Standardised sporulation methods for Diplodia, Lasiodiplodia and Neofusicoccum
David Hernández-Hernández, Felipe Siverio de la Rosa, Christiaan Grobler, Bernard Slippers

TL;DR
Researchers developed a standardized method to induce sporulation in several fungal species from the Botryosphaeriaceae family, which can help in studying their biology and disease mechanisms.
Contribution
The study introduces a standardized and reproducible method using Vogel’s Minimal Medium for sporulation in Botryosphaeriaceae fungi.
Findings
Vogel’s Minimal Medium successfully induced sporulation in multiple species of Diplodia, Lasiodiplodia, and Neofusicoccum.
Significant variation in sporulation time and yield was observed among and within species.
Reduced sucrose in the medium affected sporulation differently across species.
Abstract
Members of the Botryosphaeriaceae are widespread fungal pathogens responsible for economically important diseases in woody plants. Despite the relevance of conidia production for understanding pathogen biology, infection processes, and disease epidemiology, sporulation in vitro remains unpredictable and inconsistent across species. In this study, we evaluated the efficacy of Vogel’s Minimal Medium (VMM) for inducing pycnidial and conidial development, which has recently been shown to be effective for this purpose in Diplodia sapinea, in species of Diplodia (n = 3), Lasiodiplodia (n = 2), and Neofusicoccum (n = 26). For this purpose, we used 123 isolates recently collected in the Canary Islands (Spain), as well as 67 Neofusicoccum isolates from long-term storage. All isolates were identified through multilocus phylogenetic analysis. The results showed that D. africana, D. mutila, and D.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1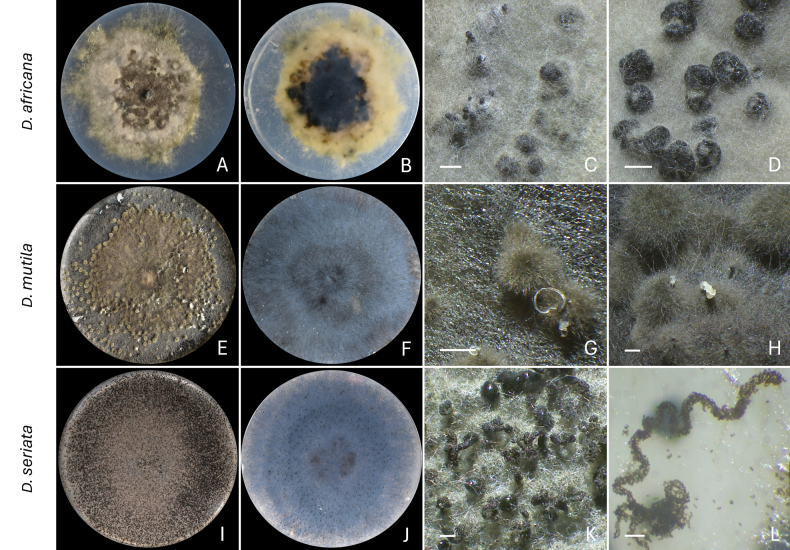 Figure 2
Figure 2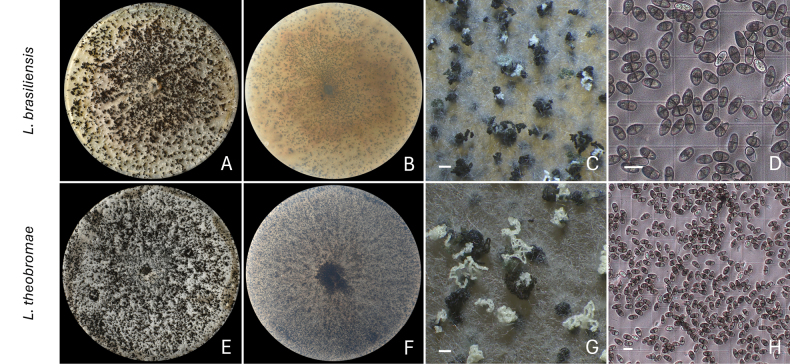 Figure 3
Figure 3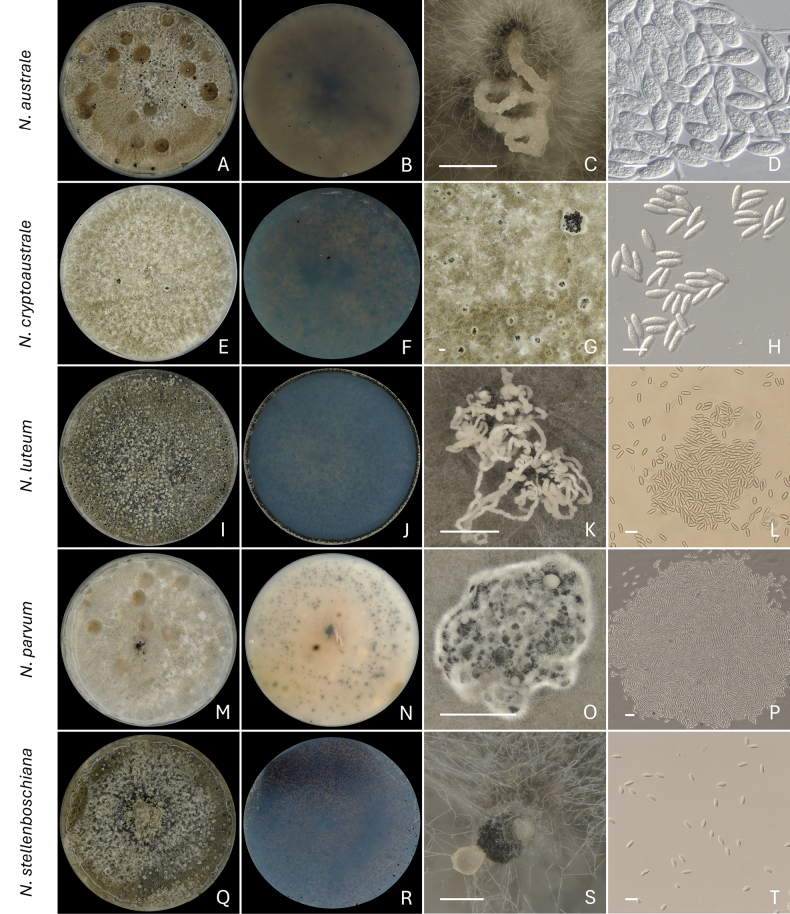 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6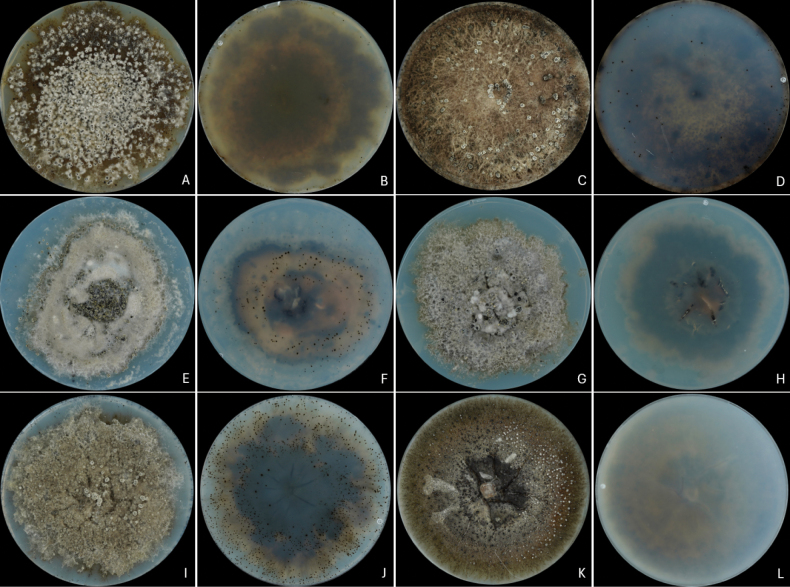 Figure 7
Figure 7| Species | Isolates | Isolates with pycnidia formation | Isolates with conidial discharge | Mean days to initial pycnidia formation | Mean days to initial conidial discharge | Mean days to conidia harvest | Mean conidial concentration (conidia mL-1) |
|---|---|---|---|---|---|---|---|
|
| 12 | 11 (91.7%) | 5 (41.7%) | 14 | 18 | 35 | 0 |
|
| 1 | 1 (100%) | 1 (100%) | 14 | 21 | 35 | 0 |
|
| 2 | 2 (100%) | 2 (100%) | 3 | 7 | 14 | 1.3 × 106 |
|
| 11 | 9 (81.8%) | 9 (81.8%) | 5 | 6 | 21 | 8.7 × 106 |
|
| 1 | 1 (100%) | 1 (100%) | 5 | 6 | 21 | 7.12 × 106 |
|
| 1 | 1 (100%) | 1 (100%) | 5 | 14 | 28 | 3.75 × 105 |
|
| 1 | 1 (100%) | 1 (100%) | 4 | 25 | 35 | 1.25 × 105 |
|
| 23 | 23 (100%) | 23 (100%) | 4 | 7 | 21 | 1.5 × 107 |
|
| 37 | 31 (83.8%) | 22 (59.5%) | 5 | 21 | 35 | 2.31 × 106 |
|
| 34 | 33 (97.1%) | 26 (76.5%) | 5 | 21 | 35 | 2 × 106 |
| Species (n° isolates) | Sucrose concentration in | ||||
|---|---|---|---|---|---|
|
|
|
|
|
| |
| Initial pycnidia formation | 14 d | 8 d | 9 d | 21 d | 21 d |
| First conidial discharge | 18 d | 14 d | NC | NC | 28 d |
| Conidial harvest | 35 d | 35 d | 35 d | 35 d | 35 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 0 | 0 | 0 | 0 | 7.5 × 104 |
|
|
|
|
|
| |
| Initial pycnidia formation | 14 d | 7 d | 8 d | 14 d | 21 d |
| First conidial discharge | 21 d | 14 d | 14 d | NC | NC |
| Conidial harvest | 35 d | 28 d | 28 d | 28 d | 28 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 0 | 0 | 0 | 0 | 0 |
|
|
|
|
|
| |
| Initial pycnidia formation | 3 d | 5 d | 5 d | 5 d | 7 d |
| First conidial discharge | 7 d | 7 d | 7 d | 7 d | 10 d |
| Conidial harvest | 14 d | 14 d | 14 d | 14 d | 14 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 1.3 × 106 | 9.29 × 106 | 1.36 × 107 | 1.07 × 107 | 3.37 × 105 |
|
|
|
|
|
| |
| Initial pycnidia formation | 5 d | 3 d | 3 d | 3 d | 4 d |
| First conidial discharge | 6 d | 7 d | 7 d | 7 d | 7 d |
| Conidial harvest | 21 d | 21 d | 21 d | 21 d | 21 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 8.7 × 106 | 1.44 × 107 | 1.09 × 107 | 1.28 × 107 | 1.84 × 106 |
|
|
|
|
|
| |
| Initial pycnidia formation | 5 d | 3 d | 3 d | 3 d | 3 d |
| First conidial discharge | 6 d | 7 d | 7 d | 7 d | 7 d |
| Conidial harvest | 21 d | 21 d | 21 d | 21 d | 21 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 7.12 × 106 | 2.01 × 107 | 9.25 × 106 | 9.18 × 106 | 1.83 × 106 |
|
|
|
|
|
| |
| Initial pycnidia formation | 5 d | 6 d | 10 d | 10 d | 14 d |
| First conidial discharge | 14 d | 14 d | 14 d | 14 d | 21 d |
| Conidial harvest | 28 d | 21 d | 28 d | 49 d | 49 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 3.75 × 105 | 2.7 × 106 | 6.25 × 105 | 0 | 0 |
