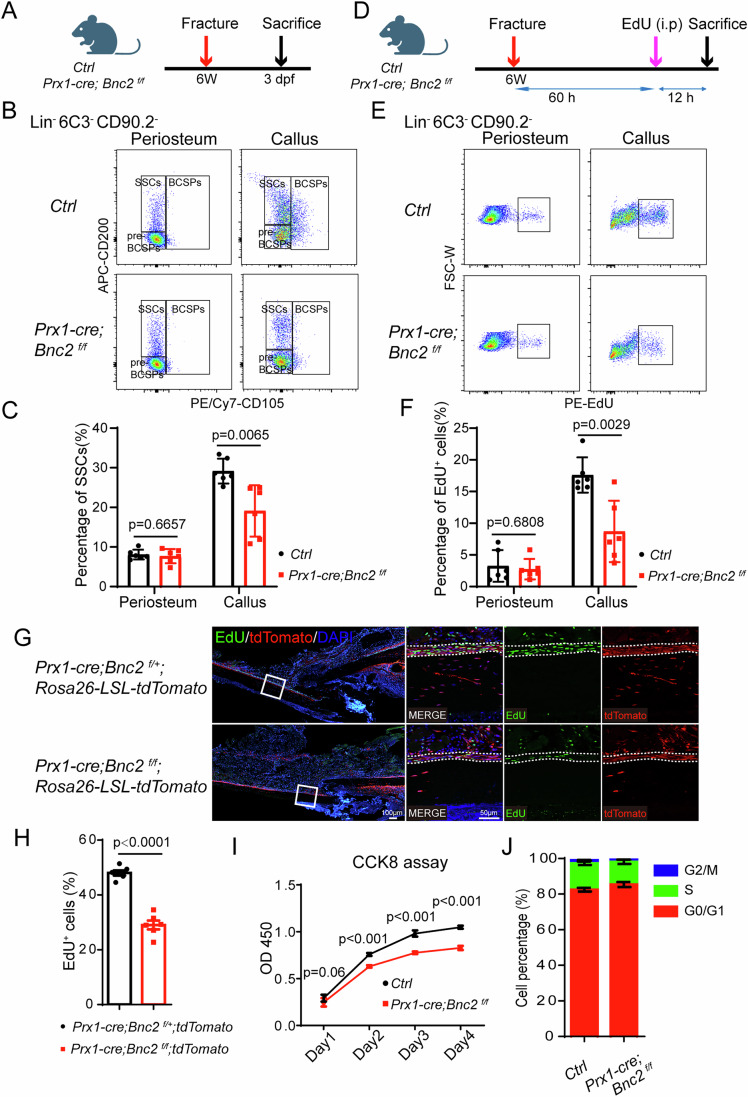 Figure 6
Figure 6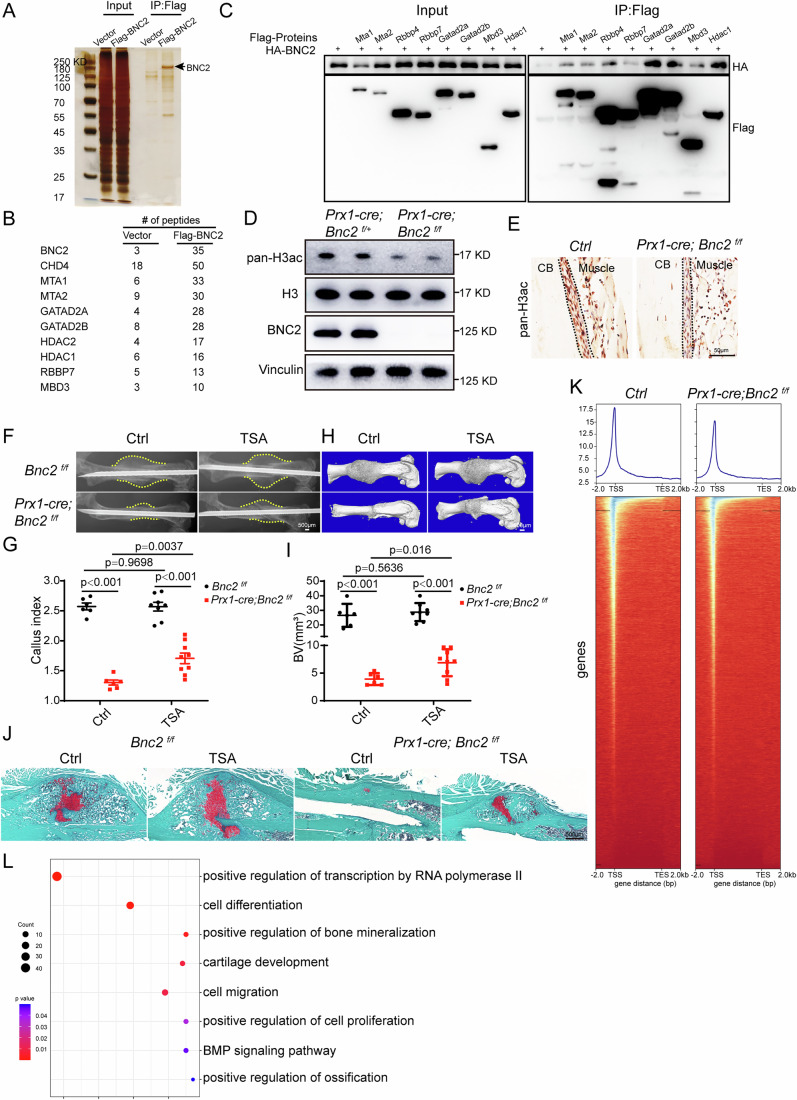 Figure 7
Figure 7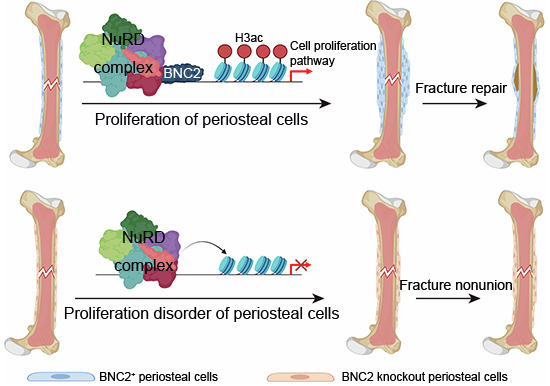 Figure 8
Figure 8 Figure 9
Figure 9- —http://dx.doi.org/10.13039/501100001809MOST | National Natural Science Foundation of China (NSFC)
- —Space Medical Experiment Project of China Manned Space Program
- —The Strategic Priority Research Program of the Chinese Academy of Science
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHistone Deacetylase Inhibitors Research · Epigenetics and DNA Methylation · Bone Metabolism and Diseases
Introduction
Bone fractures are the most common large organ traumatic injury in humans. Fracture healing is a postnatal regenerative process that recapitulates multiple events in embryonic skeletal development (Bais et al, 2009). Fracture repair usually restores the injured skeletal site to its pre-injury cellular composition, structure, and biomechanical function. However, there are still 5–10% of fractures that do not heal properly. The causes of nonunion are multifactorial, including the original trauma, the means of treatment, and the patient’s intrinsic factors, suggesting that both physical and biological factors can interfere with normal fracture healing (Wildemann et al, 2021). Stem cell function is the most important part of the patient’s intrinsic factors. Stem cell therapy is an important means to promote nonunion healing. Through specific cytokine treatment, endogenous stem cells could be recruited to the site of injury and further promote cell proliferation, migration, adhesion and differentiation (Tepper et al, 2005). SSCs in different parts of the bone have different molecular characteristics (Feng et al, 2022). For instance, Prx1 labels SSCs in both the bone marrow and periosteum (Liu et al, 2022), Ctsk specifically marks periosteal SSCs (Debnath et al, 2018), and LepR serves as a marker for SSCs localized within the bone marrow (Jeffery et al, 2022). Although multiple skeletal stem cell populations have been identified, it remains unclear whether there are skeletal stem cells that respond to injury repair without affecting homeostasis.
