Funiliomycetaceae fam. nov. (Amphisphaeriales, Ascomycota) accommodating Funiliomyces, including F. jiangxiensis sp. nov. from Tetradium ruticarpum and ten new combinations

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
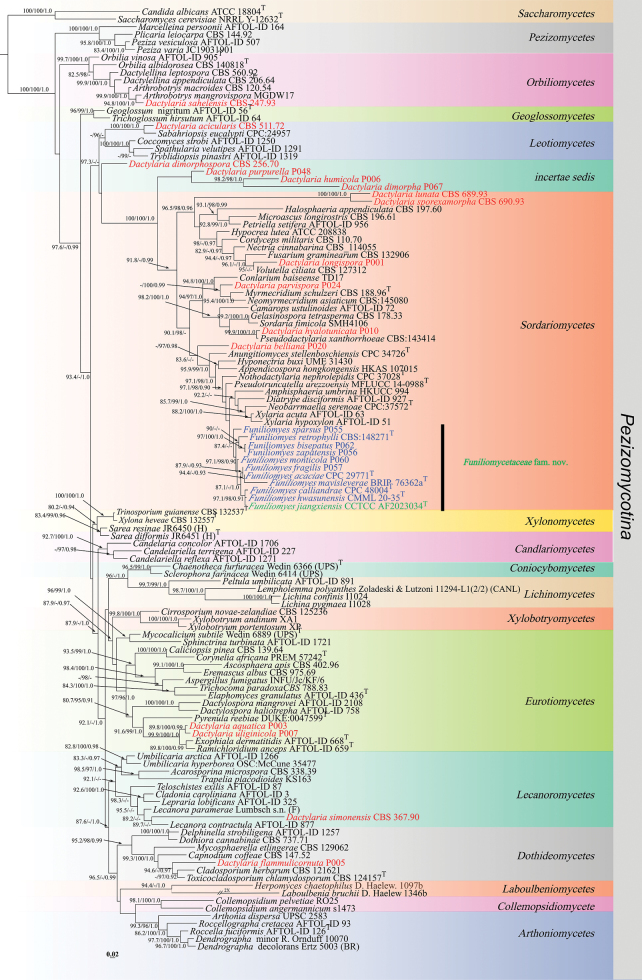 Figure 1
Figure 1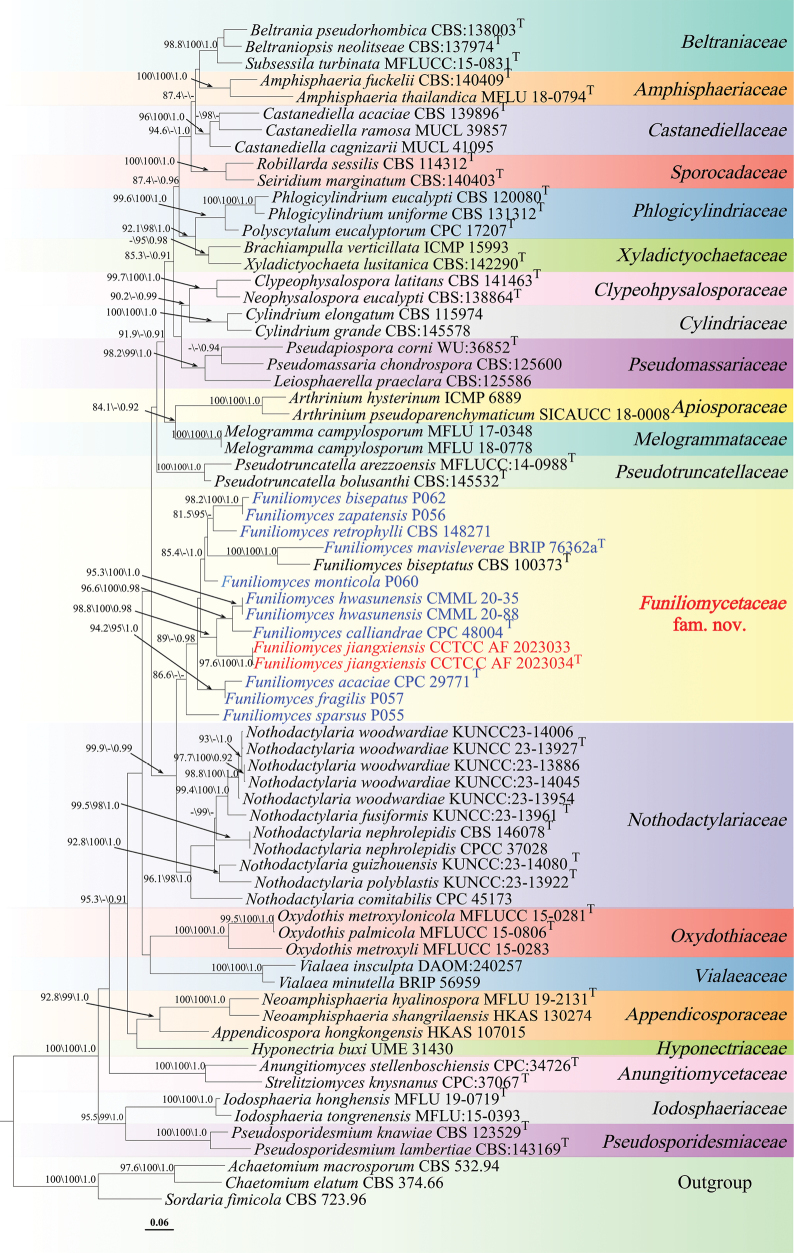 Figure 2
Figure 2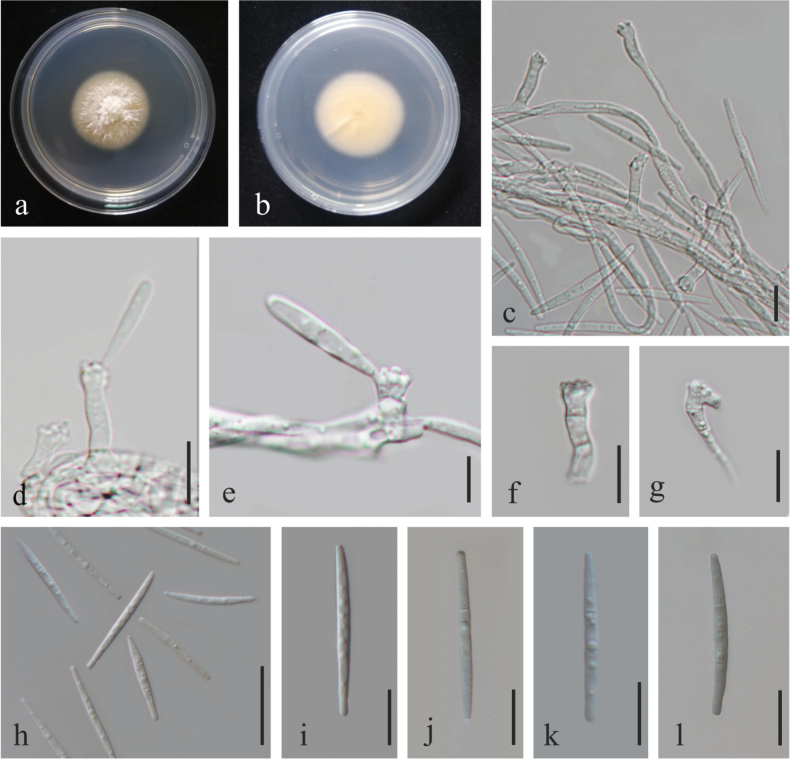 Figure 3
Figure 3| Taxon | Strain | GenBank accession numbers | References |
|---|---|---|---|
| LSU | |||
|
| CBS 338.39 |
|
|
|
| HKUCC 994 |
|
|
|
| CPC 34726T |
|
|
|
|
|
| |
|
| UPSC 2583 |
|
|
|
| CBS 120.54 |
|
|
|
| MGDW17 |
|
|
|
| CBS 402.96 |
|
|
|
| INFU/Jc/KF/6 |
|
|
|
| CPC:37572T |
|
|
|
| CBS 139.64 |
|
|
|
| AFTOL-ID 72 |
|
|
|
| AFTOL-ID 1706 |
|
|
|
| AFTOL-ID 1271 |
|
|
|
| AFTOL-ID 227 |
|
|
|
| ATCC 18804T |
|
|
|
| CBS 147.52 |
|
|
|
| Wedin 6366 (UPS)T |
|
|
|
| CBS 125236 |
| Réblová et al. (2012) |
|
| AFTOL-ID 3 |
|
|
|
| CBS 121621 |
|
|
|
| AFTOL-ID 1250 |
|
|
|
| s1473 |
|
|
|
| RO25 |
|
|
|
| TD17 |
|
|
|
| CBS 110.70 |
|
|
|
| PREM 57242T |
|
|
|
| CBS 511.72 |
|
|
|
| P003 |
| Bhilabutra et al. (2007, submitted directly) |
|
| P020 |
| Bhilabutra et al. (2007, submitted directly) |
|
| P067 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CBS 256.70 |
|
|
|
| P005 |
| Bhilabutra et al. (2007, submitted directly) |
|
| P006 |
| Bhilabutra et al. (2007, submitted directly) |
|
| P010 |
| Bhilabutra et al. (2007, submitted directly) |
|
| P001 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CBS 689.93 |
|
|
|
| P024 |
| Bhilabutra et al. (2007, submitted directly) |
|
| P048 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CBS 247.93 |
|
|
|
| CBS 367.90 |
|
|
|
| CBS 690.93 |
|
|
|
| P007 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CBS 206.64T |
|
|
|
| CBS 560.92 |
|
|
|
| AFTOL-ID 758 |
|
|
|
| AFTOL-ID 2108 |
|
|
|
| AFTOL-ID 1257 |
|
|
|
| R. Ornduff 10070 |
|
|
|
| AFTOL-ID 927 |
|
|
|
| CBS 737.71 |
|
|
|
| AFTOL-ID 436T |
|
|
|
| CBS 975.69 |
|
|
|
| AFTOL-ID 668T |
|
|
|
| CPC 29771T |
|
|
|
| P062 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CPC 48004T |
|
|
|
| P057 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CMML 20-35 |
|
|
|
|
| This study | |
|
| BRIP 76362aT |
|
|
|
| P060 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CBS:148271 |
|
|
|
| P055 |
| Bhilabutra et al. (2007, submitted directly) |
|
| P056 |
| Bhilabutra et al. (2007, submitted directly) |
|
| CBS 132906 |
|
|
|
| CBS 178.33 |
|
|
|
| AFTOL-ID 56T |
|
|
|
| CBS 197.60 |
|
|
|
| D. Haelew. 1097b |
|
|
|
| ATCC 208838 |
|
|
|
| UME 31430 |
|
|
|
| D. Haelew 1346b |
|
|
|
| AFTOL-ID 877 |
|
|
|
| Lumbsch s.n. (F) |
|
|
|
| Zoladeski and Lutzoni 11294-L1(2/2) (CANL) |
|
|
|
| AFTOL-ID 325 |
|
|
|
| I1024 |
| Garrido-Benavent (2020, submitted directly) |
|
| I1028 |
| Garrido-Benavent (2020, submitted directly) |
|
| AFTOL-ID 164 |
|
|
|
| CBS 196.61 |
|
|
|
| Wedin 6889 (UPS)T |
|
|
|
| CBS 129062 |
|
|
|
| CBS 188.96T |
| Arzanlo et al. (2007) |
|
| CBS_114055 |
|
|
|
| CBS:145080 |
|
|
|
| CPC 37028T |
|
|
|
| CBS 140818T |
|
|
|
| AFTOL-ID 905T |
|
|
|
| AFTOL-ID 891 |
|
|
|
| AFTOL-ID 956 |
|
|
|
| JC19031901 |
|
|
|
| AFTOL-ID 507 |
|
|
|
| CBS 144.92 |
|
|
|
| CBS:143414 |
|
|
|
| MFLUCC 14-0988T |
|
|
|
| DUKE:0047599T |
|
|
|
| AFTOL-ID 659T |
|
|
|
| AFTOL-ID 126T |
|
|
|
| AFTOL-ID 93 |
|
|
|
| CPC:24957 |
|
|
|
| NRRL Y-12632T |
|
|
|
| JR6451T |
|
|
|
| JR6450 |
|
|
|
| Ertz 5003 (BR) |
|
|
|
| Wedin 6414 (UPS) |
|
|
|
| SMH4106 |
|
|
|
| AFTOL-ID 1291 |
|
|
|
| AFTOL-ID 1721 |
|
|
|
| AFTOL-ID 87 |
|
|
|
| CBS 124157T |
|
|
|
| KS163 |
|
|
|
| CBS 788.83 |
|
|
|
| AFTOL-ID 64 |
|
|
|
| CBS 132537T |
|
|
|
| AFTOL-ID 1319 |
|
|
|
| AFTOL-ID 1266 |
|
|
|
| OSC:McCune 35477 |
|
|
|
| CBS 127312 |
|
|
|
| AFTOL-ID 63 |
|
|
|
| AFTOL-ID 51 |
|
|
|
| XA1 |
|
|
|
| XP |
|
|
|
| CBS 132557T |
|
|
| Taxon | Strain | GenBank accession numbers | References | ||
|---|---|---|---|---|---|
| LSU | ITS | RPB2 | |||
|
| CBS 532.94 |
|
|
|
|
|
| CBS:140409T |
|
|
|
|
|
| MFLU 18-0794T |
|
|
|
|
|
| CPC:34726T |
|
| NA |
|
|
|
|
|
|
| |
|
| ICMP 6889 |
|
|
|
|
|
| SICAUCC 18-0008 |
|
|
|
|
|
| CBS:138003T |
|
|
|
|
|
| CBS:137974T |
|
| NA |
|
|
| ICMP 15993 |
|
| NA |
|
|
| CBS 139896T |
|
| NA |
|
|
| MUCL 41095 |
|
| NA |
|
|
| MUCL 39857 |
|
| NA |
|
|
| CBS 374.66 |
|
|
|
|
|
| CBS 141463T |
|
| NA |
|
|
| CBS 115974 |
|
|
|
|
|
| CBS:145578 |
|
|
|
|
|
| CPC 29771 |
|
| NA |
|
|
| P062 |
| NA | NA | Bhilabutra et al. (2007, submitted directly) |
|
| CBS 100373T |
|
| NA |
|
|
| CPC 48004 |
|
|
|
|
|
| P057 |
| NA | NA | Bhilabutra et al. (2007, submitted directly) |
|
| CMML 20-35 |
|
| NA |
|
|
| CMML 20-88 |
| NA | NA |
|
|
|
|
|
| This study | |
|
|
|
|
| This study | |
|
| BRIP 76362a |
|
| NA |
|
|
| P060 |
| NA | NA | Bhilabutra et al. (2007, submitted directly) |
|
| CBS:148271 |
|
| NA |
|
|
| P055 |
| NA | NA | Bhilabutra et al. (2007, submitted directly) |
|
| P056 |
| NA | NA | Bhilabutra et al. (2007, submitted directly) |
|
| UME 31430 |
| NA | NA | Smith et al. (2002, submitted directly) |
|
| MFLU 19-0719T |
|
|
|
|
|
| MFLU:15-0393 |
|
| NA |
|
|
| CBS:125586 |
|
| NA |
|
|
| MFLU 17-0348 |
|
|
|
|
|
| MFLU 18-0778 |