|
|
|
|
|
| |
| Initial pycnidia formation | 4 d | 6 d | 6 d | 6 d | 10 d |
| First conidial discharge | 21 d | 21 d | 21 d | 28 d | NC |
| Conidial harvest | 21 d | 49 d | 49 d | 49 d | 49 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 1.25 × 105 | 3 × 105 | 1.75 × 105 | 0 | 0 |
|
|
|
|
|
| |
| Initial pycnidia formation | 4 d | 4 d | 6 d | 6 d | 6 d |
| First conidial discharge | 7 d | 7 d | 7 d | 7 d | 14 d |
| Conidial harvest | 21 d | 21 d | 35 d | 35 d | 35 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 1.5 × 107 | 7.21 × 106 | 2.05 × 106 | 1.7 × 106 | 0 |
|
|
|
|
|
| |
| Initial pycnidia formation | 5 d | 6 d | 6 d | 6 d | 17 d |
| First conidial discharge | 21 d | 21 d | 21 d | 21 d | 35 d |
| Conidial harvest | 28-42 d | 35-49 d | 35-49 d | 35-49 d | 35-49 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 2.31 × 106 | 9 × 105 | 6.56 × 105 | 3.17 × 105 | 0 |
|
|
|
|
|
| |
| Initial pycnidia formation | 5 d | 6 d | 6 d | 6 d | 10 d |
| First conidial discharge | 21 d | 21 d | 21 d | 21 d | 21 d |
| Conidial harvest | 28-42 d | 49 d | 49 d | 49 d | 49 d |
| Mean conidial concentration across all selected isolates (conidia mL-1) | 2 × 106 | 1.03 × 106 | 1.25 × 106 | 3.5 × 105 | 0 |
|
|
|
|---|---|
|
| 0 |
|
| – |
|
| 0.5–0.25 |
|
| 1 |
|
| 1 |
|
| 1 |
|
| 1 |
|
| 1.5 |
|
| 1.5 |
|
| 1.5 |
|
|
|
|
|
|
|
|
| |||
|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
| |||||||
| CMW 13446 | Venezuela | S. Mohali | 2003 |
|
|
|
|
| ||
|
| Venezuela | S. Mohali | 2003 |
|
|
|
|
| ||
| CMW 821 |
| South Africa | M.J. Morris | 1986 |
|
|
|
|
| |
| CMW 1133 |
| South Africa | A. Smith | 1996 |
|
|
|
|
| |
| CMW 1187 | South Africa | A. Smith | 1998 |
|
|
|
|
| ||
|
| Australia | M.J. Wingfield | 2000 |
|
|
|
|
| ||
| CMW 6838 | Australia | M.J. Wingfield | 2000 |
|
|
|
|
| ||
| CMW 6853 |
| Australia | M.J. Wingfield | 2000 |
|
|
|
|
| |
| CMW 9073 | Australia | J. Roux & D. Guest | 1999 |
|
|
|
|
| ||
| CMW 28320 |
| Cameroon | D. Begoude & J. Roux | 2007 |
|
|
|
|
| |
|
|
| Cameroon | D. Begoude & J. Roux | 2007 |
|
|
|
|
| |
|
|
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 14056 |
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
|
|
| South Africa | H.M. Maleme | 2006 |
|
|
|
|
| |
| CMW 54123 | Indonesia | F. Jami | 2018 |
|
|
|
| – | ||
| CMW 54139 | Indonesia | F. Jami | 2018 |
|
|
|
| – | ||
| CMW 54189 | Indonesia | F. Jami | 2018 |
|
|
|
| – | ||
| CMW 6217 |
| Australia | M.J. Wingfield | 2000 |
|
|
|
| – | |
| CMW 6229 |
| Australia | M.J. Wingfield | 2000 |
|
|
|
| – | |
|
|
| Australia | M.J. Wingfield | 2000 |
|
|
|
| – | |
| CMW 6543 | Australia | M.J. Wingfield | 2000 |
|
|
|
|
| ||
| CMW 6233 |
| Australia | M.J. Wingfield | 2000 |
|
|
|
|
| |
|
|
| South Africa | H. Smith | 2001 |
|
|
|
| – | |
|
|
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 14140 |
| South Africa | D. Pavlic | 2003 |
|
|
|
|
| |
| CMW 41228 |
| South Africa | J. A. Osorio | 2012 |
|
|
|
|
| |
| CMW 41613 |
| South Africa | J. A. Osorio | 2011 |
|
|
|
|
| |
|
|
| South Africa | J. A. Osorio & J. Roux | 2012 |
|
|
|
|
| |
| CMW 14071 |
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 7797 |
| Australia | G.I. Johnson | 1988 |
|
|
|
|
| |
|
|
| Australia | G.I. Johnson | 1988 |
|
|
|
| – | |
|
|
| South Africa | J. A. Osorio & J. Roux | 2012 |
|
|
|
|
| |
| CMW 42355 |
| South Africa | J. A. Osorio | 2011 |
|
|
|
|
| |
| CMW 42481 |
| South Africa | J. A. Osorio | 2011 |
|
|
|
|
| |
| CMW 13998 |
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 54121 | Indonesia | F. Jami | 2018 |
|
|
|
| – | ||
| CMW 54144 | Indonesia | F. Jami | 2018 |
|
|
|
| – | ||
| CMW 54212 | Indonesia | F. Jami | Unknown |
|
|
|
| – | ||
| CMW 54187 | Indonesia | F. Jami | 2018 |
|
|
|
| – | ||
| CMW 14029 |
| South Africa | M.J. Wingfield | 2002 |
|
|
|
|
| |
| CMW 14087 |
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 9071 | Australia | M.J. Wingfield | 2000 |
|
|
|
|
| ||
| CMW 9080 |
| New Zealand | G.J. Samuels | 2005 |
|
|
|
|
| |
|
|
| New Zealand | G.J. Samuels | 2005 |
|
|
|
|
| |
| CMW 13350 | Venezuela | L. Cedeno | 2002 |
|
|
|
|
| ||
| CMW 41225 |
| South Africa | J.A. Osorio | 2012 |
|
|
|
|
| |
| CMW 41361 |
| South Africa | J.A. Osorio | 2012 |
|
|
|
|
| |
| CMW 41368 |
| South Africa | J.A. Osorio | 2012 |
|
|
|
|
| |
|
|
| South Africa | J. Roux & M.L. Ndove | 2008 |
|
|
|
|
| |
| CMW 35499 |
| South Africa | J. Roux & M.L. Ndove | 2008 |
|
|
|
|
| |
| CMW 7054 |
| USA | E. Stevens | 2001 |
|
|
|
|
| |
|
| USA | B. Slippers & G. Hudler | 2000 |
|
|
|
|
| ||
| CMW 13986 |