Epigenetic modification plays an important role in bone remodeling and bone repair. Knockout of the DNA methylase Dnmt3b in chondrocytes leads to delayed fracture repair and endochondral ossification (Wang et al, 2018). Deletion of H3K36 methylase NSD1 or H3K4 demethylase LSD1 in mesenchymal stem cells can lead to impaired fracture repair (Shao et al, 2021; Sun et al, 2020). During the differentiation process of osteoblasts, the acetylation levels of histones H3 and H4 in the promoter region of osteoblast-related marker genes increased significantly, and the knockdown of HDAC1 or treated with HDAC inhibitor trichostatin A (TSA) could enhance the differentiation of osteoblasts (Cho et al, 2014; Lee et al, 2006). In addition, TSA could promote human dental pulp stem cell proliferation and odontoblast differentiation (Jin et al, 2013). Inhibitors of BET family proteins can be used as drugs to target epigenetic modifications to promote bone repair (Chen et al, 2019). However, the regulatory mechanism of HDAC on fracture repair needs to be further explored.
Basonuclin-2 (BNC2) is a zinc finger protein that is predominantly expressed in germ cells, which is also widely expressed in skeletal systems, such as ear cartilage, nasal cartilage, intervertebral disc nucleus pulposus, and perichondrium of long bones. Bnc2 knockout mice have a body size reduction phenotype, and knockout of Bnc2 resulted in a reduced craniofacial mesenchymal cell proliferation (Vanhoutteghem et al, 2009). Marie Bobowski-Gerard et al, found that BNC2 expression was significantly promoted during the hepatic fibrosis process, and BNC2 could play a role in promoting fibrosis by activating TGFβ and Hippo/YAP1 pathways (Bobowski-Gerard et al, 2022). However, whether Bnc2 can affect bone repair and its regulatory mechanisms remains unknown.
Here, we found that the expression of Bnc2 was significantly upregulated in SSCs at the early stage after bone fracture by transcriptome sequencing. BNC2 could label periosteal cells, and these cells were fully involved in endochondral osteogenesis during fracture healing through lineage tracing with Bnc2-creER; Rosa26-LSL-tdTomato reporter mice, which also indicated the specific function of Bnc2 in periosteal SSCs. To further explore the regulatory effect of Bnc2 on fracture healing, we constructed Bnc2 conditional knockout mice lines in different cell populations. We found that Prx1-cre; Bnc2^f/f^ mice had significant impairments in fracture repair, while this phenotype was not present in Ocn-cre; Bnc2^f/f^ and LepR-creER; Bnc2^f/f^ mice, demonstrating that BNC2 played roles in periosteal SSCs. Mechanistically, BNC2 was indispensable for SSC proliferation upon injury. BNC2 regulated the level of H3ac of callus SSCs by interacting with the NuRD complex, thereby affecting chromatin accessibility to regulate gene transcription activation and cell proliferation.
Results
Bnc2 expression is upregulated in the periosteal SSCs after fracture
In order to explore the key transcription factors that regulate SSC activation in the early stage of fracture healing, we obtained SSCs from uninjured periosteum and 3 dpf callus (3 days post-fracture) by flow cytometry for bulk RNAseq (Fig. 1A). The flow cytometry results showed that the proportion of SSCs at the callus was significantly upregulated, indicating that the periosteal SSCs underwent significant proliferation (Fig. 1A,B). By performing KEGG enrichment analysis of genes upregulated in SSCs of periosteal callus, we found that SSCs in callus were significantly increased the expression of genes associated with cell cycle and DNA replication (Appendix Fig. S1A). There were five transcription factors in the significantly upregulated genes (Fig. 1C; Appendix Fig. S1B), among which E2f3 promoted cell proliferation by regulating the transition from G1 to S phase (Leone et al, 1998), and Bnc2 was reported to regulate normal meiosis in spermatogonial stem cells (Vanhoutteghem et al, 2014). Single-cell sequencing of Prx1-cre; Rosa26-LSL-tdTomato positive periosteal cells showed that Bnc2 was specifically expressed in skeletal precursor cells (Appendix Fig. S2A–C) (Wang, Ren et al, 2025; Data ref: Wang, Ren et al, 2025). The expression of Bnc2 in periosteal SSCs was significantly higher than that in bone marrow SSCs (Appendix Fig. S2D). QRT-PCR confirmed that Bnc2 expression was significantly upregulated in the 3 dpf callus, compared with uninjured periosteum (Fig. 1D). To further investigate the expression pattern of Bnc2 during fracture repair, we constructed a Bnc2-P2A-EGFP mouse model. Bnc2-EGFP positive cells existed in the periosteum and significantly increased upon injury (Fig. 1E,F). Flow cytometry analysis of 3 dpf callus cells from Bnc2-P2A-EGFP mice showed that the proportion of SSCs in Bnc2-EGFP positive cells was significantly higher than that in Bnc2-EGFP negative cells (Fig. 1G,H). However, the absolute number of Bnc2^+^ SSCs in the periosteum was less than that of Bnc2^-^ SSCs (Fig. 1I), since there are far more BNC2^-^ than BNC2^+^ in the Lin^-^6C3^-^Thy1^-^ gate. The above results demonstrate that Bnc2 is specifically expressed in periosteal SSCs in the early stage of fracture repair and is likely to regulate the proliferation of SSCs.Figure 1. Expression of Bnc2 is upregulated in the periosteal SSCs after fracture.(A, B) SSCs flow cytometry results (A) and quantitative statistical results (B) of callus cells on 3 dpf and undamaged periosteum. Data were presented as the means ± SEM. n = 3. Unpaired t-test. BCSP, bone cartilage stromal progenitor. (C) Expression of Bnc2 in RNAseq of 3 dpf callus and periosteal SSCs. Data were presented as the means ± SEM. n = 3, 2. Unpaired t-test. (D) Undamaged periosteum and 3 dpf callus SSCs were sorted, and qRT-PCR was used to detect the expression of Bnc2. Data were presented as the means ± SEM. n = 5. Unpaired t-test. (E, F) The expression of BNC2-EGFP on 0 dpf and 3 dpf, and the quantitative statistics of the proportion of BNC2-EGFP positive cells. The area within the dotted line is the callus. Data were presented as the means ± SEM. n = 6. Unpaired t-test. (G) Flow cytometry clustering of SSCs in BNC2-EGFP negative and positive periosteal cells. (H, I) Quantitative statistics of percentage and number of SSCs in BNC2-EGFP negative and positive periosteal cells. Data were presented as the means ± SEM. n = 8. Unpaired t-test. Source data are available online for this figure.