|
|
|
|
|
| MFLU 19-2131T |
|
|
|
|
|
|
|
| NA |
| |
|
| CBS:138864T |
|
| NA |
|
|
| CPC 45173 |
|
| NA |
|
|
| KUNCC:23-13961T |
|
|
|
|
|
| KUNCC:23-14080T |
|
|
|
|
|
| CPC:37028 |
|
|
|
|
|
| CBS:146078T |
|
| NA |
|
|
| KUNCC:23-13922T |
|
|
|
|
|
| KUNCC:23-13927T |
|
|
|
|
|
| KUNCC:23-13886 |
|
|
|
|
|
| KUNCC:23-14045 |
|
|
|
|
|
| KUNCC:23-13954 |
|
| NA |
|
|
| KUNCC23-14006 |
|
| NA |
|
|
| MFLUCC15-0283 |
|
| NA |
|
|
| MFLUCC 15-0281T |
|
|
|
|
|
| MFLUCC 15-0806T |
|
|
|
|
|
| CBS 120080T |
|
|
|
|
|
| CBS 131312T |
|
| NA |
|
|
| CPC 17207T |
|
| NA |
|
|
| WU:36852T |
|
| NA |
|
|
| CBS:125600 |
|
| NA |
|
|
| CBS:123529T |
|
| NA |
|
|
| CBS:143169T |
|
| NA |
|
|
| MFLUCC:14-0988T |
|
| NA |
|
|
| CBS:145532T |
|
| NA |
|
|
| CBS 114312T |
|
| NA |
|
|
| CBS:140403T |
|
|
|
|
|
| CBS 723.96 | NA |
| NA |
|
|
| CPC:37067T |
|
|
|
|
|
| MFLUCC:15-0831T |
|
| NA |
|
|
| DAOM:240257 |
|
| NA | Hambleton et al. (2010, submitted directly) |
|
| BRIP:56959 |
|
| NA |
|
|
| CBS:142290T |
|
| NA |
|
| ITS rDNA | ||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| ||
|
| 0.110 | |||||||||||||||||||
|
| 0.181 | 0.168 | ||||||||||||||||||
|
| 0.145 | 0.157 | 0.217 | |||||||||||||||||
|
| 0.125 | 0.119 | 0.144 | 0.167 | ||||||||||||||||
|
| 0.187 | 0.185 | 0.166 | 0.209 | 0.157 | |||||||||||||||
|
| 0.150 | 0.145 | 0.185 | 0.178 | 0.145 | 0.186 | ||||||||||||||
|
| 0.137 | 0.122 | 0.189 | 0.162 | 0.128 | 0.173 | 0.136 | |||||||||||||
|
| 0.140 | 0.132 | 0.130 | 0.150 | 0.152 | 0.150 | 0.136 | 0.121 | ||||||||||||
|
| 0.240 | 0.239 | 0.228 | 0.255 | 0.225 | 0.213 | 0.238 | 0.259 | 0.242 | |||||||||||
|
| 0.250 | 0.233 | 0.237 | 0.270 | 0.223 | 0.183 | 0.223 | 0.244 | 0.195 | 0.238 | ||||||||||
|
| 0.230 | 0.229 | 0.230 | 0.264 | 0.217 | 0.182 | 0.223 | 0.229 | 0.209 | 0.231 | 0.156 | |||||||||
|
| 0.178 | 0.154 | 0.147 | 0.179 | 0.130 | 0.158 | 0.181 | 0.159 | 0.167 | 0.212 | 0.206 | 0.219 | ||||||||
|
| 0.227 | 0.214 | 0.209 | 0.255 | 0.159 | 0.179 | 0.204 | 0.202 | 0.180 | 0.240 | 0.213 | 0.221 | 0.145 | |||||||
|
| 0.286 | 0.290 | 0.308 | 0.339 | 0.270 | 0.219 | 0.292 | 0.279 | 0.229 | 0.258 | 0.262 | 0.255 | 0.223 | 0.241 | ||||||
|
| 0.235 | 0.232 | 0.278 | 0.306 | 0.256 | 0.198 | 0.269 | 0.233 | 0.196 | 0.264 | 0.273 | 0.272 | 0.204 | 0.207 | 0.253 | |||||
|
| 0.288 | 0.272 | 0.298 | 0.324 | 0.233 | 0.253 | 0.281 | 0.284 | 0.214 | 0.262 | 0.297 | 0.299 | 0.221 | 0.275 | 0.281 | 0.282 | ||||
|
| 0.310 | 0.282 | 0.293 | 0.339 | 0.276 | 0.233 | 0.293 | 0.298 | 0.245 | 0.282 | 0.286 | 0.280 | 0.218 | 0.225 | 0.300 | 0.302 | 0.297 | |||
|
| 0.316 | 0.314 | 0.308 | 0.335 | 0.302 | 0.265 | 0.292 | 0.321 | 0.280 | 0.263 | 0.257 | 0.258 | 0.298 | 0.293 | 0.297 | 0.313 | 0.318 | 0.269 | ||
|
| 0.356 | 0.329 | 0.363 | 0.369 | 0.348 | 0.275 | 0.333 | 0.318 | 0.298 | 0.270 | 0.321 | 0.306 | 0.252 | 0.271 | 0.341 | 0.330 | 0.320 | 0.316 | 0.258 | |
| LSU rDNA | ||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| 0.047 | |||||||||||||||||||
|
| 0.041 | 0.058 | ||||||||||||||||||
|
| 0.055 | 0.056 | 0.050 | |||||||||||||||||
|
| 0.049 | 0.055 | 0.040 | 0.032 | ||||||||||||||||
|
| 0.059 | 0.061 | 0.044 | 0.050 | 0.042 | |||||||||||||||
|
| 0.057 | 0.053 | 0.048 | 0.051 | 0.044 | 0.053 | ||||||||||||||
|
| 0.064 | 0.067 | 0.055 | 0.052 | 0.040 | 0.040 | 0.047 | |||||||||||||
|
| 0.060 | 0.062 | 0.051 | 0.053 | 0.039 | 0.036 | 0.053 | 0.038 | ||||||||||||
|
| 0.056 | 0.051 | 0.051 | 0.051 | 0.044 | 0.038 | 0.053 | 0.047 | 0.037 | |||||||||||
|
| 0.072 | 0.069 | 0.070 | 0.076 | 0.064 | 0.057 | 0.059 | 0.065 | 0.053 | 0.052 | ||||||||||
|
| 0.077 | 0.080 | 0.076 | 0.076 | 0.065 | 0.058 | 0.059 | 0.054 | 0.045 | 0.058 | 0.059 | |||||||||
|
| 0.061 | 0.062 | 0.062 | 0.053 | 0.058 | 0.066 | 0.058 | 0.056 | 0.070 | 0.060 | 0.080 | 0.083 | ||||||||
|
| 0.062 | 0.054 | 0.063 | 0.070 | 0.063 | 0.069 | 0.050 | 0.062 | 0.064 | 0.058 | 0.072 | 0.071 | 0.065 | |||||||
|
| 0.060 | 0.056 | 0.059 | 0.047 | 0.037 | 0.044 | 0.046 | 0.044 | 0.045 | 0.036 | 0.054 | 0.058 | 0.055 | 0.053 | ||||||
|
| 0.065 | 0.064 | 0.063 | 0.062 | 0.062 | 0.056 | 0.056 | 0.057 | 0.052 | 0.049 | 0.060 | 0.069 | 0.070 | 0.065 | 0.047 | |||||
|
| 0.067 | 0.068 | 0.064 | 0.071 | 0.063 | 0.065 | 0.063 | 0.065 | 0.065 | 0.062 | 0.080 | 0.076 | 0.073 | 0.068 | 0.065 | 0.073 | ||||
|
| 0.076 | 0.077 | 0.083 | 0.083 | 0.078 | 0.065 | 0.070 | 0.069 | 0.063 | 0.057 | 0.056 | 0.057 | 0.081 | 0.077 | 0.060 | 0.066 | 0.082 | |||
|
| 0.088 | 0.093 | 0.098 | 0.097 | 0.086 | 0.078 | 0.092 | 0.086 | 0.077 | 0.066 | 0.066 | 0.080 | 0.100 | 0.092 | 0.073 | 0.079 | 0.102 | 0.053 | ||
|
| 0.087 | 0.091 | 0.098 | 0.102 | 0.096 | 0.084 | 0.104 | 0.100 | 0.087 | 0.073 | 0.074 | 0.089 | 0.110 | 0.098 | 0.084 | 0.089 | 0.106 | 0.067 | 0.051 | |
|
| 0.076 | 0.075 | 0.066 | 0.073 | 0.067 | 0.057 | 0.083 | 0.072 | 0.061 | 0.058 | 0.074 | 0.077 | 0.084 | 0.086 | 0.062 | 0.070 | 0.085 | 0.069 | 0.081 | 0.089 |
| RPB2 rDNA | ||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
| |||||||||
|
| 0.208 | |||||||||||||||||||
|
| 0.251 | 0.215 | ||||||||||||||||||
|
| 0.234 | 0.241 | 0.286 | |||||||||||||||||
|
| 0.273 | 0.256 | 0.293 | 0.285 | ||||||||||||||||
|
| 0.252 | 0.259 | 0.270 | 0.309 | 0.262 | |||||||||||||||