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 41363 |
| South Africa | J.A. Osorio | 2012 |
|
|
|
|
| |
| CMW 26683 |
| South Africa | D. Begoude & J. Roux | 2007 |
|
|
|
|
| |
|
|
| South Africa | D. Begoude & J. Roux | 2007 |
|
|
|
|
| |
| CMW 13991 |
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 14060 |
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
|
|
| South Africa | D. Pavlic | 2002 |
|
|
|
|
| |
| CMW 23790 |
| South Africa | H.M. Maleme | 2006 |
|
|
|
| – | |
| CMW 35471 |
| South Africa | E.M. Cruywagen & M.L. Ndove | 2009 |
|
|
|
|
| |
| CMW 35481 |
| South Africa | E.M. Cruywagen & M.L. Ndove | 2009 |
|
|
|
|
| |
|
| South Africa | H.M. Maleme | 2006 |
|
|
|
|
| ||
| CMW 37745 |
| South Africa | M.J. Wingfield | Unknown |
|
| – |
| – | |
|
|
| South Africa | M.J. Wingfield | Unknown |
|
|
|
| – | |
| CMW 30605 |
| South Africa | J. Roux | 2008 |
|
|
|
|
| |
| CMW 875 |
| - | M.J. Morris | 1987 |
|
|
|
|
| |
| Species name | Isolate | Isolation year | Days to initial pycnidia formation | Days to initial conidial discharge | Days to conidia harvest | Conidia mL-1 |
|---|---|---|---|---|---|---|
|
| CMW 13446 | 2003 | NP | NC | ND | ND |
|
| 2003 | NP | NC | ND | ND | |
|
| CMW 821 | 1986 | NP | NC | ND | ND |
| CMW 1133 | 1996 | 3 | NC | ND | ND | |
| CMW 1187 | 1998 | NP | NC | ND | ND | |
|
| 2000 | NP | NC | ND | ND | |
| CMW 6838 | 2000 | NP | NC | ND | ND | |
| CMW 6853 | 2000 | NP | NC | ND | ND | |
| CMW 9073 | 1999 | 7 | 18 | 42 | 3.62 × 106 | |
|
| CMW 28320 | 2007 | 17 | NC | ND | ND |
|
| 2007 | NP | NC | ND | ND | |
|
|
| 2002 | NP | NC | ND | ND |
| CMW 14056 | 2002 | 7 | NC | ND | ND | |
|
|
| 2006 | 7 | 30 | 42 | 0 |
|
| CMW 54123 | 2018 | 19 | 30 | 42 | 1.5 × 105 |
| CMW 54139 | 2018 | NP | NC | ND | ND | |
| CMW 54189 | 2018 | 10 | 30 | 42 | 1.75 × 105 | |
|
| CMW 6217 | 2000 | 10 | NC | ND | ND |
| CMW 6229 | 2000 | 10 | NC | ND | ND | |
|
| 2000 | 19 | 30 | 42 | 1.24 × 107 | |
| CMW 6543 | 2000 | 19 | 30 | 42 | 1.27 × 106 | |
|
| CMW 6233 | 2000 | 19 | 30 | 42 | 0 |
|
| 2001 | 19 | 30 | 42 | 6.75 × 105 | |
|
|
| 2002 | NP | NC | ND | ND |
| CMW 14140 | 2003 | NP | NC | ND | ND | |
|
| CMW 41228 | 2012 | 4 | 32 | 42 | 6.75 × 105 |
| CMW 41613 | 2011 | 7 | NC | ND | ND | |
|
| 2012 | 7 | NC | ND | ND | |
|
| CMW 14071 | 2002 | NP | NC | ND | ND |
|
| CMW 7797 | 1988 | NP | NC | ND | ND |
|
| 1988 | NP | NC | ND | ND | |
|
|
| 2012 | 7 | NC | ND | ND |
| CMW 42355 | 2011 | NP | NC | ND | ND | |
| CMW 42481 | 2011 | 7 | NC | ND | ND | |
|
| CMW 13998 | 2002 | 38 | NC | ND | ND |
|
| CMW 54121 | 2018 | 7 | NC | ND | ND |
|
| CMW 54144 | 2018 | 7 | 21 | 42 | 1.5 × 105 |
| CMW 54212 | Unknown | 10 | 24 | 42 | 5 × 104 | |
|
| CMW 54187 | 2018 | 4 | 24 | 42 | 0 |
|
| CMW 14029 | 2002 | 14 | NC | ND | ND |
| CMW 14087 | 2002 | 18 | 32 | 42 | 5 × 104 | |
|
| CMW 9071 | 2000 | 7 | NC | ND | ND |
| CMW 9080 | 2005 | NP | NC | ND | ND | |
|
| 2005 | 4 | NC | ND | ND | |
| CMW 13350 | 2002 | NP | NC | ND | ND | |
| CMW 41225 | 2012 | 4 | NC | ND | ND | |
| CMW 41361 | 2012 | NP | NC | ND | ND | |
| CMW 41368 | 2012 | NP | NC | ND | ND | |
|
|
| 2008 | NP | NC | ND | ND |
| CMW 35499 | 2008 | 10 | NC | ND | ND | |
|
| CMW 7054 | 2001 | 24 | NC | ND | ND |
|
| 2000 | NP | NC | ND | ND | |
|
| CMW 13986 | 2002 | NP | NC | ND | ND |
| CMW 41363 | 2012 | NP | NC | ND | ND | |
|
| CMW 26683 | 2007 | NP | NC | ND | ND |
|
| 2007 | NP | NC | ND | ND | |
|
| CMW 13991 | 2002 | NP | NC | ND | ND |
| CMW 14060 | 2002 | 7 | NC | ND | ND | |
|
| 2002 | NP | NC | ND | ND | |
|
| CMW 23790 | 2006 | NP | NC | ND | ND |
| CMW 35471 | 2009 | 14 | NC | ND | ND | |
| CMW 35481 | 2009 | NP | NC | ND | ND | |
|
| 2006 | NP | NC | ND | ND | |
|
| CMW 37745 | Unknown | 10 | 24 | 42 | 5 × 104 |
|
| Unknown | 7 | NC | ND | ND | |
|
| CMW 30605 | 2008 | 7 | 24 | 42 | 5 × 104 |
| CMW 875 | 1987 | NP | NC | ND | ND |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Fungal Plant Pathogen Control · Ocular Infections and Treatments
Introduction
Sporulation plays an essential role in the survival, dissemination, and infection strategies of fungi. These reproductive structures are often highly resistant to desiccation and other environmental stresses, allowing pathogens to survive in soil, on plant surfaces, or within plant tissues for extended periods (Carlile et al. 2001; Huang and Hull 2017). Sporulation also facilitates the ability of pathogens to infect new hosts and their geographic dispersal, which is critical for the epidemiology of plant diseases (West 2014). Studying spore morphology has therefore contributed significantly to understanding fungal biology and ecology.