BNC2 could label periosteal cells
As Bnc2 was highly expressed in the SSCs of the periosteum (Appendix Fig. S2A–D), we sought to explore whether the specificity effect of Bnc2 on periosteal SSCs is due to its expression pattern. We constructed Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP (Col2.3-GFP) mice and treated the newborn mice with tamoxifen (Fig. 2A). Short-term labeling showed that Bnc2-creER specifically label the periosteal cells, including the periosteum near the growth plate and in the diaphyseal segment, with a few labeling in the metaphyseal cancellous bone region and no labeling at all in the bone marrow cavity of the diaphyseal segment (Fig. 2B). For adult Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mice, short-term labeling also showed that Bnc2-creER could specifically label periosteal cells (Fig. 2C,D). Long-term tracing showed similar results, except that the periosteum was more adequately labeled. However, periosteal Bnc2-creER positive cells did not transform into Col1-GFP positive osteoblasts in long-term tracing (Fig. 2E,F). By flow cytometry analysis of Bnc2-creER; Rosa26-LSL-tdTomato mice, we also confirmed that BNC2 mainly labeled periosteal cells, less labeled for metaphysis region, and hardly labeled for bone marrow in diaphysis region (Fig. 2G–I). These data suggest that BNC2 could label periosteal cells, and BNC2-positive periosteal cells are in a relatively resting state and do not contribute to osteogenesis in homeostasis.Figure 2Bnc2-creER specifically labels periosteum in homeostasis.(A, B) Lineage tracing of neonatal Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mice at 2 days after tamoxifen induction. (C, D) Lineage tracing of 8-week-old Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mice at 2 days after tamoxifen induction. (E and F) Lineage tracing of 4-week-old Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mice at 2 weeks after tamoxifen induction. (G) Schematic diagram of tamoxifen induction strategy in Bnc2-creER; Rosa26-LSL-tdTomato mice for flow cytometry. (H, I) The proportion and quantitative statistics of Bnc2-creER; Rosa26-LSL-tdTomato labels in periosteum, metaphyseal and diaphyseal bone marrow of long bone. Data were presented as the means ± SEM. n = 5. Unpaired t-test. Source data are available online for this figure.
BNC2-positive periosteal cells contribute to endochondral ossification during fracture healing
Since the expression of Bnc2 was significantly higher in SSCs from callus than in periosteal SSCs (Fig. 1C,D), we sought to explore whether BNC2-positive cells could be involved in fracture healing. We performed fracture surgery in Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mice (Fig. 3A). Bnc2-creER positive cells could fully label the proliferated periosteal cells on 3 dpf (Fig. 3B), and contribute to callus chondrocytes on 7 dpf (Fig. 3C). Besides, a small number of osteoblasts formed by intramembranous ossification at the distal part of callus were Bnc2-creER positive (Fig. 3C). Furthermore, Bnc2-creER positive cells could fully differentiate into osteoblasts formed by endochondral ossification in callus (Fig. 3D), but only made a very small contribution to the osteoblasts of callus in bone marrow on 14 dpf (Fig. 3D). After 56 days post-fracture, Bnc2-creER positive cells could form neo-bone at the healing site of fracture (Fig. 3E). At the distal end of the fracture, Bnc2-creER positive cells still only labeled the periosteal cells and did not contribute to bone formation at uninjured site (Fig. 3E). Then we tried to investigate whether Bnc2-creER positive periosteal cells participate in intramembrane osteogenesis during injury repair. We performed a bone drilling injury model with Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mice (Fig. 3F). BM SSCs, rather than periosteal SSCs, were reported to contribute to new bone formation in bone drilling injury model (Jeffery et al, 2022). Consistently, Bnc2-creER positive cells could still adequately label periosteal cells near the injured site, but had little contribution to osteoblasts formed at the injured site (Fig. 3G).Figure 3Bnc2-creER^+^ periosteal cells are involved in fracture healing.(A) Schematic diagram of Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mouse fracture model. (B) Slides on 3 dpf revealed Bnc2-creER; Rosa26-LSL-tdTomato positive cells specifically labeled thickened periosteum. The area within the dotted line is the callus. (C) Slides on 7 dpf revealed Bnc2-creER; Rosa26-LSL-tdTomato positive cells labeled a large number of chondrocytes, but significantly fewer osteoblasts which were derived from intramembrane osteogenesis. The area within the dotted line is the callus. (D) Slides on 14 dpf showed Bnc2-creER; Rosa26-LSL-tdTomato positive cells labeled a large number of osteoblasts in the external callus and almost no osteoblasts in the internal callus. The area within the dotted line is the callus. (E) Slides on 56 dpf revealed Bnc2-creER; Rosa26-LSL-tdTomato positive cells could form new periosteal cells, osteoblasts and osteocytes at the fracture site, while Bnc2-creER; Rosa26-LSL-tdTomato positive cells still only labeled periosteal cells at the distal fracture site. (F) Schematic diagram of a bone drilling model performed on Bnc2-creER; Rosa26-LSL-tdTomato; Col1-GFP mice. (G) The labeling of Bnc2-creER; Rosa26-LSL-tdTomato in periosteum and traumatic region on the 7th day after drilling. Source data are available online for this figure.
LepR has been shown to label BM SSCs and mainly contribute to the formation of callus within the bone marrow(Jeffery et al, 2022; Shu et al, 2021). Our studies in LepR-creER; Rosa26-LSL-tdTomato; Col1-GFP mice further confirmed that LepR-creER predominantly labels trabecular bone and BM SSCs (Fig. 4A,B). We observed that Bnc2-creER^+^ cells made very little contribution to the callus in the bone marrow (Fig. 3D). To further confirm the specificity of Bnc2 for periosteal cell labeling, LepR-dreER; Rosa26-RSR-GFP reporter mice was mated with Bnc2-creER; Rosa26-LSL-tdTomato mice to obtain dual-reporter mice. Fracture modeling was performed on Bnc2-creER; Rosa26-LSL-tdTomato; LepR-dreER; Rosa26-RSR-GFP mice (Fig. 4C). Utilizing dual-reporter mouse models, we demonstrated that under uninjured conditions, Bnc2-creER exclusively labels periosteal cells, whereas LepR-dreER labels trabecular bone and BM SSCs (Fig. 4D). During fracture healing, the specificity of Bnc2-creER; Rosa26-LSL-tdTomato positive cells’ contribution to the external callus was demonstrated, and LepR-dreER; Rosa26-RSR-GFP positive cells mainly formed the internal callus in bone marrow, and a small amount of LepR-dreER; Rosa26-RSR-GFP positive cells was found in the external callus. However, there was no colocalization of Bnc2-creER; Rosa26-LSL-tdTomato and LepR-dreER; Rosa26-RSR-GFP positive cells in the external callus (Fig. 4E,F).Figure 4Bnc2-creER^+^ periosteal cells are specifically involved in the formation of external callus.(A, B) Lineage tracing of 8-week-old LepR-creER; Rosa26-LSL-tdTomato; Col1-GFP mice at 2 days after tamoxifen induction. (C) Schematic diagram of Bnc2-creER; Rosa26-LSL-tdTomato; LepR-dreER; Rosa26-RSR-GFP mouse fracture model. (D) Lineage tracing of 8-week-old Bnc2-creER; Rosa26-LSL-tdTomato; LepR-dreER; Rosa26-RSR-GFP mice at 7 days after tamoxifen induction (0 dpf). (E, F) Slides on 7 dpf (E) and 14 dpf (F) showed Bnc2-creER; Rosa26-LSL-tdTomato positive cells participated in the formation of external callus, but not in the formation of internal callus. And LepR-dreER; Rosa26-RSR-GFP positive cells were mainly involved in the formation of internal callus but contributed little to the formation of external callus. The area within the dotted line is the callus. Source data are available online for this figure.