|
| 0.259 | 0.247 | 0.285 | 0.256 | 0.303 | 0.304 | ||||||||||||||
|
| 0.262 | 0.229 | 0.271 | 0.296 | 0.296 | 0.309 | 0.313 | |||||||||||||
|
| 0.280 | 0.286 | 0.280 | 0.299 | 0.298 | 0.298 | 0.296 | 0.301 | ||||||||||||
|
| 0.238 | 0.262 | 0.319 | 0.308 | 0.307 | 0.311 | 0.311 | 0.293 | 0.340 | |||||||||||
|
| 0.276 | 0.282 | 0.344 | 0.312 | 0.315 | 0.316 | 0.349 | 0.323 | 0.353 | 0.285 | ||||||||||
|
| 0.326 | 0.305 | 0.315 | 0.315 | 0.338 | 0.324 | 0.367 | 0.298 | 0.342 | 0.349 | 0.375 | |||||||||
|
| 0.376 | 0.392 | 0.420 | 0.418 | 0.392 | 0.426 | 0.433 | 0.399 | 0.440 | 0.366 | 0.397 | 0.446 | ||||||||
|
|
|
|
|
|
|
| |||
|---|---|---|---|---|---|---|---|---|---|
|
|
| ||||||||
|
| 2–2.5 µm, hyaline, | 7–60 × 2–3.5 µm, brown, 0–7-septate | 7–25 × 2–3.5 µm, brown, with flat-tipped denticles (0.5–1.5 × 0.5 µm) | narrowly fusoid ellipsoid, 2-septate, hyaline | (16–)25– 34(–37) × 2(–2.5) µm | epiphytic | USA |
| |
|
| 1–3 µm, hyaline to moderately brown | 5–20 µm × 3–3.5 µm, moderately brown, cylindrical, 0–2-septata | cylindrical, moderately brown, with successive denticles and geniculate structure | cylindrical, 2-septata, individually, hyaline to pale smoky | (22–) 27–33 (–35) × 1.5–2 µm | saprobic | Japan |
| |
|
| – | – | – | – | – | saprobic | Undefined ( | Brazil |
|
|
| 2–3 µm, hyphae | hyaline (appearing subhyaline with age), mostly reduced to conidiogenous cells | 10–25 × 3–4 µm hyaline, prominent cylindrical denticles, 1–3 × 1.5 µm | spindle-shaped, apex subobtuse, base truncate, (3–)5–6(–8)-septate, hyaline | (37–)40–45(–47) × (2.5–)3 µm | epiphytic | Brazil |
| |
|
| pale brown | 15–30 × 4 μm at the base, 0–3 thin septa, subhyaline to pale brown | thin-walled, hyaline, slightly lobed; denticles absent; rhexolytic secession with inconspicuous scars | clavate, 2-septate, hyaline | 18–26 × 1.5, base 0.6 µm wide | saprobic | Netherlands | de Hoog and van Oorscho (1985) | |
|
| – | 6–35 × 2.2–2.8 µm, hyaline, aseptate or septate | 2–2.8 μm wide,terminal, integrated, hyaline | clavate, blunt end,hyaline, 1–5 septate | 10–60 × 2.2–2.8 μm | endophytic | Korea |
| |
|
| 1.7–2.6 µm, hyaline | 5–37 × 2–4 µm, hyaline, septate, sometimes reduced to conidiogenous cells | 4–18 × 1–4 µm, with conspicuous, cylindrical denticles, up to 0.9 µm wide | narrowly fusoid-ellipsoid, guttulate, 0–3-septate, hyaline | 20–40 × 1.5–4 µm | endophytic | China | This study | |
|
| – | – | – | – | – |
| an unidentified ornamental plant | Australia |
|
|
| 1–1.5 μm, colorless | 15–40 × 2–2.5 μm, colorless, septate | 12–17 × 2–3 µm, with conspicuous, truncate denticles in the apical region | fusiform, 1-septate, colourless | 30–37 × 1–1.5 µm | saprobic | Cuba |
| |
|
| 1.5–2.5 μm, hyaline | conidiophores reduced to conidiogenous cells or with supporting cell | with 1–2 × 1 μm apex denticulate | medianly 1-septate, straight to narrowly fusoid, 6–20 × 2.5–3.5 | (26–)30–33(–37) × (1.5–)2 µm | epiphytic | Colombia |
| |
|
| 1–1.5 μm, pale brown | 0–1-septate, pale brown usually reduced to a conidiogenous cell. | 7–12 × 3–4 µm, pale brown or almost colorless, with large, conspicuous, truncate denticles, 1.5–2 μm | subcylindrical, (2–)3-septated with a false septum near each end, colorless or almost colorless | 26–36 × 1.5–2 µm | saprobic | decaying leaves (unidentified) | Cuba |
|
|
| 1–1.5 μm, light brown | 12–60 × 1–2 µm, septate, light brown, up to 24 µm wide at apex | polyblastic, denticulate, sympodial, inflated at apex | cylindrical, 2-septata, hyaline, septa visible near extremities | 18–26 × 1–1.5 µm | saprobic | Cuba | Castañeda Ruiz (1988) | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Lichen and fungal ecology · Mycorrhizal Fungi and Plant Interactions
Introduction
Dactylaria was established by Saccardo (1880) with D. purpurella as the type species, originally identified from dead leaves of Castanopsis cuspidata var. sieboldii in Japan. De Hoog (1985) expanded the circumscription of Dactylaria, recognizing 41 species classified into four sections: Dactylaria, Mirandina (G. Arnaud ex Matsush.) de Hoog, Diplorhinotrichum (Höhn.) de Hoog, and Pleurophragmium (Constantin) de Hoog. Later, Goh and Hyde (1997) expanded this framework by providing a taxonomic key for 37 additional species associated with plant material, increasing the number of recognized species to 82. Subsequent to Goh and Hyde’s (1997) review, a further 14 species have been described in Dactylaria (Paulus et al. 2003). The genus has undergone considerable taxonomic changes in recent years. Several new species have been described (Cooper 2005; Barbosa et al. 2013; Mel’nik et al. 2013; Magalhães et al. 2014; Crous et al. 2016; Crous et al. 2022; Tan and Shivas 2024; Crous et al. 2025; de Souza et al. 2025; Liu et al. 2025), while the application of molecular phylogenetic data has resulted in the transfer of some species to other genera, such as Dactylaria hyalotunicata, which is now recognized as Pseudodactylaria hyalotunicata (Crous et al. 2017). Conversely, species from other genera have been reassigned to Dactylaria based solely on traditional morphology, including Subulispora argentina and S. malaysiana (de Souza et al. 2025), although molecular evidence supporting these transfers is still lacking.