Spore morphology reflects ecological adaptations that influence dispersal, host interactions, and infection efficiency. Hydrophobic spores adhere more effectively to plant surfaces, while wind-dispersed types dominate open fields, and mucilaginous spores are linked to humid environments (Braun and Howard 1994; Hughes et al. 1999; Calhim et al. 2018; Wang et al. 2021). Beyond dispersal, spores initiate infection by attaching to the plant cuticle through extracellular mucilage enriched with polysaccharides, glycoproteins, and enzymes such as cutinases (Hamer et al. 1988; Mendgen and Deising 1993; Arya et al. 2021). Their morphology can also aid in evading plant immunity, contributing to virulence (Almeida et al. 2019). Thus, spores function not only as propagules but also as active agents in host colonisation – a key consideration for disease control strategies.
Spore size, shape, and texture have historically played an important role in fungal taxonomy (Phillips et al. 2013). Although molecular resources are now essential for accurate species delimitation in fungi, morphological characters remain useful for preliminary identification, including in the Botryosphaeriaceae, which are the focus of this study (Slippers et al. 2017). Small but characteristic variations in the morphology of conidia provide useful diagnostic features at the genus and, in some cases, species level. For example, Diplodia species usually produce ellipsoidal conidia that become dark-walled and may develop fine longitudinal striations; Lasiodiplodia species often have large, thick-walled conidia with prominent longitudinal grooves; whereas Neofusicoccum species generally form fusiform, hyaline conidia that may become pigmented with age (Crous et al. 2006; Phillips et al. 2013; Zhang et al. 2021). Such characteristics may, however, overlap among species, and their usefulness for identification is therefore limited to scenarios in which the taxa present in a given environment are limited and already known.
The Botryosphaeriaceae (Botryosphaeriales, Dothideomycetes) comprise a diverse assemblage of endophytes and opportunistic pathogens associated with woody plants worldwide. Their taxonomy has historically been challenging because many species exhibit overlapping morphological characters and pronounced pleomorphism, with sexual morphs rarely encountered for many taxa and asexual morphs traditionally treated under separate generic names. Modern classifications have therefore relied heavily on multilocus phylogenetic frameworks, which have stabilised generic concepts and enabled robust species delimitation, while also revealing complexes of cryptic species within morphologically conserved groups. Consequently, reliable identification in the Botryosphaeriaceae is now typically based on analyses combining the internal transcribed spacer region (ITS1–5.8S–ITS2; ITS), the translation elongation factor 1-alpha gene (tef1-α), the beta-tubulin gene (tub2), and, for some genera, the DNA-directed RNA polymerase II second-largest subunit gene (rpb2), supported by curated reference datasets and ex-type or representative sequences, rather than morphology alone (Crous et al. 2006; Phillips et al. 2013; Slippers et al. 2013, 2017; Garcia et al. 2021). This taxonomic framework is particularly important when comparing phenotypes across closely related species and genera.
Members of the Botryosphaeriaceae typically sporulate on necrotic plant tissues, producing pycnidiospores (Slippers and Wingfield 2007; Úrbez-Torres 2011). Although the sexual morph has been described, it is rarely observed in nature (Phillips et al. 2012; Obrador-Sánchez and Hernández-Martínez 2020). Sporulation is influenced by multiple abiotic factors such as temperature, humidity, and nutrient availability, which are often species-specific and difficult to predict (Copes and Hendrix 2004; Vieira et al. 2021). Dispersal of these spores is mainly driven by wind, rain splash, and insect activity (Ahimera et al. 2004; Amponsah et al. 2009; Úrbez-Torres et al. 2010b; Billones-Baaijens et al. 2018).
In the Botryosphaeriaceae, studies specifically focused on spore traits or laboratory-based sporulation protocols remain limited. Nevertheless, several investigations have provided insights into the physiological and environmental factors regulating conidial production and germination. For instance, the effects of moisture and temperature have been explored for sporulation of Botryosphaeria dothidea, showing that conidial production is markedly enhanced under high humidity and is optimal at warm temperatures (25–30 °C), whereas drier conditions or suboptimal temperatures significantly reduce sporulation (Sutton and Arauz 1991). Úrbez-Torres et al. (2010a) assessed conidial germination across temperature gradients and found that optimal ranges varied among species. Further, Sammonds et al. (2019) demonstrated that conidia of N. luteum, N. parvum, and B. dothidea germinated equally on hydrophobic and hydrophilic surfaces, with germination enhanced by the presence of cellulose. Collectively, these findings underscore the complex interplay of abiotic cues in modulating sporulation and germination, while also highlighting the methodological challenges associated with standardising spore production. Despite their ecological and epidemiological importance, such investigations remain scarce in the Botryosphaeriaceae, partly due to difficulties in inducing sporulation under laboratory conditions.
Botryosphaeriaceae species do not regularly sporulate on conventional media such as Malt Extract Agar (MEA) or Potato Dextrose Agar (PDA). Accordingly, many culture media and incubation regimes have been tested in attempts to induce sporulation (Phillips 2016; Vieira et al. 2021). Among the most commonly used media is water agar supplemented with plant material, particularly pine needles, which stimulate pycnidial production by providing organic compounds and nutrients (Smith et al. 1996; Crous et al. 2006, 2019). In addition to pine needles, other substrates such as poplar twigs (González-Domínguez et al. 2017) and pistachio leaves (Chen et al. 2014; Agustí-Brisach et al. 2019) have also been used successfully. Although in vitro culture-based protocols are most common, alternative methods such as direct inoculation of plant tissues with mycelial plugs, e.g. green grapevine shoots, have also proven effective (Amponsah et al. 2011). The success and number of spores obtained using these methods are variable, possibly due to the inherently variable nature of host material derived from different species and ecological contexts.
A significant recent breakthrough in sporulation of the Botryosphaeriaceae was reported by Oostlander et al. (2023), who demonstrated that D. sapinea readily sporulates on Vogel’s Minimal Medium (VMM), producing large numbers of pycnidia within three weeks of incubation at 25 °C under constant white light. The medium was originally developed by Vogel (1956) for the growth of Neurospora crassa and is commonly used to induce its asexual sporulation (Greenwald et al. 2010). Oostlander et al. (2023) successfully induced the production of 1.7 × 10^6^ conidia mL^-1^ per Petri dish after 21 days of incubation. These conidia could be used for inoculation of seedlings, as well as for transformation. This technique not only regularly produces large numbers of spores but also provides a more standardised medium for comparing experiments. More recently, Slippers et al. (2024) demonstrated that the same method was also effective for Oblongocollomyces ednahkunjekuae.
In this study, we aimed to test the protocol developed by Oostlander et al. (2023) on three widespread genera within the Botryosphaeriaceae, namely Diplodia, Lasiodiplodia, and Neofusicoccum, which are important pathogens of woody plants worldwide (Crous et al. 2006; Batista et al. 2021; Garcia et al. 2021). We evaluated the ability of VMM to induce sporulation in three species of Diplodia, two species of Lasiodiplodia, and 26 species of Neofusicoccum. Additionally, we evaluated different sucrose concentrations in the VMM formulation to determine optimal conditions for sporulation among species and isolates, and we assessed whether long-term storage on Malt Yeast Agar medium influenced sporulation of Neofusicoccum spp. on this medium.