Our findings demonstrate that BNC2 could label a subpopulation of periosteal SSCs that exhibit robust engagement in endochondral ossification during fracture repair. These BNC2^+^ cells orchestrate the regeneration of functional periosteal tissue and cortical bone architecture critical for structural restoration, while showing negligible contribution to bone marrow-derived internal callus formation during the healing process.
Loss of Bnc2 in periosteal SSCs shows significant impairment in fracture healing
To explore whether Bnc2 is necessary for periosteal SSCs function in bone repair, we delete Bnc2 in SSCs/osteoblasts/osteocytes using Prx1-cre; Bnc2^f/f^ mice, and in BM SSCs using LepR-creER; Bnc2^f/f^ mice (Fig. 5A,B). X-Ray analysis showed that the callus volume was significantly smaller in Prx1-cre; Bnc2^f/f^ mice but not in LepR-creER; Bnc2^f/f^ mice (Fig. 5C,D). The fracture nonunion and reduced newly formed bone volume was observed in Prx1-cre; Bnc2^f/f^ mice but not in LepR-creER; Bnc2^f/f^ mice, as assessed by micro-CT scanning and quantification (Fig. 5E–G). Consistently, Safranine O/Fast green (SOFG) and immunostaining of COL2 demonstrated that loss of Bnc2 in Prx1-cre^+^ but not in LepR-creER^+^ cells impaired endochondral osteogenesis during fracture healing (Fig. 5H,I). In order to explore whether Bnc2 deficiency in Prx1-cre^+^ cells affects intramembrane osteogenesis during repair, Prx1-cre; Bnc2^f/f^ mice were subjected to a bone drilling injury model, as this injury was repaired by intramembranous ossification (Colnot, 2009). SOFG staining and OPN (Osteopontin) immunofluorescence staining showed that Prx1-cre; Bnc2^f/f^ mice could normally form trabecular bone at the site of injury 14 days after surgery (Appendix Fig. S3A,B). Micro-CT scanning results also showed that there was no significant difference between Prx1-cre; Bnc2^f/f^ and control mice during drilling damage repair (Appendix Fig. S3C,D).Figure 5. Loss of Bnc2 in periosteal SSCs shows significant impairment in fracture healing.(A) Schematic diagram of Prx1-cre; Bnc2^f/f^ mice fracture model. (B) Schematic diagram of LepR-creER; Bnc2^f/f^ mice fracture model. (C, D) X-ray results (C) and quantitative statistics of callus index (D) in Prx1-cre; Bnc2^f/f^ and LepR-creER; Bnc2^f/f^ mice at different stages after fracture. Data were presented as the means ± SEM. n = 5, 5, 3. Unpaired t-test. The dotted line represents the edge of the callus. (E, F) Micro-CT scanning results (E) and quantitative statistics of new bone formation at the fracture site (F) of Prx1-cre; Bnc2^f/f^ and LepR-creER; Bnc2^f/f^ mice on 28 dpf. Data were presented as the means ± SEM. n = 5, 5, 3. Unpaired t-test. (G) Quantitative statistics of percentage of union and nonunion fracture samples on 28 dpf in Prx1-cre; Bnc2^f/f^ and LepR-creER; Bnc2^f/f^ mice. n = 5, 5, 3. (H) SOFG staining of Prx1-cre; Bnc2^f/f^ and LepR-creER; Bnc2^f/f^ mice on 7 dpf and 14 dpf. The area within the dotted line is the callus. (I) COL2 immunofluorescence staining of Prx1-cre; Bnc2^f/f^ and LepR-creER; Bnc2^f/f^ mice on 7 dpf and 14 dpf. The area within the dotted line is the callus. Source data are available online for this figure.
To dissect the cell type-specific functions of BNC2 in fracture healing, we employed complementary genetic strategies: To eliminate the influence of the extensive expression of Prx1-Cre during development (SSCs/osteoblasts/osteocytes), we used the Prx1-CreER mouse model to knock out Bnc2 at adulthood, thereby achieving the knockout of Bnc2 in periosteal cells in the cortical bone region. X-ray analysis showed a decreased callus volume of Prx1-creER; Bnc2^f/f^ mice during the fracture healing process (Appendix Fig. S4A–C). The micro-CT results showed that there was an obvious nonunion of bone fracture in Prx1-creER; Bnc2^f/f^ mice (Appendix Fig. S4D,E). Besides, we used the Ocn-cre; Bnc2^f/f^ mice to selectively ablate Bnc2 in mature osteo-lineage cells. Quantitative analysis revealed comparable callus volumes between Ocn-cre; Bnc2^f/f^ mice and controls during fracture healing (Appendix Fig. S5A,B), with micro-CT at 4 weeks post-fracture confirming complete bony union in both groups (Appendix Fig. S5C,D). Histological assessment via SOFG staining and COL2 immunostaining demonstrated normal endochondral ossification dynamics in Ocn-cre; Bnc2^f/f^ mice callus (Appendix Fig. S5E,F). Moreover, tamoxifen-inducible Ocn-creER; Rosa26-LSL-tdTomato lineage tracing exhibited strict exclusion from periosteal stem cell pools and chondrogenic lineages throughout healing phases (Appendix Fig. S6A–C).
These data establish a functional dichotomy: Prx1-cre; Bnc2^f/f^ and Prx1-creER; Bnc2^f/f^ mice display arrested healing, whereas both LepR-creER; Bnc2^f/f^ and Ocn-cre; Bnc2^f/f^ mice models maintain normal repair kinetics. This genetic epistasis analysis unequivocally identifies periosteal SSC-derived BNC2 as an indispensable regulator of fracture healing.