Currently, 134 epithets are listed under Dactylaria in Index Fungorum (2025) with 117 possible species listed in Species Fungorum (2025). Traditionally, members of Dactylaria are characterized by hyaline, cylindrical, rachis-bearing conidiogenous cells with apical, scattered, sympodial, tubular, or cylindrical denticles, and their conidia are hyaline, cylindrical, fusiform, or navicular, and can be unicellular or septate, with liberation occurring through schizolytic secession (de Hoog 1985; Goh and Hyde 1997; Paulus et al. 2003). Dactylaria species exhibit diverse ecological roles, with most species functioning as saprobes or parasites on a wide range of substrates including soil (Bhatt and Kendrick 2011) and plant materials (Paulus et al. 2003; Pinnoi et al. 2003; Senwanna et al. 2021; Wijesinghe et al. 2022). Additionally, some species act as endophytes that colonize living plant tissues (Wu et al. 1996), while others serve as nematode-trapping fungi (Krizková et al. 1976; Kumar and Singh 2011), plant pathogens, or opportunistic agents of animal diseases (Kralovic and Rhodes 1995; Goh and Hyde 1997).
To date, only 31 species have available sequence data in GenBank, with these data primarily corresponding to the LSU (large subunit ribosomal RNA gene) and SSU (small subunit ribosomal RNA gene) regions. No comprehensive, genus-wide phylogenetic study including the type species has been conducted and the genus’ systematic placement remains poorly resolved. Most critically, the phylogenetic position of its type species, D. purpurella, has yet to be clarified (Bussaban et al. 2005; Réblová 2009; Lin et al. 2021). LSU-based analyses conducted by Réblová (2009), which primarily addressed the sexual morph of Rhodoveronaea and the reevaluation of Pleurophragmium, demonstrated that D. purpurella, D. monticola, and D. parvispora occupy distinct lineages. The results excluded Dactylaria from synonymy with Pleurophragmium and further highlighted the polyphyletic nature of the genus. Despite its long taxonomic history, Dactylaria is taxonomically heterogeneous, leading to persistent controversy regarding its definition and classification (Goh and Hyde 1997; Paulus et al. 2003). Most species were historically established solely on morphological characteristics, particularly before the application of molecular tools (Goh and Hyde 1997; Cooper 2005; Flavia et al. 2013; Mel’nik et al. 2013; Magalhães et al. 2014). In recent years, with the advent of molecular techniques, newly described Dactylaria species have incorporated both morphological and molecular data, though typically limited to single-gene analyses. For example, D. acaciae (Crous et al. 2016), D. retrophylli (Crous et al. 2022), D. calliandrae (Crous et al. 2025), and D. hwasunensis (Liu et al. 2025) were identified using LSU sequences, whereas D. mavisleverae (Tan and Shivas 2024) was confirmed based on ITS sequences.
Interestingly, recent phylogenetic analyses of dactylaria-like taxa have revealed that Funiliomyces biseptatus consistently clusters with several “Dactylaria” species, forming a well-supported clade with uncertain taxonomic position within the order Amphisphaeriales (Crous et al. 2016, 2022, 2025). Funiliomyces biseptatus, the type and currently the only described species of the genus Funiliomyces, was established by Aptroot (2004) from a dead leaf of a Bromeliaceae plant. The genus is characterized by its fusiform ascospores with two submedian septa and furnished with both apical and median appendages. Notably, F. biseptatus represents a sexual morph, whereas the associated “Dactylaria” species are asexual forms. The close phylogenetic affinity between the sexual morph of Funiliomyces and asexual morphs of dactylaria-like taxa forms a robust, statistically well-supported clade that is phylogenetically distinct from all currently described families within Amphisphaeriales.
Amphisphaeriales was introduced by Eriksson and Hawksworth (1986). Subsequent multigene phylogenetic studies have consistently demonstrated that Amphisphaeriales and Xylariales represent distinct orders within the subclass Xylariomycetidae (Senanayake et al. 2015; Samarakoon et al. 2016; Hongsanan et al. 2017; Crous et al. 2018b). According to molecular dating analyses, the crown age of Amphisphaeriales has been estimated at around 133 MYA (Hyde et al. 2020). Members of Amphisphaeriales are characterized by gelatin-covered, interascal, tapering or reticulate paraphyses, host-immersed perithecia with or without a clypeus, hymenial asci with amyloid apices and apical rings, variable, hyaline or pigmented ascospores with variable septa, and holoblastic conidiogenesis with denticulate conidia and solitary or united conidiophores (McLaughlin and Spatafora 2014). The circumscription of Amphisphaeriales by Hyde et al. (2020) included 17 families: Amphisphaeriaceae, Appendicosporaceae, Apiosporaceae, Beltraniaceae, Castanediellaceae, Clypeophysalosporaceae, Cylindriaceae, Hyponectriaceae, Iodosphaeriaceae, Melogrammataceae, Oxydothidaceae, Phlogicylindriaceae, Pseudomassariaceae, Pseudosporidesmiaceae, Pseudotruncatellaceae, Sporocadaceae, and Xyladictyochaetaceae, while Vialaeaceae was excluded. Hyde et al. (2024) later incorporated Vialaeaceae into the order. In addition, two families originally placed in Xylariales, Nothodactylariaceae, and Anungitiomycetaceae (Crous et al. 2019b), have been transferred to Amphisphaeriales based on phylogenetic evidence (Réblová et al. 2021; Zhang et al. 2025).
Endophytic fungi associated with medicinal plants are well recognized as a rich reservoir of structurally diverse bioactive metabolites, and their interactions with host plants often contribute to the biosynthesis of pharmacologically important compounds, making them a focal point for natural product and fungal taxonomy research (Tiwari and Bae 2022; Wen et al. 2022; Gupta et al. 2023; Chandra et al. 2024). Tetradium ruticarpum (A.Juss.) Hartley, commonly known as Evodia rutaecarpa or Wu Zhu Yu in Chinese, is a medicinal plant of high value in traditional Chinese medicine (Chinese Pharmacopoeia Commission 2010). Its fruits are widely used for their pharmacological activities, including antitumor, cardiovascular protective, anti-inflammatory, antimicrobial, antioxidant, and anti-obesity effects (Li and Wang 2020; Xiao et al. 2023). Research on endophytic fungi associated with T. ruticarpum remains limited. Previously reported endophytes include Sclerotium Tode ex Fr., which produces multiple bioactive compounds (Zhu 2007), as well as other taxa identified only to the genus level, such as Cyanodermella, Guignardia, Hypoxylon, and Nigrospora (Ho et al. 2012). Two novel species, Cyphellophora guangxiensis (Mi et al. 2025b) and Pseudokeissleriella tetradii (Mi et al. 2025a), were recently reported from T. ruticarpum.
In this study, we elucidate the long-recognized polyphyly of Dactylaria**sensu lato. Phylogenetic analyses based on the LSU gene show that species historically assigned to this genus, for which LSU sequence data are available, are distributed across at least six distinct classes within Pezizomycotina, further supporting its non-monophyletic status. Phylogenetic inference using a concatenated LSU-ITS-RPB2 dataset and interfamily genetic differences recover a monophyletic lineage within Amphisphaeriales, comprising Funiliomyces biseptatus, ten species previously classified under Dactylaria, and our newly isolated strains. We thus formally propose the new family Funiliomycetaceae fam. nov. to accommodate this well-delimited clade. Additionally, we describe one new species Funiliomyces jiangxiensis from Tetradium ruticarpum, formally transfer the ten aforementioned Dactylaria species to Funiliomyces as new combinations, and present a table summarizing the asexual morphological features, lifestyle, host associations, and distribution of all known Funiliomyces species.