Methods
Fungal isolates and identification
The efficacy of VMM in supporting sporulation of Botryosphaeriaceae species was evaluated using 123 isolates from the fungal collection of the Instituto Canario de Investigaciones Agrarias (Canary Islands, Spain). All strains were obtained between 2021 and 2024 from a range of hosts, including Delonix regia, Dracaena draco, Ficus macrocarpa, Persea americana Mill., Phoenix canariensis, Prunus domestica, Pinus radiata, Rosa sp., Schinus molle, Schotia brachypetala, and Vitis vinifera, and some have been previously reported by Hernández et al. (2023) (Suppl. material 1: table SS1). Taxonomic authorities for the taxa identified in this study are provided at first mention in the Results. Species-level identification of all isolates was based on sequencing four loci: the internal transcribed spacer regions 1 and 2, including the intervening 5.8S nrDNA gene (ITS), the translation elongation factor 1-alpha gene (tef1-α), the beta-tubulin gene (tub2), and the DNA-directed RNA polymerase II second-largest subunit gene (rpb2). Primers used were ITS1/ITS4 (White et al. 1990), EF1-728F/EF1-986R (Carbone and Kohn 1999), Bt2a/Bt2b (Glass and Donaldson 1995), RPB2-LasF/RPB2-LasR (Cruywagen et al. 2017) for Lasiodiplodia species, and RPB2bot6F/RPB2bot7R (Sakalidis et al. 2011) for Neofusicoccum species. PCR amplification protocols followed those described by Pavlic et al. (2007), Phillips et al. (2013), Slippers et al. (2013), and Cruywagen et al. (2017). Maximum likelihood and Bayesian phylogenetic analyses were performed using RAxML-HPC2 v.8.2.12 (Stamatakis 2014) and MrBayes v.3.2.7 (Ronquist et al. 2012), respectively, and were executed via the CIPRES Science Gateway (Miller et al. 2010) using representative and type sequences from all species of the Diplodia, Lasiodiplodia, and Neofusicoccum genera, as described in Hernández et al. (2023).
Evaluation of VMM for stimulating pycnidial and conidial formation
All strains were initially grown on 2% Malt Extract Agar (MEA; Condalab #1708.25) plates for one week at 22 ± 1 °C. Subsequently, mycelial plugs were transferred to VMM plates (two plates per isolate) prepared according to Oostlander et al. (2023), with the agar concentration increased to 2%. Plates were sealed with Parafilm® and incubated under constant white light (Sylvania, F36W/T8/ACTIVA, 0002219) at 25 °C. The distance between the light source and the surface of the VMM plates was 52 cm. All plates were positioned with the mycelial surface facing upwards to ensure direct light exposure. Initial pycnidia formation and first conidial discharge were recorded daily. Plates were maintained in incubation until peak sporulation was reached for each isolate, for a maximum of 35 days. Upon observing conidial release, primarily in cirrus form, strains producing abundant and homogeneous cirri across the entire plate, in most species, were used to determine the conidial concentration produced under the specified incubation conditions. The incubation time required for spore release to allow conidial harvesting was also recorded. The protocol developed by Oostlander et al. (2023) for harvesting conidia was followed with the following modifications: instead of pipetting 2 mL of Tween 20 solution (0.01% v/v in distilled water) onto the surface of each culture, the solution was added until the entire surface of the plate was covered, ensuring that all pycnidia and cirri were submerged. Each plate was incubated at 22 ± 1 °C for at least 30 minutes, with gentle shaking every 10 minutes to dislodge conidia from the pycnidia. Plates were then rinsed several times with the Tween solution, and the suspension was transferred to a 15 mL tube. Tubes were centrifuged for 15 minutes at 4700 rpm to obtain a conidial pellet. The Tween solution was removed, and the pellet was resuspended in 30% glycerol and transferred to a new 1.5 mL tube. The concentration of each conidial suspension was measured twice using a Neubauer-improved counting chamber. The resulting suspensions were stored at −80 °C, and spore viability was confirmed one month later by plating an aliquot on MEA to verify spore germination and subsequent mycelial growth.
Effect of sucrose concentration on pycnidial development and conidial release
In certain species, such as N. parvum, pycnidia may develop beneath dense aerial mycelium or become embedded within the culture medium, which can hinder efficient conidial harvesting. In other cases, pycnidia formation is restricted to the oldest area of the culture or to the edges of the colony on the Petri dish. To determine whether these features could be modified, the concentration of sucrose in VMM was adjusted. The effect of sucrose concentration on sporulation was evaluated over a 7-week period for Neofusicoccum species, 3 weeks for Lasiodiplodia species, and 2–5 weeks for Diplodia species, reflecting the time required for isolates of each genus to reach peak sporulation. Variation in assay duration among species was based on weekly macroscopic evaluations of each culture, which were used to determine the point at which each isolate reached peak sporulation. The recommended sucrose concentration in VMM (1.5% w/v) was adjusted to 1%, 0.5%, 0.25%, and 0%. A total of 18 representative isolates were selected for this assay: D. africana (n = 2), D. mutila (n = 1), D. seriata (n = 2), Lasiodiplodia brasiliensis (n = 2), L. theobromae (n = 1), N. australe (n = 1), N. cryptoaustrale (n = 1), N. luteum (n = 2), N. parvum (n = 4), and N. stellenboschiana (n = 2) (Suppl. material 1: table SS1). All cultures were incubated under the conditions described above. The onset of pycnidial formation and first conidial discharge were recorded daily, together with the incubation time required to achieve sufficient sporulation for conidial harvesting and concentration measurements.
Sporulation of Neofusicoccum strains in long-term storage
A total of 67 strains representing 26 Neofusicoccum species were selected from the Culture Collection of the Forestry and Agricultural Biotechnology Institute (FABI) (CMW) at the University of Pretoria, South Africa. The cultures were stored on Malt Yeast Agar (MYA; 3 g malt extract, 3 g yeast extract, 10 g glucose, 5 g peptone, and 25 g agar) slants at 4 °C. This assay aimed to determine whether the findings on pycnidia formation and conidial discharge in recently isolated Neofusicoccum strains from the Canary Islands, as described above, were applicable to a broader range of strains, species, hosts, and geographic origins, and to assess the potential impact of long-term storage on their ability to sporulate on VMM. Strains were initially transferred from the collection to MEA plates and incubated at 22 ± 1 °C for one week. These MEA plates were retained for 2 weeks after transfer to VMM to confirm pycnidia formation or conidial exudation. Subsequently, each strain was subcultured onto two VMM plates and incubated under the same conditions described above. Initial pycnidia formation and first conidial discharge were recorded daily, along with the incubation time required for conidial harvest and the concentration of conidia obtained, following the same protocol described previously. In addition, genomic DNA was extracted from all isolates, and the ITS region was amplified using primers ITS1 and ITS4, as described above. The obtained amplicons were sequenced at the University of Pretoria Sequencing Facility to confirm that all isolates were uncontaminated and belonged to the genus Neofusicoccum.
Results
Fungal isolates and identification
The phylogenetic analyses based on the ITS, tef1-α, tub2, and rpb2 loci enabled the identification of all 123 isolates at the species level (Suppl. material 1: tables S1, S3–S5, figs 1–3). Isolates were assigned to ten Botryosphaeriaceae species: Diplodia africana Damm & Crous (n = 12), D. mutila (Fr.) Fr. (n = 1), D. seriata De Not. (n = 2), Lasiodiplodia brasiliensis M.S.B. Netto, M.W. Marques & A.J.L. Phillips (n = 11), L. theobromae Griffon & Maublanc (n = 1), Neofusicoccum australe (Slippers, Crous & M.J. Wingfield) Crous, Slippers & A.J.L. Phillips (n = 1), N. cryptoaustrale Pavlic, Maleme, Slippers & M.J. Wingfield (n = 1), N. luteum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips (n = 23), N. parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips (n = 37), and N. stellenboschiana Tao Yang & Crous (n = 34). Forty-seven of these isolates had been preliminarily identified and reported by Hernández et al. (2023), while the remaining 76 isolates were identified for the first time in this study.