Bnc2 deficiency inhibits SSC activation in the early stage of fracture healing
The periosteal thickening response in Prx1-cre; Bnc2^f/f^ mice was significantly impaired on 3 dpf (Appendix Fig. S7A), thus we sought to explore whether the impairment of fracture healing in Prx1-cre; Bnc2^f/f^ mice begin with early obstruction of periosteal cell proliferation. For this, the proportion of SSCs in Prx1-cre; Bnc2^f/f^ mice was analyzed by flow cytometry (Fig. 6A). The proportion of SSCs significantly reduced in the 3 dpf callus but not in the uninjured periosteum of Prx1-cre; Bnc2^f/f^ mice, compared with littermate control (Fig. 6B,C). To further confirm the effect of Bnc2 on SSC activation, we labeled Prx1-cre; Bnc2^f/f^ mice with EdU injection (Fig. 6D). The proportion of EdU^+^ precursor cells significantly reduced in the 3 dpf callus but not in the uninjured periosteum of Prx1-cre; Bnc2^f/f^ mice, compared with littermate control (Fig. 6E,F). Additionally, EdU staining results also showed that Prx1-cre; Bnc2^f/f^ mice developed disorders at the stage of periosteum thickening, and the proportion of EdU-positive periosteal cells decreased significantly (Fig. 6G,H). In addition, a significant increase in EdU^+^BNC2-EGFP^+^ cells was also detected in Bnc2-EGFP mice after fracture (Appendix Fig. S8A,B). In addition, periosteal cells were cultured in vitro, and the proliferation ability of these cells was assessed by CCK8 assay and cell cycle analysis. The proliferation capacity of periosteal cells was significantly decreased in Prx1-cre; Bnc2^f/f^ mice (Fig. 6I,J). Overexpression of Bnc2 significantly increased the proliferation capacity of periosteal cells (Appendix Fig. S9A,B). To determine whether BNC2 affects chondrogenesis, we induced the periosteal cells of Prx1-cre; Bnc2^f/f^ mice differentiated into chondrocytes. The results showed that loss of BNC2 did not affect the chondrogenic differentiation (Appendix Fig. S9C–E). And depletion of BNC2 with Prx1-cre did not show cartilage phenotype in homeostasis (Appendix Fig. S10). However, we detected a significant disorder in cartilage formation in the callus of Prx1-Cre; Bnc2^f/f^ mice (Fig. 5H,I). In addition, we found that the absence of Bnc2 led to a significant decrease in the proliferation ability of periosteal cells (Fig. 6). Therefore, we believe that the cartilage formation disorder during the fracture repair process in Prx1-Cre; Bnc2^f/f^ mice is mainly caused by the obstruction of precursor cell proliferation. The above results show that loss of Bnc2 in periosteal SSCs can lead to significant impaired proliferation in the initial stage of fracture repair, suggesting that Bnc2 probably regulates the fracture healing process through the activation of periosteal SSCs.Figure 6Prx1-cre; Bnc2^f/f^ mice have a significant decrease in SSC activation in the early stages of fracture healing.(A) Schematic diagram of SSCs analysis in Prx1-cre; Bnc2^f/f^ mice after fracture. (B, C) Flow analysis of the proportion of SSCs in undamaged periosteum and 3 dpf callus in Prx1-cre; Bnc2^f/f^ mice. Data were presented as the means ± SEM. n = 6. Unpaired t-test. (D) Schematic diagram of in vivo EdU labeling in Prx1-cre; Bnc2^f/f^ mice after fracture. (E, F) Flow analysis of the proportion of EdU-positive precursor cells in undamaged periosteum and 3 dpf callus in Prx1-cre; Bnc2^f/f^ mice. Data were presented as the means ± SEM. n = 6. Unpaired t-test. (G, H) EdU staining and quantitative statistics of the proportion of EdU-positive periosteal cells in 2 dpf sections of Prx1-cre; Bnc2^f/f^ mice. Data were presented as the means ± SEM. n = 6. Unpaired t-test. (I) CCK8 assay of periosteal cells in Prx1-cre; Bnc2^f/f^ mice. Data were presented as the means ± SEM. n = 6. Unpaired t-test. (J) Cell cycle analysis of periosteal cells in Prx1-cre; Bnc2^f/f^ mice. Data were presented as the means ± SEM. n = 3. Unpaired t-test. Source data are available online for this figure.
BNC2 regulates fracture healing by interacting with the NuRD complex
To explore how Bnc2 regulates the proliferation of SSCs, we used mass spectrometry to identify the BNC2 interacting proteins. Silver staining results showed that Flag-BNC2 was significantly enriched, and specific proteins were pulled down (Fig. 7A). Mass spectrometry analysis showed that the components of the NuRD (nucleosome remodeling and deacetylase) complex were significantly enriched in the Flag-BNC2 immunoprecipitation samples (Fig. 7B). Further co-immunoprecipitation experiments confirmed that there was a significant interaction between BNC2 and the components of the NuRD complex (Fig. 7C). The NuRD complex is one of the major chromatin remodeling complexes, and plays an important role in regulating gene transcription, genome integrity and cell cycle progression (Basta, 2015). The NuRD complex can regulate gene transcription by binding to enhancers and promoters of genes to reduce the level of histone acetylation. Therefore, we use Prx1-cre; Bnc2^f/+^; Rosa26-LSL-tdTomato and Prx1-cre; Bnc2^f/f^; Rosa26-LSL-tdTomato mice to obtain tdTomato-positive cells on 3 dpf by flow cytometry (Appendix Fig. S11A). The acetylation levels of histone H3 were significantly reduced in tdTomato positive callus cells of Prx1-cre; Bnc2^f/f^; Rosa26-LSL-tdTomato mice (Fig. 7D). Meanwhile, the decreased H3 acetylation in periosteal cells of Prx1-cre; Bnc2^f/f^ mice was also demonstrated by pan-H3ac immunohistochemical staining (Fig. 7E). Then, we tried to demonstrate whether the impairment of fracture healing in Prx1-cre; Bnc2^f/f^ mice could be alleviated by intervention with the NuRD complex. Through intraperitoneal injection of the HDAC inhibitor TSA in Prx1-cre; Bnc2^f/f^ and Bnc2^f/f^ mice, we observed that TSA could partially restore the phenotype of fracture healing disorder in Prx1-cre; Bnc2^f/f^ mice, as manifested by a partial increase in callus volume (Fig. 7F,G) and increased bone volume (Fig. 7H,I) at the callus on 14 dpf. The callus composition of Prx1-cre; Bnc2^f/f^ mice treated by TSA is basically the same as that of Bnc2^f/f^ mice, except that the overall volume is still smaller than Bnc2^f/f^ mice, as demonstrated by SOFG staining (Fig. 7J).Figure 7BNC2 regulates H3ac of callus cells through the NuRD complex.(A) Silver staining results of Flag-BNC2 pull-down. (B) Enrichment of components of the NuRD complex in LC-MS/MS results of Flag-BNC2 pull-down. (C) Co-immunoprecipitation to verify the interaction between BNC2 and components of the NuRD complex. (D) Pan-H3ac was detected by WB in Lin^-^ Prx1-cre; Rosa26-LSL-tdTomato^+^ cells of Prx1-cre; Bnc2^f/f^ mice in 3 dpf. (E) Pan-H3ac levels in periosteal cells of Prx1-cre; Bnc2^f/f^ mice on 2 dpf were determined by immunohistochemistry. CB, cortical bone. (F, G) X-Ray results (F) and callus index (G) on 14 dpf in TSA-treated Prx1-cre; Bnc2^f/f^ mice. Data were presented as the means ± SEM. n = 6, 6, 8, 9. Unpaired t-test. The dotted line represents the edge of the callus. (H, I) Micro-CT scanning results (H) and bone volume (I) at callus of 14 dpf in TSA-treated Prx1-cre; Bnc2^f/f^ mice. Data were presented as the means ± SEM. n = 6, 6, 8, 9. Unpaired t-test. (J) SOFG staining of sections on 14 dpf in TSA-treated Prx1-cre; Bnc2^f/f^ mice. (K) ATAC-seq results of SSCs on 3 dpf in Prx1-cre; Bnc2^f/f^ mice showed chromatin accessibility on the gene body and heat maps of peaks. (L) GO analysis of down-regulated peaks in the SSCs on 3 dpf of Prx1-cre; Bnc2^f/f^ mice. Fisher’s exact test. Source data are available online for this figure.