Materials and methods
Collection, isolation, and morphological studies
The healthy roots of Tetradium ruticarpum were collected from Jiangxi Province, China. The samples were placed into sterile self-sealing plastic bags, transported to the lab and stored at 4 °C until processing. Tissue isolation and culture method were carried out by following the method of Deng et al. (2014). The specific disinfection time was adjusted according to T. ruticarpum. The roots were repeatedly washed by using running tap water and air dried. The surface of each sample was disinfected using 75% ethanol for 1 min, and 3.3% sodium hypochlorite solution for 1 min, followed by three rinses in sterile distilled water before finally being dried on sterile filter papers. After disinfection, the roots were cut into small pieces (ca. 5 mm^2^) by a sterile scalpel and placed in Potato Dextrose Agar plates (PDA), incubated at 25 °C in darkness. When the fungi develop from the tissue segment, a few hyphal fragments were picked up and transferred to new PDA plates to obtain pure cultures.
Micromorphological characteristics from pure cultures were observed and captured using a Nikon ECLIPSE Ni-U compound microscope (Nikon Corporation, Japan), equipped with a Nikon DS-Fi3 camera. All measurements were calculated using PhotoRuler v. 1.1 software (The Genus Inocybe, Hyogo, Japan). For conidiophores and conidia, morphological data are presented as range followed by mean ± standard deviation (SD). The figures were processed using Adobe Photoshop CS6 Extended v.10.0 software (Adobe Systems, USA). The fungal cultures were deposited in the Jiangxi Agricultural University Culture Collection (JAUCC), Nanchang, China, and the China Center for Type Culture Collection (CCTCC), respectively. The dry cultures were deposited in the Herbarium of Fungi, Jiangxi Agricultural University (HFJAU), Nanchang, China. The newly introduced fungal species was submitted to the Index Fungorum (2025) and Faces of Fungi database (Jayasiri et al. 2015).
DNA extraction, PCR amplification and sequencing
Fresh mycelium was extracted using the CTAB method (Doyle and Doyle 1987). According to the locus-specific databases of the Dactylaria species (Crous et al. 2016; Crous et al. 2022), three loci (LSU, ITS, and RPB2) were chosen for polymerase chain reaction (PCR) using the primer pairs LR0R/LR5 (Vilgalys and Hester 1990), ITS1/ITS4 (White et al. 1990), and fRPB2-5F/fRPB2-7CR (Liu et al. 1999), respectively. The final volume of the PCR reaction mixture was 25 μL, contained 12.5 μL 2×Taq PCR MasterMix (Qingke, Changsha, China), 1 μL each forward and reverse primer (0.2 μM), 1 μL template DNA (circa 50–100 ng), and 9.5 μL ddH_2_O. For ITS and LSU genes, the PCR parameters were initial denaturation 94 °C for 5 min; followed by 35 cycles consisting of denaturation at 94 °C for 30 seconds, annealing at 55 °C for 50 seconds and extension at 72 °C for 60 seconds; and final extension at 72 °C for 10 min. For RPB2 gene, the PCR parameters were initial denaturation 94 °C for 5 min; followed by 40 cycles consisting of denaturation at 94 °C for 60 seconds, annealing at 58 °C for 60 seconds and extension at 72 °C for 60 seconds; and a final extension at 72 °C for 10 min. The PCR products were purified and sequenced by QingKe Biotechnology Co. (Changsha, China). All sequences were assembled using BioEdit v.7.2.6 (Hall 1999). The new sequences were deposited in GenBank (https://www.ncbi.nlm.nih.gov/). The sequence alignments and resulting phylogenetic trees have been deposited in TreeBASE (www.treebase.org).
Reference datasets
We combined the new ribosomal and protein coding sequences with data from the National Center for Biotechnology Information (NCBI). Accession numbers for all sequences used for the molecular analyses are provided in Tables 1, 2.
-
Pezizomycotina: To determine the phylogenetic placement of Dactylaria within Pezizomycotina, we compiled a dataset representing all major ascomycete classes and conducted LSU single-gene analyses (Beimforde et al. 2020). Of the 31 Dactylaria species with molecular data in GenBank, five (D. acerosa, D. appendiculata, D. candidula, D. junci, and D. lanosa) lack LSU sequences and were excluded. The final dataset included 124 species, comprising 26 Dactylaria species. Accession numbers for all sequences are provided in Table 1.
-
Amphisphaeriales: To assess the placement of our two new strains, 71 taxa were selected based on the Pezizomycotina phylogeny of Dactylaria and recent publications (Crous et al. 2016, 2022, 2025; Tan and Shivas 2024; Liu et al. 2025). Phylogenetic analyses of Dactylaria species affiliated with Amphisphaeriales were performed using LSU, ITS, and RPB2 sequences. Accession numbers for all sequences are listed in Table 2.
Phylogenetic analyses
Phylogenetic analyses were conducted using Maximum Likelihood (ML) and Bayesian Inference (BI). Sequences for both datasets were obtained from GenBank (http://www.ncbi.nlm.nih.gov/). All sequences were aligned with the online version of MAFFT v. 7 (https://mafft.cbrc.jp/alignment/server/) (Katoh and Standley 2013) and manually refined in BioEdit v. 7.2.6 (Hall 1999), with gaps treated as missing data. Dataset 1 contained only LSU sequences, whereas for Dataset 2, LSU, ITS, and RPB2 sequences were concatenated into a supermatrix using PhyloSuite v. 1.2.2 (Zhang et al. 2020). Alignment file formats were subsequently converted with ALTER (Alignment Transformation Environment; http://www.sing-group.org/ALTER/).
ML analyses were carried out on the dataset using IQ-TREE v.1.6 (http://iqtree.cibiv.univie.ac.at/) with partitioning, and the optimal substitution model for each partition was determined by IQ-TREE. (Trifinopoulos et al. 2016). Clade support for the ML analyses was assessed using the Shimodaira-Hasegawa-like approximate likelihood ratio test (SH-aLRT) with 1,000 replicates (Guindon et al. 2010) and 1,000 replicates of the ultrafast bootstrap (UFB) (Hoang et al. 2017). Nodes with support values of both SH-aLRT ≥ 80 and UFB ≥ 95 were considered supported, nodes with one of SH-aLRT ≥ 80 or UFB ≥ 95 were weakly supported, and nodes with both SH-aLRT < 80 and UFB < 95 were unsupported, and other parameters were used for the default settings (Liu et al. 2022).
Bayesian Inference (BI) analyses were performed on the CIPRES Science Gateway (http://www.phylo.org/) (Miller et al. 2010) using MrBayes 3.2.7 (Ronquist et al. 2012). For both datasets, each gene partition was analyzed under the GTR+I+G model, which was selected as the best-fit model using MrModeltest 2.3 in PAUP* v4.0b (Swofford 2002; Nylander 2004). Two independent Markov Chain Monte Carlo (MCMC) runs were conducted for 10,000,000 generations, sampling every 1,000 generations, with the first 25% of samples discarded as burn-in. Convergence was assessed by ensuring the average standard deviation of split frequencies (ASDSF) fell below 0.01. Bayesian posterior probabilities (BPP) were used to assess node support (Rannala and Yang 1996). Resulting trees were visualized in FigTree v. 1.4.4 (Rambaut 2018) and edited using the online service tvBOT (https://www.chiplot.online/tvbot.html) (Xie et al. 2023). Genetic distances were estimated using the Kimura 2-parameter (K2P) model in MEGA-X (Kumar et al. 2018) based on ITS, LSU, and RPB2 sequences of species representing different family within Amphisphaeriales.
Results
Molecular analysis
Based on a nucleotide BLAST search in GenBank database using the LSU sequence, the closest matches are Dactylaria species, including Funiliomyces hwasunensis (≡ D. calliandrae) CMML 20-35 [GenBank PQ741487; identities = 820/824 (99%), gaps = 0/824 (0%)], F. calliandrae (≡D. calliandrae) CPC 48004 [GenBank PV664963; identities = 805/811 (99%), gaps = 0/811 (0%)], F. fragilis (≡ D. fragilis) P057 [GenBank EU107290; identities = 798/807 (99%), gaps = 0/811 (0%)]. The closest match for the ITS sequence is Fusidium griseum Trtsf08 [GenBank GU479905; identities = 468/495 (95%), gaps = 7/495 (1%)], D. acerosa ICMP 13178 [GenBank OR543730; identities = 465/494 (94%), gaps = 4/494 (4%)], and F. calliandrae (≡ D. calliandrae) CPC 48004 [GenBank PV664937; identities = 454/479 (95%), gaps = 6/479 (4%)]. For the RPB2 sequence, the closest matches included Dicyma funiculosa CBS 323.86 [GenBank KU684306; identities = 579/735 (79%), gaps = 6/735 (0%)], and Xylaria liquidambaris FCATAS879 [GenBank MZ707110; identities = 553/700 (79%), gaps = 8/700 (1%)]. This situation may be due to the limited availability of RPB2 gene sequences for this genus.