Evaluation of VMM for stimulating pycnidial and conidial formation
To assess the capacity of VMM to promote sporulation, 123 isolates were initially evaluated (Fig. 1). Of these, 50 isolates that produced abundant and homogeneous cirri across the entire plate were selected to determine the conidial concentration that each species could produce under the specified incubation conditions (Suppl. material 1: table SS2). The results obtained with the selected isolates of Diplodia (Fig. 2), Lasiodiplodia (Fig. 3), and Neofusicoccum (Fig. 4) confirmed the effectiveness of VMM in inducing pycnidia formation across a broad range of species within these genera (Table 1). All isolates of D. mutila, D. seriata, L. theobromae, N. australe, N. cryptoaustrale, and N. luteum produced pycnidia. Additionally, pycnidia were observed in 91.7% of D. africana, 81.8% of L. brasiliensis, 83.8% of N. parvum, and 97.1% of N. stellenboschiana isolates. The mean time to pycnidia formation varied among species, occurring at 3 days for D. seriata; 4 days for N. cryptoaustrale and N. luteum; 5 days for L. brasiliensis, L. theobromae, N. australe, N. parvum, and N. stellenboschiana; and 14 days for D. mutila and D. africana.
Harvesting conidia from Lasiodiplodia brasiliensis (D106 = CMW 67234) incubated on VMM for 21 days. A, B Top and bottom views of the culture; C Culture immersed in a solution of 0.01% (v/v) Tween 20; D Conidial suspension recovered from the culture; E Conidia pellet after centrifugation; F Quantification of conidia under a light microscope. Scale bar: 20 μm.
Sporulation of Diplodia species. A–DD. africana (D242 = CMW 67242) after 21 days of incubation; E–HD. mutila (D120 = CMW 67239) after 21 days of incubation; I–LD. seriata (13.1.1 = CMW 67254) after 14 days of incubation. A, E, I Top view of the culture; B, F, J Bottom view of the culture; C, D Pycnidia; G, H, K Pycnidia releasing conidia in the form of cirri; L Cirrus released from a pycnidium. Scale bars: 0.4 mm (C–D); 0.5 mm (G–H); 1 mm (K); 0.1 mm (L).
Sporulation of Lasiodiplodia species after 14 days of incubation. A–DL. brasiliensis (D106 = CMW 67234); E–HL. theobromae (D113 = CMW 67235). A, E Top view of the culture; B, F Bottom view of the culture; C–G Pycnidia releasing conidia in the form of cirri; D, H Quantification of conidia under a light microscope. Scale bars: 1 mm (C, G); 20 μm (D, H).
Sporulation of Neofusicoccum species. A–DN. australe (D31 = B018 = CMW 64262) after 28 days of incubation; E–HN. cryptoaustrale (D39 = B043 = CMW 64270) after 35 days of incubation; I–LN. luteum (D89 = CMW 64314) after 21 days of incubation; M–PN. parvum (D50 = B110 = CMW 64281) after 28 days of incubation; Q–TN. stellenboschiana (D43 = B050 = CMW 64274) after 28 days of incubation. A, E, I, M, Q Top view of the culture; B, F, J, N, R Bottom view of the culture; C, K, O, S Pycnidia releasing conidia in the form of cirrus; D, H, L, P, T Conidia; G Pycnidia. Scale bars: 1 mm (C, G); 20 μm (D, H, L, P, T); 0.5 mm (K, O, S).
All isolates of D. mutila, D. seriata, L. theobromae, N. australe, N. cryptoaustrale, and N. luteum released conidia in the form of cirri. However, not all isolates of D. africana, L. brasiliensis, N. parvum, and N. stellenboschiana were able to release conidia from the pycnidia, and those isolates that sporulated did so by releasing their conidia in the form of cirri (Table 1). Only 41.7% of D. africana isolates released conidia, while 81.8%, 59.5%, and 76.5% of L. brasiliensis, N. parvum, and N. stellenboschiana isolates were able to do so, respectively. Species such as D. seriata, L. brasiliensis, L. theobromae, and N. luteum produced the most abundant and homogeneous cirri across the entire culture. The time required to observe the first conidial discharge differed among species. The first species to release conidia after being subcultured onto VMM were L. brasiliensis and L. theobromae, followed by D. seriata and N. luteum. Subsequently, N. australe started to release conidia, whereas D. africana, D. mutila, N. parvum, N. stellenboschiana, and N. cryptoaustrale were the last to do so.
The decision on the optimal time for harvesting conidia, from the day of subculturing onto VMM, was based on the cessation of new pycnidia and cirri formation. Each culture was monitored daily until no further pycnidia or cirri were produced. Consequently, the optimal time for conidia harvesting ranged from 14 days for the earliest species to 35 days for the slowest (Table 1). D. africana, D. mutila, N. cryptoaustrale, N. parvum, and N. stellenboschiana required the longest time for conidia collection and also showed the greatest delay in initiating conidial release from pycnidia. Species such as N. parvum and N. stellenboschiana exhibited prominent mycelial development, particularly N. parvum, which concealed and delayed the sporulation process, resulting in a delay in conidia harvesting. In the case of D. africana, pycnidia were produced exclusively in the oldest area of the culture, around the 5-mm mycelial plug placed onto the VMM plate.
With regard to the mean number of conidia yielded (Table 1), N. luteum generated the highest concentration, followed by L. brasiliensis and L. theobromae. Although N. parvum and N. stellenboschiana required the longest time for conidia harvest, these species yielded concentrations of up to 2 × 10^6^ conidia mL^-1^. The only Diplodia species for which conidial concentration could be quantified was D. seriata. The lowest conidia production was recorded for N. australe and N. cryptoaustrale, with less than 4 × 10^5^ conidia mL^-1^ for both species. Although pycnidia and conidia were macroscopically observed in D. africana and D. mutila, no conidia were detected using the Neubauer-improved counting chamber.
Effect of sucrose concentration on pycnidia development and conidia release
Daily macroscopic evaluation of all species (Figs 5, 6) revealed that, although all tested species developed pycnidia under most conditions, their ability to release conidia varied significantly, depending on the species and sucrose availability. In general, higher or intermediate sucrose concentrations (particularly 1.5% and 1%) promoted earlier development and greater sporulation in most species, whereas low or absent sucrose levels often delayed pycnidia formation and inhibited or entirely suppressed conidial release, especially in species such as D. africana, D. mutila, and N. cryptoaustrale (Table 2).
Colony morphology of Diplodia and Lasiodiplodia species used in the sucrose-concentration assay on pycnidia development and conidial release. Each photograph was taken prior to conidial harvesting. See Table 2 for harvest dates.
Colony morphology of Neofusicoccum species used in the sucrose-concentration assay on pycnidia development and conidial release. Each photograph was taken prior to conidial harvesting. See Table 2 for harvest dates.
Diplodia africana formed pycnidia at all sucrose levels, with conidia release occurring only at 1.5%, 1%, and 0%, although conidia were only quantified at 0% sucrose. In D. mutila, despite visible pycnidia development across treatments, no conidia were quantified, indicating high sensitivity to environmental or nutritional constraints. In contrast, D. seriata consistently released conidia under all sucrose-supplemented conditions, with optimal yields at 0.5% and 0.25%. Similarly, L. brasiliensis and L. theobromae responded favourably to intermediate concentrations, particularly 1%, which supported both early development and maximal conidia production, whereas 0% sucrose resulted in the lowest sporulation. Among Neofusicoccum spp., a clear preference for 1–1.5% sucrose was evident. For example, N. australe reached peak conidial production at 1%, whereas N. luteum, N. stellenboschiana, and N. parvum exhibited earlier and more prolific sporulation at 1.5%. Notably, N. cryptoaustrale displayed low productivity under all conditions, suggesting intrinsic physiological limitations under the experimental conditions (Table 3). Across all species, sucrose deprivation (0%) consistently resulted in delayed pycnidia development and minimal or absent conidial release.