To further explore the pathways through which BNC2 regulates the proliferation of SSCs in fracture healing, SSCs of Prx1-cre; Bnc2^f/f^ and control mice on 3 dpf were obtained by flow cytometry for ATAC-seq. We found a significant decrease in chromatin accessibility near the gene transcription start sites (TSS) of callus SSCs in Prx1-cre; Bnc2^f/f^ mice (Fig. 7K). The pathway related to transcriptional activation, cell proliferation and differentiation were significantly enriched through GO analysis of the down-regulated peaks in Prx1-cre; Bnc2^f/f^ mice callus SSCs (Fig. 7L). These data suggest that the loss of Bnc2 leads to a decrease in chromatin accessibility of genes related to transcriptional activation and cell proliferation in callus SSCs, thus hindering the proliferation of callus SSCs.
These results indicate that BNC2 regulates the acetylation levels of histone H3 through its interaction with the NuRD complex, thus affecting chromatin accessibility and ultimately regulating gene transcriptional activation. Pharmacological inhibition of HDAC could partially alleviate the phenotype of fracture healing disorder in Prx1-cre; Bnc2^f/f^ mice.
Discussion
In this study, we found that the expression of Bnc2 was significantly upregulated in SSCs in the early stages of fracture. By lineage tracing of constructed fluorescent reporter mice, we found that Bnc2 could label periosteal cells and could be fully involved in fracture repair, ultimately forming neo-bone and periosteum. However, Bnc2-labeled periosteal cells do not differentiate into osteoblasts and osteocytes in homeostasis, indicating that they are in a resting state and can be activated and participate in repair only when receiving injury stimulation.
In order to explore how Bnc2 regulates fracture repair, we constructed a conditional knockout mouse model of Bnc2. The fracture repair of Prx1-cre; Bnc2^f/f^ mice was significantly abnormal, which was manifested as periosteal cell proliferation disorder and subsequent endochondral osteogenesis obstruction. However, Ocn-cre; Bnc2^f/f^ and LepR-creER; Bnc2^f/f^ mice did not have the above phenotype, suggesting that Bnc2 specifically promoting fracture repair in stem/precursor cells of periosteum. Moreover, we found that the chondrogenic differentiation characteristics of periosteal cells of Prx1-cre; Bnc2^f/f^ mice were unchanged. This may imply that the loss of Bnc2 leads to the premature entry of periosteal cells into the differentiation stage, and the cause of impaired fracture healing in Prx1-cre; Bnc2^f/f^ mice may be due to the decreased proliferation ability of precursor cells. Significant downregulation of BNC2 was detected in both ZKSCAN3 knockout and CLOCK knockout human mesenchymal stem cell senescence models (Hu et al, 2020; Liang et al, 2021). Therefore, we can hypothesize that the deletion of Bnc2 leads to the premature aging of periosteal cells, which is manifested by a decrease in proliferative ability and early entry into differentiation, which ultimately leads to the lack of sufficient precursor cells in the later stage of fracture repair and eventually causes impaired fracture repair. This study demonstrates that only Prx1-cre; Bnc2^f/f^ mice exhibit an impaired fracture repair phenotype, whereas LepR-creER; Bnc2^f/f^ mice complete fracture repair normally. This indicates that Bnc2 can serve as a factor that specifically regulates the functional involvement of periosteal cells in fracture repair. However, lineage tracing results indicate that Bnc2 gene primarily labels periosteal cells but also labels a small number of metaphyseal bone marrow cells, suggesting that Bnc2 is not a perfect marker gene for periosteal cells. More ideal marker genes for periosteal stem cells may yet be identified.
Epigenetic regulation is crucial for bone development and repair. Histone H3K4 tri-methyltransferase Ash1l regulates the differentiation of mesenchymal stem cells by epigenetic means (Yin et al, 2019). Ptip regulates the proliferation and differentiation of COL2-positive precursor cells by decreasing the H3K27ac level in the Pgk1 promoter region (Liang et al, 2024). However, the epigenetic control of skeletal stem cells fate remains to be studied. We found that Bnc2 regulates periosteal cell proliferation by changing chromatin accessibility of transcriptional activators through interaction with the NuRD complex, and thus affects fracture repair, which is actually the regulation of histone acetylation on Bnc2-positive periosteal cells from resting to activated state.
We found that Bnc2^+^ periosteal cells are crucial for fracture repair, but not all Bnc2^+^ periosteal cells are SSCs. Therefore, by combining with the identified SSC marker gene, we can assume that Bnc2^+^ SSCs in the periosteum are mainly involved in fracture repair, while Bnc2^-^ SSC is mainly involved in homeostasis maintenance. This is the direction we continue to explore. In addition, we found that BNC2 regulated the expression of genes related to proliferation and differentiation pathways through interaction with the NuRD complex, but the key direct downstream target genes of BNC2 were not identified. And due to the wide range of action of HDAC inhibitors, while promoting fracture repair, HDAC inhibitors may have temporarily unknown side effects. Subsequently, we will further identify the target genes of BNC2 to achieve more precise regulation and provide new targets for the treatment of nonunion of bone fractures.
In summary, we found that Bnc2 specifically labeled resting periosteal cells in homeostasis, and that Bnc2-positive periosteal cells contributed adequately to endochondral osteogenesis during fracture repair. Meanwhile, BNC2 regulates the proliferation of periosteal SSCs in an epigenetic manner through interaction with the NuRD complex in the initial stage of fracture repair (Appendix Fig. S12A). This study suggests that intervention in histone acetylation may promote the recovery of some bone repair defects caused by stem cell proliferation disorders.