Phylogenetic analysis
The phylogenetic hypothesis inferred from the LSU gene dataset for Pezizomycotina is presented in Fig. 1, with Candida albicans (ATCC 18804) and Saccharomyces cerevisiae (NRRL Y-12632) selected as outgroup taxa to root the tree, following previous large-scale phylogenetic studies of Pezizomycotina (Beimforde et al. 2020). The LSU alignment comprised 898 bp (including gaps), with 445 parsimony-informative sites, 94 singletons, and 359 constant sites. Maximum likelihood (ML) analysis recovered 629 distinct alignment patterns, with 7.33% undetermined characters or gaps. The best-scoring ML tree (–lnL = 22860.942) is shown in Fig. 1. Overall, the topologies obtained from ML and Bayesian inference (BI) analyses were highly congruent, differing only in the placement of a few taxa and in several unresolved clades. In this analysis, twenty-two Dactylaria species are distributed across six different classes within Pezizomycotina, namely Orbiliomycetes (one species), Leotiomycetes (one species), Eurotiomycetes (two species), Sordariomycetes (16 species), Dothideomycetes (one species), and Lecanoromycetes (one species). The remaining four species could not be confidently assigned to any currently recognized class of Pezizomycotina. The phylogenetic tree reveals that Dactylaria is polyphyletic, with most species clustering in Sordariomycetes. The exact taxonomic position of the type species, D. purpurella, remains uncertain. It forms a distinct lineage with D. humicola and D. dimorpha, representing an incertae sedis lineage in Pezizomycotina. In addition, our newly isolated strain clustered in the Sordariomycetes clade with ten species of Funiliomyces (formerly treated as Dactylaria) in Funiliomycetaceae fam. nov. with branch support values of SH-aLRT = 87.4%, UFB = 94%, and BPP = 0.88 (UFB value < 95% and BPP < 0.90 are not shown on the tree) (Fig. 1).
Maximum likelihood phylogenetic tree of Pezizomycotina based on LSU sequences, with Shimodaira-Hasegawa-like approximate likelihood ratio test (SH-aLRT) (left), ultrafast bootstrap (UFB) (middle), and Bayesian posterior probabilities (BPP) values (right) near the corresponding node. Only one of SH-aLRT > 80 or UFB > 95 for ML and BYPP > 0.90 for BI is indicated along the branches (SH-aLRT/UFB/BPP). The “Dactylaria” species are indicated in red, the newly generated sequences are indicated in green, and species for reclassification from Dactylaria to Funiliomyces are in blue. Ex-type strains are marked with T after the strain number. The ML phylogram is available in TreeBASE (study accession S32460; http://purl.org/phylo/treebase/phylows/study/TB2:S32460).
The phylogenetic tree of Amphisphaeriales based on the three-gene dataset, rooted with Achaetomium macrosporum (CBS 532.94), Chaetomium elatum (CBS 374.66), and Sordaria fimicola (CBS 723.96), is shown in Fig. 2. The combined dataset of LSU, ITS, and RPB2 consisted of 2642 bp including gaps (LSU 857 bp, ITS 728 bp, and RPB2 1057 bp), which contained 1501 parsimony-informative characters, 247 singleton sites, and 1339 constant characters. The maximum likelihood matrix had 1804 distinct alignment patterns with 37.19% undetermined characters or gaps. The best maximum likelihood tree, with a final likelihood value of -29373.458, is shown in Fig. 2. The ML tree and BI analysis tree were similar, except for some interchanged positions and some unseparated clade. In this study, phylogenetic analyses revealed that our two new strains, together with ten previously described “Dactylaria” species and one Funiliomyces species, formed a distinct yet unplaced lineage within Amphisphaeriales (Fig. 2). The overall tree topology (Fig. 2) recovered in this study is largely congruent with those reported by Crous et al. (2022, 2025). Our phylogenetic analyses further confirmed that Dactylaria sensu lato is polyphyletic. Dactylaria purpurella, the type species of the genus, forms an independent lineage within Pezizomycotina and is distantly related to the Amphisphaeriales clade comprising ten “Dactylaria” taxa. (Fig. 1). Accordingly, these species in Amphisphaeriales are not congeneric with the type of Dactylaria. Based on phylogenetic evidence and following the principles of monophyly and the “One Fungus, One Name” concept, these ten species are here transferred to the genus Funiliomyces. Although Funiliomyces biseptatus is only known from its sexual morph, whereas the transferred species are asexual, phylogenetic placement indicates that they represent different morphs of the same genus. In addition, our analyses also showed that the two newly generated strains, CCTCC AF 2023033 and CCTCC AF 2023034, form a distinct branch and cluster as sister lineages with Funiliomyces hwasunensis (≡ Dactylaria hwasunensis) and F. calliandrae (≡ D. calliandrae), supported by SH-aLRT = 98.8%, UFB = 100%, and Bayesian posterior probability = 0.98 (Fig. 2), indicating that they represent a phylogenetically novel species in Funiliomyces.
Maximum likelihood phylogenetic tree of Amphisphaeriales based on combined LSU, ITS, and RPB2 sequences, with Shimodaira-Hasegawa-like approximate likelihood ratio test (SH-aLRT) (left), ultrafast bootstrap (UFB) (middle), and Bayesian posterior probabilities (BPP) values (right) near the corresponding node. Only one of SH-aLRT > 80 or UFB > 95 for ML and BYPP > 0.90 for BI is indicated along the branches (SH-aLRT/UFB/BPP). Ex-type strains are marked with T after the strain number. The newly generated sequences are indicated in red and species for reclassification are in blue. The ML phylogram is available in TreeBASE (study accession S32462; http://purl.org/phylo/treebase/phylows/study/TB2:S32462).
Furthermore, the phylogenetic tree inferred from the combined three-locus dataset resolved twenty main clades within Amphisphaeriales (Fig. 2), corresponding to the families Beltraniaceae (98.8\100\1.0), Amphisphaeriaceae (100\100\1.0), Castanediellaceae (96\100\1.0), Sporocadaceae (100\100\1.0), Phlogicylindriaceae (99.6\100\1.0), Xyladictyochaetaceae (-\95\0.98), Clypeophysalosporaceae (99.7\100\1.0), Cylindriaceae (100\100\1.0), Pseudomassariaceae (98.2\99\1.0), Apiosporaceae (100\100\1.0), Melogrammataceae (100\100\1.0), Pseudotruncatellaceae (100\100\1.0), a distinct clade formed by Funiliomyces (86.6--), Nothodactylariaceae (96.1\98\1.0), Oxydothidaceae (100\100\1.0), Appendicosporaceae (92.8\99\1.0), Hyponectriaceae (-/-/-), Anungitiomycetaceae (100\100\1.0), Vialaeaceae (100\100\1.0), Pseudosporidesmiaceae (100\100\1.0), and Iodosphaeriaceae (100\100\1.0). Both Bayesian inference and maximum likelihood analyses supported the monophyly of this Funiliomyces clade and its sister relationship to Nothodactylariaceae (SH-aLRT = 99.9%, UFB = 94%, BPP = 0.99; UFB < 95% is not shown on the tree). Based on these phylogenetic results, a new family, Funiliomycetaceae fam. nov., is proposed to accommodate the Funiliomyces lineage.
Genetic distance analysis among families within Amphisphaeriales
To further clarify the phylogenetic position of the “Dactylaria” lineage within Amphisphaeriales, we conducted genetic divergence analyses based on ITS, LSU, and RPB2 sequences.
Within Amphisphaeriales, the ITS genetic distances among 19 families range from 0.110 (between Sporocadaceae and Beltraniaceae) to 0.369 (between Pseudosporidesmiaceae and Iodosphaeriaceae), indicating substantial inter-family variation in this locus (Table 3). When focusing on Funiliomycetaceae, its ITS genetic distances to the other 19 families range from 0.156 to 0.306, with the smallest distance to Nothodactylariaceae (0.156) and the largest to Iodosphaeriaceae (0.306). Notably, the minimum divergence of Funiliomycetaceae from other families (0.156) is still greater than the smallest inter-family distance within the order (0.110), suggesting that Funiliomycetaceae is not closely allied with any established family.
For the LSU dataset, the genetic distances among the 20 families range from 0.032 between Xyladictyochaetaceae and Phlogicylindriaceae to 0.110 between Sporocadaceae and Iodosphaeriaceae. The LSU distances between Funiliomycetaceae and the other families vary from 0.036 with Pseudotruncatellaceae to 0.084 with Iodosphaeriaceae. Again, the smallest LSU distance between Funiliomycetaceae and Pseudotruncatellaceae (0.036) remains higher than the overall minimum value (0.032) observed within the order, supporting its independent molecular identity.
For the RPB2 dataset, only 13 families were included because not all families within Amphisphaeriales have publicly available RPB2 sequences. Nevertheless, the inter-family distances reveal clear divergence of Funiliomycetaceae. Overall, the inter-family distances range from 0.208 between Melogrammataceae and Beltraniaceae to 0.446 between Anungitiomycetaceae and Oxydothidaceae. The RPB2 distances between Funiliomycetaceae and the other families range from 0.252 with Beltraniaceae to 0.426 with Anungitiomycetaceae. The minimum distance between Funiliomycetaceae and Beltraniaceae (0.252) is notably higher than the smallest inter-family distance in the dataset (0.208), indicating consistent divergence.
Collectively, the analyses based on ITS, LSU, and RPB2 sequences demonstrate that Funiliomycetaceae is genetically distinct from all other families within Amphisphaeriales. Across three loci, its inter-family distances are consistently greater than the smallest distances observed among other established families, providing robust molecular evidence that Funiliomycetaceae represents an independent family within the order.
Taxonomy
Funiliomycetaceae
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
L.X. Mi, K.D. Hyde, H.Y. Song & D.M. Hu fam. nov.
1F5906FE-B7AA-56E9-BDE3-BAAB5CA80D61
Index Fungorum: IF904529
Facesoffungi Number: FoF18896
Etymology.
Name refers to the type genus Funiliomyces.
Type genus.
Funiliomyces Aptroot, Studies in Mycology 50(2): 309 (2004).
Description.