Sporulation of Neofusicoccum strains in long-term storage
From the 26 selected Neofusicoccum species (taxonomic authorities are provided in Table 4) preserved in the CMW collection (Table 4), 20 developed pycnidia on VMM (Table 5). The species for which all isolates failed to produce pycnidia under these conditions were N. andinum, N. kwambonambiense, N. luteum, N. mangiferae, N. stellenboschiana, and N. terminaliae. The time required for first visible pycnidial formation among the remaining species ranged from 3 to 38 days. Some isolates from 12 species (N. australe, N. cryptoaustrale, N. dianense, N. eucalypticola, N. eucalyptorum, N. lumnitzerae, N. microconidium, N. ningerense, N. occulatum, N. pandanicola, N. variabile, and N. vitifusiforme) discharged conidia, producing visible cirri from pycnidia. The incubation time required for the first conidial discharge ranged from 18 to 32 days.
Among all Neofusicoccum species evaluated (Table 5), the highest conidial concentration was recorded for N. eucalypticola isolate CMW 6539, yielding 1.24 × 10^7^ conidia mL^-1^. In contrast, isolate CMW 6543 of the same species produced substantially fewer conidia (1.27 × 10^6^ conidia mL^-1^). Neofusicoccum australe isolate CMW 9073 produced 3.62 × 10^6^ conidia mL^-1^, followed by N. eucalyptorum isolate CMW 10125 and N. lumnitzerae isolate CMW 41228, both yielding 6.75 × 10^5^ conidia mL^-1^. Neofusicoccum dianense isolates CMW 54189 and CMW 54123 produced 1.75 × 10^5^ and 150,000 conidia mL^-1^, respectively. The lowest concentrations were observed in N. ningerense (CMW 54212), N. pandanicola (CMW 14087), N. variabile (CMW 37745), and N. vitifusiforme (CMW 30605), all with 5.0 × 10^4^ conidia mL^-1^, although isolate CMW 54144 of N. ningerense produced 1.5 × 10^5^ conidia mL^-1^.
No association was detected between sporulation on VMM and the geographic origin of the isolates or their host of isolation (Fig. 7). In contrast, a relationship was observed with culture age, defined by date of isolation. Isolates obtained during the 1980s consistently failed to produce pycnidia or conidia under the tested conditions. By comparison, a few isolates from the late 1990s, such as N. australe isolates CMW 1133 (1996) and CMW 9073 (1999), formed pycnidia, although conidial production was limited. Isolates from 2000–2007 displayed greater variability, with some strains yielding high conidial concentrations (e.g., N. eucalypticola isolates CMW 6543 and CMW 6539, both isolated in 2000, producing 1.27 × 10^6^ and 1.24 × 10^7^ conidia mL^-1^, respectively), while others from the same period showed no sporulation. More recent isolates (2010–2018) generally displayed higher sporulation frequencies, exemplified by N. dianense isolates CMW 54123 and CMW 54189 (2018), producing 1.5 × 10^5^ and 1.75 × 10^5^ conidia mL^-1^ respectively, and N. ningerense isolate CMW 54144 (2018), which reached 1.5 × 10^5^ conidia mL^-1^ in some cases.
Sporulation of Neofusicoccum strains after long-term preservation and subsequent incubation on VMM for 28 days. A, B Front and reverse sides of N. australe CMW 9073; C, DN. cryptoaustrale CMW 23785; E, FN. eucalypticola CMW 6543; G, HN. eucalyptorum CMW 10125; I, JN. ningerense CMW 54212; K, LN. vitifusiforme CMW 30605.
Discussion
In this study, we extend the work of Oostlander et al. (2023) by testing and refining their sporulation protocol for D. africana, D. mutila, D. seriata, L. brasiliensis, and L. theobromae, as well as 26 species of Neofusicoccum. Our findings demonstrate that VMM, under constant white light at 25 °C, effectively promotes the formation of pycnidia and the discharge of conidia in a wide array of taxa, but with notable differences between species and isolates. Such inter- and intraspecific variation is not unexpected in the Botryosphaeriaceae and likely reflects a combination of biological and culture-related factors. Species-specific developmental regulation and nutritional requirements can influence both the timing of pycnidial initiation and the efficiency of conidial discharge, which is consistent with the contrasting sporulation responses observed across taxa under different sucrose concentrations in our assays. In addition, isolate-level variation may reflect differences in physiological status linked to culture history (e.g. number of transfers, growth morphology, and the extent of aerial mycelium), which can affect pycnidial maturation and cirri production even under otherwise standardised conditions. Moreover, culture age and long-term storage history can reduce reproductive competence, as reflected by the progressive loss of sporulation capacity observed in older strains in our work. This study therefore represents a significant advancement in the development of standardised and reproducible methodologies for inducing sporulation in members of the Botryosphaeriaceae.
The protocol described in this study using VMM demonstrated rapid, reproducible, and substrate-independent sporulation. Pycnidia development began within three days for D. seriata, four days for N. cryptoaustrale and N. luteum, and five days for L. brasiliensis, L. theobromae, N. australe, N. parvum, and N. stellenboschiana. The quantity of conidia produced exceeded those reported in previous studies. For instance, N. luteum reached a mean concentration of 1.5 × 10^7^ conidia mL^-1^, more than 600 times higher than the highest value reported by Amponsah et al. (2008). Notably, both studies identified N. luteum as the most prolific species in terms of conidia production. The VMM protocol not only excels in speed and yield but also in operational simplicity. It eliminates the need for plant tissue substrates and photoperiod control. Cultures maintained under constant white light at 25 °C showed consistent pycnidia formation in most species. Moreover, the release of conidia in cirri by the majority of species further simplified harvesting, which was achieved by flooding plates with 0.01% Tween 20, followed by gentle agitation, centrifugation, and final resuspension. Unlike traditional methods, there was no need for manual disruption of pycnidia.
When comparing our VMM-based results with those of Oostlander et al. (2023) for D. sapinea, it is notable that some species and strains using the protocol described here produced even higher sporulation levels. Oostlander et al. (2023) reported a maximum of 1.7 × 10^6^ conidia mL^-1^ per plate (5.5 cm diameter). In our assays, using two 6.0 cm plates per isolate, D. seriata produced 1.3 × 10^6^ conidia mL^-1^, comparable to the values reported by Oostlander et al. (2023). However, significantly higher yields were recorded for L. brasiliensis (8.7 × 10^6^ and 7.12 × 10^6^ conidia mL^-1^), N. parvum (2.31 × 10^6^ conidia mL^-1^), N. stellenboschiana (2.0 × 10^6^ conidia mL^-1^), and particularly N. luteum (1.5 × 10^7^ conidia mL^-1^). These data confirm the broad applicability of VMM beyond Diplodia, extending its efficacy to support spore-based studies across the Botryosphaeriaceae.
In our study, continuous white light at 25 °C favoured pycnidia formation and conidia discharge. Light conditions have long been recognised as influential in fungal development (Smith and Fergus 1971). Although previous research, such as Úrbez-Torres and Gubler (2009), demonstrated that alternating light and dark periods promoted sporulation in some species, the constant white light conditions initially established by Oostlander et al. (2023) proved effective for most taxa tested here. Nevertheless, future studies should evaluate whether varying light regimes (e.g. white light-dark cycles, variation in light wavelength, or near-UV exposure) could further enhance sporulation outcomes across additional species or isolates using VMM.
Sucrose concentration in VMM had a substantial impact on pycnidia development and conidia discharge, underscoring the critical role of carbon availability in regulating sporulation efficiency in the Botryosphaeriaceae. Although all tested species formed pycnidia across a range of sucrose levels, both the timing and quantity of conidia released varied considerably. Optimal sporulation was observed at sucrose concentrations of 1.5%–1% for L. brasiliensis, L. theobromae, N. luteum, N. parvum, and N. stellenboschiana, suggesting that balanced carbon availability supports not only structural development but also the metabolic processes required for active conidia release. In contrast, low or absent sucrose levels delayed pycnidia formation and complicated sporulation in species such as D. mutila and N. cryptoaustrale. Notably, D. africana formed pycnidia at all sucrose concentrations, and conidial discharge occurred at 1.5%, 1%, and 0%; however, conidia were microscopically observed only at 0% sucrose, indicating delayed and markedly reduced sporulation compared with other species. In contrast, D. seriata showed optimal sporulation at 0.5% and 0.25% sucrose. These patterns indicate that sucrose functions not only as a nutrient but also potentially as a regulatory cue in reproductive development. Precise adjustment of sucrose concentration is therefore essential for optimising sporulation protocols.