Methods
Reagents and tools tableReagent/resourceReference or sourceIdentifier or catalog number Experimental models Bnc2-P2A-EGFP miceGemPharmatechCustomizedBnc2-P2A-creER miceGemPharmatechCustomizedBnc2^f/f^ miceGemPharmatechCustomizedOcn-creER miceBIOCYTOGENCustomizedPrx1-Cre miceJackson LaboratoryB6.Cg-Tg(Prrx1-cre)1Cjt/JRosa26-LSL-tdTomato miceZilong Qiu (Shanghai Jiao Tong University, Shanghai, China)LepR-dreER; Rosa26-RSR-GFP miceBo O. Zhou (CAS Center for Excellence in Molecular Cell Science, Shanghai, China)LepR-creER miceBo O. Zhou (CAS Center for Excellence in Molecular Cell Science, Shanghai, China)Ocn-cre micePeiqiang Su (The First Affiliated Hospital, Sun Yat-sen University, Guangzhou, China)Prx1-creER miceBaojie Li (Shanghai Jiao Tong University) Recombinant DNA Phage-Flag-Bnc2This studyPlenti-HA-Bnc2This studyPlex-Flag-Mta1This studyPlex-Flag-Mta2This studyPlex-Flag-Rbbp4This studyPlex-Flag-Rbbp7This studyPlex-Flag-Gatad2aThis studyPlex-Flag-Gatad2bThis studyPlex-Flag-Mbd3This studyPlex-Flag-Hdac1This study Antibodies Rabbit anti-COL2BosterCat # BA0533Goat anti-OPNR&DCat # AF808Rabbit anti-pan-H3acAbcamCat # ab300641Rabbit anti-H3Cell Signaling TechnologyCat #9715Mouse anti-FlagSigmaCat #F3165Rabbit anti-HACell Signaling TechnologyCat #3724Donkey anti-rabbit 488Molecular ProbesCat # A21206Donkey anti-goat Cy3Jackson ImmunoResearchCat #705-165-147PerCP/Cy5.5-conjugated anti-CD45BiolegendCat #103132PerCP/Cy5.5-conjugated anti-CD31BiolegendCat #102420PerCP/Cy5.5-conjugated anti-Ter119BiolegendCat #116228FITC-conjugated anti-mouse 6C3/Ly-51BiolegendCat #108305Brilliant Violet 605™-conjugated anti-mouse CD90.2BiolegendCat #140317PE/Cy7-conjugated anti-mouse CD105BiolegendCat #120409APC-conjugated anti-mouse CD200BiolegendCat #123809 Oligonucleotides and other sequence-based reagents PCR PrimersThis study Chemicals, enzymes and other reagents Trichostatin AMCECat # HY-15144FLAG-M2 peptideSigmaCat #F3290Protease inhibitorsMCECat # HY-K0010Protein A/G PLUS-AgaroseSanta CruzCat # sc-2003Flag-M2 agarose beadsSigmaCat # A2220Silver staining kitSangon BiotechCat # C510027Red blood cell lysis bufferBeyotimeCat #C3702α-MEMCorningCat #10-022-CVRCollagenaseSigmaCat # C0130Dispase IISigmaCat # D4693EdURiboBioCat # C00054Cell-Light EdU Apollo643 In Vitro KitRiboBioCat # C10310-2Cell-Light EdU Apollo488 In Vitro KitRiboBioCat # C10310-3DAPISigmaCat #D9542Fluorescence mounting mediumDAKOCat # S3023TamoxifenSigmaCat # T5648OCT-freeze mediumEprediaCat #22-110-6502 Software Flowjo v10.8.1BDGraphPad Prism v8GraphPadIllustratorAdobe Other VS-120 microscopeOlympusSp8 STED microscopeLeicaLSM980 microscopeZEISSCytoflexBeckman CoulterBD Arial FusionBDMX2 X-ray systemVet Ray TechnologySkyScan 1272Bruker
Animals
Bnc2-P2A-EGFP, Bnc2-P2A-creER, and Bnc2^f/f^ mice were generated with the assistance of GemPharmatech. Ocn-creER mice were generated with the assistance of BIOCYTOGEN. Prx1-Cre mice was purchased from the Jackson Laboratory. Rosa26-LSL-tdTomato mice were kindly provided by Dr. Zilong Qiu. LepR-dreER; Rosa26-RSR-GFP and LepR-creER mice were kindly provided by Dr. Bo O. Zhou. Ocn-cre mice were kindly provided by Dr. Peiqiang Su. Prx1-creER mice were kindly provided by Dr. Baojie Li. CreER expressing mice were induced by intraperitoneal injection daily with 100 mg/kg mice of tamoxifen dissolved in corn oil for three or six times. All utilized mice were maintained on the C57/BL6 background, both male and female mice were analyzed. All animals were housed under specific pathogen-free conditions.
Mouse femoral fracture model
Six to ten-week-old mice were used for fracture modeling. Mice were anesthetized with Avertin. Intramedullary fixation was performed on the femur with a pin, and the fracture was performed in the middle of the femur with a dentist’s microdrill. After the muscle was repositioned, absorbable sutures were used to close the skin. X-ray were performed weekly after the fracture with the MX2 X-ray system, and micro-CT scans were performed 4 or 5 weeks after surgery with Bruker, SkyScan 1272. Callus index was defined as the ratio of the maximum diameter of the callus to the diameter of the femur.
Mouse bone drill hole model
The drill hole model is used to characterize the bone injury repair model, mainly involved in the intramembrane osteogenesis (Colnot, 2009). Briefly, a skin incision was made in the middle of the tibia, the subcutaneous tissue was bluntly separated, the tibia was exposed, and then a 0.7 mm diameter needle was used to drill through one side of the cortical bone. Once this was done, the skin was repositioned and sutured. Bone formation was detected by micro-CT on the 14th day after surgery.
Histological analysis
Freshly isolated mouse samples were fixed at 4 °C with 4% PFA for 48 h, and then decalcified at 350 mM EDTA, PH6.5 after washing with PBS. For paraffin sections, the samples were dehydrated by concentration gradient ethanol and then paraffin-embedded and sliced at a thickness of 7 μm. For frozen sections, samples were dehydrated with 30% sucrose and then embedded with tissue OCT-freeze medium (Epredia, 22-110-6502) and sliced at a thickness of 20 μm.
Immunohistochemistry and immunofluorescence staining
The frozen sections were restored to room temperature and rehydrated with PBS. Then the antigenic repair was performed with protease K in a water bath at 37 °C for 10 min. The sections were blocked and permeabilized with PBS containing 10% horse serum and 0.3% Triton X-100 for 1 hour at room temperature. The primary antibody (rabbit anti-COL2, Boster, BA0533, 1:200; goat anti-OPN, R&D, AF808, 1:500; rabbit anti-pan-H3ac, Abcam, ab300641, 1:200) was incubated at 4 °C overnight and washed with PBS. The secondary antibody coupled with fluorescein (donkey anti-rabbit 488, Molecular Probes, A21206, 1:1000; donkey anti-goat Cy3, Jackson ImmunoResearch, 705-165-147, 1:1000) was incubated at room temperature for 1 hour and washed with PBS. The nucleus was counterstained with DAPI (Sigma, D9542) and finally mounted with fluorescence mounting medium (Dako, S3023). Stained images were acquired with Olympus VS-120, Leica Sp8 STED and ZEISS LSM980 microscopes.
In vivo EdU labeling and staining
EdU solution (5 mg/mL) (RiboBio, C00054) was intraperitoneally injected into mice with 10 μL/g of mice. After 12 h of injection, the mice were sacrificed, and the hind limbs were collected. The section sample was prepared according to the conventional paraffin section preparation method. The EdU staining procedure was performed according to the kit’s instructions (RiboBio, C10310-2 and C10310-3).
Isolation of periosteal cells
The muscles of the femur and tibia were removed first, then both ends of the bone were coated with a low melting agarose (10% in TAE buffer) and digested with minimum essential medium alpha (α-MEM; Corning, 10-022-CVR) containing 1 mg/mL collagenase (Sigma, C0130) and 2 mg/mL Dispase II (Sigma, D4693) at 37 °C, 150 rpm, 30 min × 2. The cell suspension can be filtered through a 70-μm cell strainer (Falcon, 352350) for flow cytometry, sorting, or induction of differentiation.
Flow cytometry
In brief, freshly isolated primary cells were treated with red blood cell lysis buffer (Beyotime, C3702) and then stained with antibodies as following: PerCP/Cy5.5-conjugated anti-CD45 (Biolegend, 103132), PerCP/Cy5.5-conjugated anti-CD31 (Biolegend, 102420), PerCP/Cy5.5-conjugated anti-Ter119 (Biolegend, 116228), FITC-conjugated anti-mouse 6C3/Ly-51 (Biolegend, 108305), Brilliant Violet 605™-conjugated anti-mouse CD90.2 (Biolegend, 140317), PE/Cy7-conjugated anti-mouse CD105 (Biolegend, 120409), APC-conjugated anti-mouse CD200 (Biolegend, 123809). The above primary antibody was diluted by 1:500 and incubated on ice for 30 min away from light. After washing with 2% FBS PBS, flow cytometry was performed with Cytoflex (Beckman Coulter) and cell sorting was performed with BD Arial Fusion (BD Biosciences), and data were analyzed with FlowJo software.