Saprobic, endophytic, or epiphytic on diverse plant hosts in tropical to temperate regions. Sexual morph: Ascomata black, subglobose, immersed to erumpent. Wall composed of irregular layers of regularly melanized, flattened cells, with no color change in KOH. Paraphyses absent. Asci cylindrical, 8-spored, with a thickened apex bearing a central refractive, IKI-negative apical apparatus, and enclosed by parenchymatous tissue. Ascospores pale brown, torpedo-shaped, 2-septate, upper cell pointed, lower cell rounded, hyaline granules or oil droplets, bearing two hyaline mucilaginous appendages. Asexual morph: Mycelium consisting of branched, septate, smooth, hyaline to pale brown hyphae, sometimes forming hyphal ropes. Conidiophores macronematous, mononematous, solitary or in small groups, erect, straight to flexuous, subcylindrical to cylindrical (apex sometimes inflated), simple or occasionally branched, hyaline to brown, septate, sometimes reduced to conidiogenous cells. Conidiogenous cells integrated, terminal or lateral, sympodial, mono- to polyblastic, hyaline to brown, cylindrical to clavate, with flat-tipped or denticulate apices; denticles (when present) large, cylindrical to geniculate, cylindrical, truncate, or pimple-like, or lacking entirely. Conidia solitary, hyaline to pale smoky, smooth, septate, narrowly fusiform to cylindrical, with obtuse, subobtuse, or tapering apices and truncate or rounded bases; dimensions variable among species.
Notes.
A new family, Funiliomycetaceae, is proposed to accommodate a distinct, strongly supported clade comprising the genus Funiliomyces and several related lineages historically identified as “Dactylaria”. Funiliomycetaceae shares general sexual characters with Amphisphaeriales, including immersed to semi-immersed ascomata and unitunicate asci with an apical ring (Aptroot 2004). However, the family is readily distinguished from all currently accepted families in the order by its unique ascospore morphology: the ascospores are fusiform, consistently biseptate, and bear both apical and median appendages, a combination of characters not reported in other families of Amphisphaeriales (Aptroot 2004). The asexual morph of Funiliomycetaceae is hyphomycetous, characterized by conidiogenous cells that are sympodial, terminal or lateral, integrated, and denticulate (when present, denticles are large and cylindrical to geniculate), producing solitary, hyaline to pale smoky, smooth, septate, narrowly fusiform to cylindrical conidia with obtuse to tapering apices and truncate or rounded bases. Funiliomycetaceae shares similar sexual features with Nothodactylariaceae (Nothodactylaria), such as Nothodactylaria nephrolepidis (Cours et al. 2019b). However, Funiliomycetaceae is phylogenetically distinct from the families in Amphisphaeriales, as supported by multi-locus phylogenetic analyses (Fig. 2) and inter-familial genetic distance comparisons (Table 3).
Funiliomyces
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
Aptroot, Stud. Mycol. 50 (2): 309 (2004)
8987BC25-26FA-557A-A755-8B01ACE7817F
Index Fungorum: IF500077
Facesoffungi Number: FoF18897
Type species.
Funiliomyces biseptatus Aptroot, Stud. Mycol. 50 (2): 309 (2004). Index Fungorum: IF500164.
Holotype.
CBS H-10505.
Type information.
Brazil, Minas Gerais, Catas Altas, Serro do Caraça, Parque Natural do Caraça, near Funil, 1 km NW of monastery Santuário do Caraça, 20°06'S, 43°29'W, on dead leaf of Bromeliaceae in rock field, 18 Sept. 1997, A. Aptroot, holotype herb. CBS H-10505, isotypes herb. SP, living culture ex-type CBS 100373, also dried culture CBS H-10506.
Description.
See Aptroot (2004) on page 309.
Emended diagnosis.
Saprobic, endophytic, or epiphytic on diverse plant hosts in tropical to temperate regions. Sexual morph: Ascomata black, subglobose, immersed to erumpent. Wall composed of irregular layers of regularly melanized, flattened cells, with no color change in KOH. Paraphyses absent. Asci cylindrical, 8-spored, with a thickened apex bearing a central refractive, IKI-negative apical apparatus, and enclosed by parenchymatous tissue. Ascospores pale brown, torpedo-shaped, 2-septate, upper cell pointed, lower cell rounded, hyaline granules or oil droplets, bearing two hyaline mucilaginous appendages. Asexual morph: Mycelium consisting of branched, septate, smooth, hyaline to pale brown hyphae, sometimes forming hyphal ropes. Conidiophores macronematous, mononematous, solitary or in small groups, erect, straight to flexuous, subcylindrical to cylindrical (apex sometimes inflated), simple or occasionally branched, hyaline to brown, septate, sometimes reduced to conidiogenous cells. Conidiogenous cells integrated, terminal or lateral, sympodial, mono- to polyblastic, hyaline to brown, cylindrical to clavate, with flat-tipped or denticulate apices; denticles (when present) large, cylindrical to geniculate, cylindrical, truncate, or pimple-like, or lacking entirely. Conidia solitary, hyaline to pale smoky, smooth, septate, narrowly fusiform to cylindrical, with obtuse, subobtuse, or tapering apices and truncate or rounded bases, dimensions variable among species.
Notes.
Funiliomyces was first described by Aptroot (2004) as a monospecific genus, containing only its type species F. biseptatuss. With respect to ten species previously assigned to Dactylaria, our phylogenetic evidence shows they are phylogenetically distant from the type species of the genus (Dactylaria purpurella) (Fig. 1). However, these ten species cluster with Funiliomyces biseptatus, forming a distinct, well-supported lineage (Fig. 2). Therefore, in accordance with the “One Fungus, One Name” principle, we transfer these ten species to Funiliomyces and introduce the new species F. jiangxiensis within this genus. This emendation expands the genus to encompass both sexual and asexual morphs. The type species, F. biseptatus, represents the sexual morph, characterized by torpedo-shaped ascospores with two nearly central septa and appendages. In contrast, the eleven other species represent the asexual morphs, producing hyaline, septate conidiophores with sympodial, denticulate conidiogenous cells and solitary, hyaline, clavate or fusoid-ellipsoid conidia.
Funiliomyces
jiangxiensis
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
L.X. Mi, K.D. Hyde, H.Y. Song & D.M. Hu sp. nov
C4E7F4E0-6D94-56B2-B84C-E02A364C46B2
Index Fungorum: IF904530
Facesoffungi Number: FoF18898
Etymology.
The name refers to the place where the fungal was collected.
Funiliomyces jiangxiensis sp. nov. (HFJAU10125, holotype) a, b Colonies on the front and back of PDA medium (for 5 days); c Hyphae and sporulation structures; d, e Conidiophores bearing a conidium initial on one of its denticles; f, g Conidiophores; h–l Conidia. Scale bars: 10 μm (c, d); 5 μm (e); 10 μm (f, g); 20 μm (h); 10 μm (i–l).
Holotype.
HFJAU10125.
Description.
Endophytic fungus isolated from the roots of Tetradium ruticarpum. Sexual morph: Undetermined. Asexual morph: Mycelium consisted of hyaline, smooth, branched, septate, 1.7–2.6 µm diam hyphae. Conidiophores (6–)8.5–17.5(–23.5) × 1.8–3.5(–4) µm (mean ± SD = 12.5 ± 4.3 × 3 ± 0.4 µm, n = 35), macronematous, mononematous, hyaline, subcylindrical, arising from terminal or intercalary parts of aerial hyphae, mostly reduced to conidiogenous cells (rarely with a supporting cell), tapering towards the base, apex polyblastic, sympodial, inflated or geniculous-sinuous, with conspicuous, cylindrical denticles, up to 0.9 µm wide. Conidia 20–40 × 1.5–3.5 μm (mean ± SD = 27.8 ± 3.9 × 2.6 ± 0.3 µm, n = 40), hyaline, narrowly fusoid-ellipsoid, 0–3-septate, guttulate, apex sub-obtuse, base truncate.
Culture characteristics.
Colonies on PDA at 25 °C for 5 days, convex, white with cream margin, reverse pale brown with cream margin, no pigment in agar.
Material examined.
CHINA, Jiangxi Province, Ganzhou City, Ganxian District, 25.6121°N, 115.1211°E 412.9 m asl, isolated from healthy roots of Tetradium ruticarpum, 26 June 2022, Lixue Mi, dry culture HFJAU10125 (holotype); ex-type JAUCC 5298 = CCTCC AF 2023034; ibid., Yichun City, Zhangshu county-level city, 27.9931°N, 115.2123°E, 46 m asl, 21 September 2021, Lixue Mi, dry culture HFJAU10124; living culture: JAUCC 4255 = CCTCC AF 2023033.
Notes.
In the multi-gene analysis, Funiliomyces jiangxiensis (strains CCTCC AF 2023033 and CCTCC AF 2023034) forms a distinct lineage that groups with F. hwasunensis (≡ Dactylaria hwasunensis) and F. calliandrae (≡ D. calliandrae) as a sister branch, supported by high statistical values in the phylogenetic tree (SH-aLRT/UFB/BPP = 98.8/100/0.98). Morphologically, Funiliomyces jiangxiensis can be distinguished from F. hwasunensis by their conidiogenous cells and conidia. Specifically, Funiliomyces jiangxiensis develops distinct denticles on its conidiogenous cells, a feature that is absent in F. hwasunensis. Additionally, there is a notable difference in the number of septa in their conidia: Funiliomyces jiangxiensis has conidia with 0–3 septa, whereas F. hwasunensis produces conidia with more septa, ranging from 1 to 5. In terms of morphology, F. jiangxiensis resembles F. calliandrae but can be distinguished from it. Both have denticles on conidiogenous cells, yet those of F. jiangxiensis (0.9 μm) are shorter than F. calliandrae’s (1–3 μm). Additionally, the conidia of F. jiangxiensis (20–40 μm) are shorter than those of F. calliandrae [(37–)40–45(–47) μm]. The species was isolated from the roots of Tetradium ruticarpum (medicinal plant), which provides a basis for further research on its potential functional properties.
Other species in Funiliomyces
Funiliomyces
acaciae
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(Crous) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
F81157E9-1057-56E8-A44A-9566BD4A5A14
Index Fungorum: IF819073
Basionym.