Geographic origin and host species did not influence sporulation capacity in this study, whereas a progressive loss of sporulation capacity was associated with increasing culture age. Strains collected from Australia, Indonesia, Spain, and South Africa all demonstrated the ability to produce pycnidia and conidia under standardised conditions. Isolates obtained by the Instituto Canario de Investigaciones Agrarias (Spain) between 2021 and 2024 showed robust sporulation, whereas older strains from the CMW collection (isolated between 1986 and 2018 and maintained on MYA at 4 °C since isolation) exhibited reduced capacity, with some failing to sporulate entirely. This observation is consistent with previous findings by Baskarathevan et al. (2009) and Tariq et al. (2015), suggesting that long-term storage or repeated subculturing may compromise reproductive viability. Nonetheless, the presence of older isolates with high conidial yields (e.g. N. australe CMW 9073, producing 3.62 × 10^6^ conidia mL^-1^ after storage since 1999) indicates that additional factors such as species identity, storage conditions, or the number of prior subcultures also influence sporulation capacity. Isolates and conidial suspensions obtained using this protocol can be cryopreserved in 40% glycerol at −80 °C, as described by Oostlander et al. (2024). Further studies are required to determine optimal preservation conditions for individual species and to assess whether cryopreservation under these or alternative conditions affects strain viability and subsequent sporulation.
Species-level sporulation patterns revealed marked differences across genera. Diplodia seriata consistently exhibited the fastest and most abundant sporulation across several sucrose concentrations, whereas D. africana and D. mutila showed poor or no conidial discharge, suggesting species-specific limitations or unidentified sporulation triggers. Lasiodiplodia brasiliensis and L. theobromae sporulated efficiently under all conditions tested. In Neofusicoccum, sporulation behaviour varied widely both among species and among isolates. Neofusicoccum luteum was the most efficient species at a sucrose concentration of 1.5%. In contrast, N. australe and N. cryptoaustrale sporulated better at lower sucrose concentrations, whereas N. parvum and N. stellenboschiana showed delayed and inconsistent sporulation, with lower conidial yields and some isolates failing to form pycnidia.
Several media have previously been used to induce sporulation in Botryosphaeriaceae species, often supplemented with plant material. These include 2% MEA, 2% PDA, Oatmeal Agar (OA) (Crous et al. 2019), 2% water agar (Phillips et al. 2013), and Prune Agar (PA) (Amponsah et al. 2008). A limitation of these media is their fixed composition, which restricts precise manipulation of individual components for investigating the influence of specific chemical variables on sporulation. In contrast, VMM allows precise modification of each constituent component, enabling the formulation to be tailored to the physiological requirements of target species. Among its components, sucrose serves as a key carbon source, and its concentration was shown here to strongly influence sporulation efficiency. Oostlander et al. (2023) conducted a side-by-side evaluation of several commonly used culture media (CD, LNA, OA, and PDA) alongside VMM under controlled incubation conditions and observed pycnidia formation and harvestable spores on VMM (and VMM supplemented with pine needles), but not on CD, LNA, OA, or PDA for D. sapinea. Based on this evidence, and because the primary objective of the present study was to establish a single standardised protocol applicable across a large and taxonomically diverse isolate set, we focused on VMM and optimisation of its key parameters rather than including parallel medium controls for each isolate. Nevertheless, systematic cross-medium comparisons across multiple taxa remain an important direction for future research.
Plant materials, particularly pine needles but also twigs from various woody plants, are commonly used as substrates to stimulate sporulation in the Botryosphaeriaceae. Similar approaches are applied to other fungal taxa, such as leaves of Rhododendron pulchrum for Guignardia endophyllicola (Phyllosticta capitalensis) (Okane et al. 2001), Dianthus caryophyllus leaves for Fusarium species (Fisher et al. 1982) and Pestalotiopsis species (Liu et al. 2010), or Gardenia jasminoides and Hydrangea macrophylla leaves to induce perithecia and ascospores in Glomerella singulata, Mycosphaerella allicina, Metasphaeria sp., Guignardia sp., and Pleospora herbarum (Furukawa and Kishi 2002). However, plant materials are not always readily available, and their chemical composition can vary depending on host species, geographic origin, and physiological state. This variability introduces inconsistency into sporulation studies, whereas the use of VMM provides compositional consistency that ensures reproducibility across experiments and laboratories. Moreover, while plant substrates restrict pycnidia development to their surfaces, VMM enables pycnidia formation across the entire Petri dish.
Conclusion
Collectively, these findings demonstrate that VMM offers a defined, reproducible, and technically simple framework for inducing sporulation in the Botryosphaeriaceae under the incubation conditions tested here. Its defined composition, high reproducibility, technical simplicity, and substantial conidial yields make it a valuable and versatile tool for both mycological and plant pathology research. In a fungal family characterised by pronounced interspecific and intraspecific variability in sporulation behaviour, the establishment of a medium applicable to species from three genera represents a significant methodological advancement. This study provides a robust quantitative and procedural framework that bridges laboratory experimentation and ecological or applied contexts, supporting cross-laboratory reproducibility and enabling more controlled studies. However, further research is required, including systematic cross-medium comparisons under identical incubation conditions, to optimise VMM formulations for specific taxa, uncover the molecular and physiological mechanisms underpinning sporulation, and integrate morphological and molecular data to enhance taxonomic resolution.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agustí-Brisach C, Moral J, Felts D et al. (2019) Interaction between Diaporthe rhusicola and Neofusicoccum mediterraneum causing branch dieback and fruit blight of English Walnut in California, and the effect of pruning wounds on the infection. Plant Disease 103(6): 1196–1205. 10.1094/PDIS-07-18-1118-RE 30958106 · doi ↗ · pubmed ↗
- 2Ahimera N, Gisler S, Morgan DP et al. (2004) Effects of single-drop impactions and natural and simulated rains on the dispersal of Botryosphaeria dothidea conidia. Phytopathology 94(11): 1189–1197. 10.1094/PHYTO.2004.94.11.118918944454 · doi ↗ · pubmed ↗
- 3Almeida MC, Antunes D, Silva BMA et al. (2019) Early interaction of Alternaria infectoria conidia with macrophages. Mycopathologia 184: 383–392. 10.1007/s 11046-019-00339-631183740 · doi ↗ · pubmed ↗
- 4Amponsah NT, Jones EE, Ridgway HJ et al. (2008) Production of Botryosphaeria species conidia using grapevine green shoots. New Zealand Plant Protection 61: 301–305. 10.30843/nzpp.2008.61.6840 · doi ↗
- 5Amponsah NT, Jones EE, Ridgway HJ et al. (2009) Rainwater dispersal of Botryosphaeria conidia from infected grapevines. New Zealand Plant Protection 62: 228–233. 10.30843/nzpp.2009.62.4824 · doi ↗
- 6Amponsah NT, Jones EE, Ridgway HJ et al. (2011) Identification, potential inoculum sources and pathogenicity of Botryosphaeriaceous species associated with grapevine dieback disease in New Zealand. European Journal of Plant Pathology 131: 467–482. 10.1007/s 10658-011-9823-1 · doi ↗
- 7Arya GC, Sarkar S, Manasherova E et al. (2021) The plant cuticle: An ancient guardian barrier set against long-standing rivals. Frontiers in Plant Science 12: 663165. 10.3389/fpls.2021.663165 PMC 826741634249035 · doi ↗ · pubmed ↗
- 8Baskarathevan J, Jaspers MV, Jones EE et al. (2009) Evaluation of different storage methods for rapid and cost-effective preservation of Botryosphaeria species. New Zealand Plant Protection 62: 234–237. 10.30843/nzpp.2009.62.4825 · doi ↗