Flag-BNC2 pull-down
Flag-BNC2 plasmid was transfected into 293FT cells for the overexpression of the Flag-BNC2 protein. Two days after transfection, cells were collected and treated with EBC buffer containing protease inhibitors (MCE, HY-K0010). Then, protein A/G PLUS-Agarose (Santa Cruz, sc-2003) was added to the cell lysate and shaken at 4 °C for 3 h for preincubation, and the supernatant was transferred after centrifugation. Added Flag-M2 agarose beads (Sigma, A2220) and incubated overnight at 4 °C. Discarded the supernatant after centrifuging, rinsed with pre-cooled EBC three times, then eluted protein with FLAG-M2 peptide (100 μg/mL) (Sigma, F3290). After SDS-PAGE electrophoresis, the samples were silver-stained to detect protein enrichment (Sangon Biotech, C510027). Finally, the eluted liquid samples were identified by LC-MS/MS.
Pharmacological inhibition of HDACs in mice
Bnc2^f/f^ and Prx1-cre; Bnc2^f/f^ mice were randomly grouped and intraperitoneally injected daily with TSA (MCE, HY-15144) or solvent as controls at 10 mg/kg starting on day 1 after fracture and continuing until day 14 after surgery according to Amy M Avila’s research(Avila et al, 2007). The mice were sacrificed at 14 dpf, and the fracture repair status was examined by X-ray, micro-CT, and histological analysis.
Micromass culture
Periosteal cells were digested, resuspended at 1 × 10^7^ cells/mL, and plated in a 12.5 μL droplet of cell suspension in the center of a 24-well-plate; the plate was placed at 37 °C for 2 h, and chondrogenic differentiation medium, which contained DMEM (Corning, 10-013-CVR), 10 ng/mL TGFβ3 (Peprotech, 100-36E), 100 nM dexamethasone (Sigma, D1756), 50 μg/mL L-ascorbic acid 2-phosphate (Sigma, A8960), 1 mM sodium pyruvate (Sigma, 25-000-CIR), 40 μg/mL proline (Sigma, P5607), and 1% ITS (Cyagen, ITSS-10201-10), was then gently added. On the fourth day of induced differentiation, micromasses were acidified with 0.1 N HCl and were then stained with 1% Alcian blue (Sigma, A5268) or RNA extraction to detect the expression of marker genes.
Quantitative real-time PCR
Total RNA was isolated from different tissues and cells with TRIzol Reagent (Sigma, T9424) and reverse-transcribed with a PrimeScript RT Reagent Kit (Takara, RR037A). Real-time fluorescence quantitative PCR was performed in a Bio-Rad CFX Connect Real-Time System. The primer sets used were Bnc2: sense 5’-GTGATCAGTGTAAACATGGCTGGGT-3’, anti-sense 5’-GGATGTATCCTCGGACATAGTCCCT-3’; Sox9: sense 5’- TTCCTCCTCCCGGCATGAGTG-3’, anti-sense 5’- CAACTTTGCCAGCTTGCACG-3’; Col2: sense 5’- CGGTCCTACGGTGTCAGG-3’, anti-sense 5’-GCAGAGGACATTCCCAGTGT-3’; Acan: sense 5’- AATCCCCAAATCCCTCATAC-3’, anti-sense 5’- CTTAGTCCACCCCTCCTCAC-3’; Col10: sense 5’-TTCTGCTGCTAATGTTCTTGACC-3’, anti-sense 5’- GGGATGAAGTATTGTGTCTTGGG-3’; Hprt: sense 5’-GTTAAGCAGTACAGCCCCAAA-3’, anti-sense 5’- AGGGCATATCCAACAACAAACTT-3’.
Bulk RNAseq
RNA purification, reverse transcription, library construction and sequencing were performed at Shanghai Majorbio Bio-pharm Biotechnology Co., Ltd. (Shanghai, China) according to the manufacturer’s instructions. The transcriptome library was prepared following Illumina® Stranded mRNA Prep, Ligation (San Diego, CA) using 1 μg of total RNA. After quantification by Qubit 4.0, the sequencing library was performed on the NovaSeq X Plus platform (PE150) using NovaSeq Reagent Kit (NovaSeq 6000). The raw paired-end reads were trimmed and quality-controlled by fastp with default parameters. Then, clean reads were separately aligned to the reference genome with orientation mode using HISAT2 software. The mapped reads of each sample were assembled by StringTie in a reference-based approach.
ATAC-seq
The cells were then thawed in a 37 °C water bath, pelleted, and washed with cold PBS. The cell pellets were resuspended in lysis buffer, pelleted, and tagmented using the Assembled Tn5 Transposome and Tagmentation buffers (Active Motif). Tagmented DNA was then purified using the QIAGEN PCR Purification Kit, amplified with ten cycles of PCR, and purified using the QIAGEN PCR Purification Kit. The resulting material was quantified with Qubit (Thermo Fisher) and sequenced with PE150 sequencing on the Illumina platform. Raw reads were filtered to obtain high-quality clean reads by removing sequencing adapters, short reads (length <35 bp), and low-quality reads using trim-galore (v0.6.4). Then FastQC (v0.11.9) and Multiqc (v1.8) is used to ensure high reads quality. The clean reads were mapped to the mouse genome (assembly mouse genome mm10) using the Burrow-Wheeler Aligner (BWA v0.7.17) software. PCR duplicates were removed using Picard (v2.22.2-0). Peak detection was performed using the MACS2 (v2.2.6) peak finding algorithm with 0.05 set as the q value cutoff and −100 set as the shift. Annotation of peak sites to gene features was performed using the Homer annotatePeaks.pl (v4.10). TSS heatmap was generated using deepTools (v3.4.3) computeMatrix reference-point.
Statistics
Data were derived from independently obtained datasets and expressed as mean ± SEM. Two groups were compared using paired or unpaired two-tailed t-tests. The number of samples shown in each figure is the number of biological replicates.
Ethics statement
We comply with all ethical regulations regarding animal testing and research. All animal experiments were conducted in the Animal Laboratory of Shanghai Institute of Biochemistry and Cell Biology, CAS Center for Excellence in Molecular Cell Science, in accordance with the protocol approved by the Committee for Animal Health and Use of Shanghai Institute of Biochemistry and Cell Biology, CAS Center for Excellence in Molecular Cell Science, Chinese Academy of Sciences (Approval number: SIBCB-S350-2403-16).
Supplementary information
Appendix Peer Review File Source data Fig. 1 Source data Fig. 2 Source data Fig. 3 Source data Fig. 4 Source data Fig. 5 Source data Fig. 6 Source data Fig. 7
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang L, Ren Q, Chen S, Lou L, Hu X, Xing W, Suo J, Sun J, Greenblatt MB, Feng H, et al (2025) Gene expression omnibus GSE 249206 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE 249206) [DATASET]