Dactylaria acaciae Crous, Persoonia 37: 321 (2016).
Holotype.
CBS H-22876.
Type information.
USA, Hawaii, Oahu, on leaves of Acacia koa (Fabaceae), 30 September 2015, J.J. Le Roux (holotype CBS H-22876, culture ex-type CPC 29771 = CBS 142087).
Description.
See the original description in D’Souza et al. (2002) on page 141.
Funiliomyces
bisepatus
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(Matsushima) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
35065E27-80F5-5BFF-A470-B6A5F948E900
Index Fungorum: IF312614
Basionym.
Dactylaria biseptata Matsushima, Icones Microfungorum a Matsushima lectorum: 48 (1975).
Holotype.
MFC-4029.
Type information.
Japan, Ohdaigahara, Nara Pref, on a rotten leaf of Rhododendron metternichii (Ericaceae), July 1970, MFC-4029 (holotype).
Description.
See the original description in Matsushima (1975) on page 48–49.
Funiliomyces
calliandrae
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(Crous) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
49F7F1F4-0A9A-57DD-8C14-DADE48E5762E
Index Fungorum: IF859210
Basionym.
Dactylaria calliandrae Crous et al. Persoonia 54: 376–377 (2025).
Holotype.
CBS H-25715.
Type information.
Brazil, Minas Gerais, Viçosa, Clonar nursery, on living leaf of Calliandra tweediei (Fabaceae), 25 February, 2024, P.W. Crous, HPC 4399 (holotype CBS H-25715; culture ex-type COAD 3994 = CPC 48004).
Description.
See the original description in Crous et al. (2025) on page 376–377.
Funiliomyces
fragilis
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(de Hoog) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
BEA3A81B-7FCA-508A-BFFB-06B2C5C603EE
Index Fungorum: IF104169
Basionym.
Dactylaria fragilis de Hoog, Studies in Mycology 26: 30 (1985).
Holotype.
No.6074(CBS).
Type information.
The Netherlands, Opsterland, Oldeterp, on cupules of Fagus sylvatica (Fagaceae), H.A. van der Aa, October, 1977.
Description.
See the original description in de Hoog and van Oorscho (1985) on page 30.
Funiliomyces
hwasunensis
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(H.F. Liu & H.K. Sang) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
0E0930FD-B913-5AEF-BDA3-199EF4DB63F0
Index Fungorum: IF857258
Basionym.
Dactylaria hwasunensis H.F. Liu & H.K, IMA Fungus 16(e138479): 10 (2025).
Holotype.
CMML 20-35H.
Type information.
Korea, South Jeolla Province, Hwasun, isolated from roots of Zoysia japonica (Poaceae), October 2020, H. Liu & H. Sang, holotype CMML 20-35H, ex-holotype CMML 20-35, ex-isotype CMML 20-88.
Description.
See the original description in Liu et al. (2025) on page 12–14.
Funiliomyces
mavisleverae
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(Y.P. Tan, Bishop-Hurley & Marney) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
C0E4BDBC-B516-562F-AB90-5A208DABF199
Index Fungorum: IF902836
Basionym.
Dactylaria mavisleverae Y.P. Tan, Bishop-Hurley & Marney, Index of Australian Fungi 46: 3 (2024).
Holotype.
BRIP 76362a.
Type information.
Australia, Queensland, Brisbane, phylloplane of unidentified ornamental plant, January, 2024, T.S. Marney, BRIP 76362a (holotype).
Description.
See the original description in Tan et al. (2024) on page 3–4.
Funiliomyces
monticola
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(R. F. Castañeda & W. B. Kendr.) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
7E02CE3E-BF51-5EC0-A89D-F02181465D82
Index Fungorum: IF361523
Basionym.
Dactylaria monticola R.F. Castañeda & W.B. Kendr, University of Waterloo Biology Series, 35: 30 (1991).
Holotype.
INIFAT C 91/82.
Type information.
Cuba, Granma, Buey Arriba, La Estrella, on dead leaves of Andira inermis (Leguminosae), R.F. Castañeda, 14 March 1991.
Description.
See the original description in Castañeda and Kendr (1991) on page 30.
Funiliomyces
retrophylli
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(Crous) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
D277145E-E22D-5B20-8EEC-CAA7314CC75B
Index Fungorum: IF844283
Basionym.
Dactylaria retrophylli Crous, Fungal Systematics and Evolution 10: 41 (2022).
Holotype.
HPC 3260.
Type information.
Colombia, Finca El Cedral, on leaves of Retrophyllum rospigliosii (Podocarpaceae), M.J. Wingfield, February, 2020, HPC 3260 (holotype CBS H-24817, culture ex-type CPC 39510 = CBS 148271).
Description.
See the original description in Crous et al. (2022) on page 41–42.
Funiliomyces
sparsus
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(R. F. Castañeda & W. B. Kendr.) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
0F863F0C-362F-5604-BC23-01C7C748F899
Index Fungorum: IF361528
Basionym.
Dactylaria sparsa R.F. Castañeda & W.B. Kendr, University of Waterloo Biology Series. 35:33 (1991).
Holotype.
INIFAT C 91/68-2.
Type information.
Cuba, C. Habana, Santiago de las Vegas, on decaying leaves, R.F. Castañeda, 18 February 1990.
Description.
See the original description in Castañeda and Kendr (1991) on page 33.
Funiliomyces
zapatensis
Taxon classification
Animalia
AmphisphaerialesFuniliomycetaceae
(R.F. Castañeda) L.X. Mi, H.Y. Song, D.M. Hu & K.D. Hyde comb. nov.
8E847D83-3E72-582C-818E-5CFA62B73CD2
Index Fungorum: IF125340
Basionym.
Dactylaria zapatensis R.F. Castañeda, Fungi Cubenses III (La Habana): 5 (1988)
Holotype.
INIFAT C85/98
Type information.
Cuba, Matanzas, Ciénaga de Zapata, on fallen leaves of Nectandra coriacea (Lauraceae), R.F. Castañeda Ruiz, 26 May 1985.
Description.
See the original description in Castañeda Ruiz (1988) on page 5.
Funiliomyces currently includes only a single sexual morph species, F. biseptatus; therefore, we summarized the asexual morphological features, lifestyle, host associations, and distribution for all species (Table 4) to facilitate comparison within the genus.
Discussion
Previous studies have consistently demonstrated that Dactylaria is polyphyletic, yet the taxonomic placement of its type species, D. purpurella, has never been formally resolved. To date, there has been no phylogenetic study specifically designed to address the classification of D. purpurella; rather, it has generally been included only as part of broader datasets, without being treated as a focal taxon. As a result, its placement has remained ambiguous. For instance, Bussaban et al. (2005) showed that D. purpurella clustered with Ochroconis rather than other Dactylaria species based on ITS sequence data. Similarly, Réblová (2009) using LSU sequence data, demonstrated that D. purpurella, D. monticola, and D. parvispora fall into distinct clades, with D. purpurella forming an isolated lineage, while D. monticola grouping within Xylariales, and D. parvispora clustering with Papulosa amerospora in Papulosaceae. Subsequent studies further added to this complexity, placing D. purpurella within Magnaporthales (Klaubauf et al. 2014) or Venturiales (Lin et al. 2021). Taken together, these conflicting results highlight not only the unstable phylogenetic position of D. purpurella, but also the broader taxonomic challenges in delimiting generic boundaries within Dactylaria. In our study, the LSU-based phylogenetic analysis of Pezizomycotina demonstrated that Dactylaria is polyphyletic, with its species distributed across multiple classes, predominantly within Sordariomycetes. However, the phylogenetic placement of four species, D. purpurella (the type species), D. humicola, D. dimorpha, and D. dimorphospora, has not been assigned to any of the known classes within Pezizomycotina and thus remains unresolved (Fig. 1). As the first systematic investigation of the phylogenetic position of Dactylaria, based on all available LSU sequences data, our work provides a comprehensive overview of its taxonomic placement. The results emphasize the necessity of additional sampling and multi-locus phylogenetic analyses to resolve the taxonomy and evolutionary relationships within the genus, thereby laying the foundation for future refinement of its classification.
Although Dactylaria is polyphyletic, new species have continued to be introduced into the genus in recent years (Crous et al. 2016, 2022, 2025), all of which are placed within Amphisphaeriales, forming a single unresolved lineage. Fortunately, we also collected two new strains that cluster with the recently published Dactylaria species. Phylogenetic analysis based on the combined LSU, ITS, and RPB2 loci reveals that ten previously described “Dactylaria” species, our newly obtained isolates, and Funiliomyces biseptatus (type strain CBS 100373) cluster together into a single clade within Amphisphaeriales (Fig. 2). Topologically, this clade is comparable to lineages recovered in multiple previous studies, which remained unplaced within any established family of Amphisphaeriales (Crous et al. 2016, 2022, 2025). With the discovery of additional species of Nothodactylariaceae (Zhang et al. 2025) and their incorporation into the phylogenetic analyses of this study, this well-distinguished clade is recovered as a sister lineage to Nothodactylaria, the sole genus currently recognized in Nothodactylariaceae, with strong phylogenetic support (SH-aLRT/UFB/BPP = 99.9/94/0.99). In addition, multi-locus genetic distance analyses (ITS, LSU, and RPB2) revealed substantial divergence between this clade and other families within Amphisphaeriales, further supporting its independence from previously recognized families. Based on our results (Fig. 2, Table 3), both the phylogenetic analyses and genetic distance divergence clearly support that this distinct lineage represents a family-level taxon within Amphisphaeriales. Therefore, we establish the new family Funiliomycetaceae within Amphisphaeriales to accommodate this distinct lineage.
Morphologically, Funiliomycetaceae integrates both sexual and asexual morphs. Funiliomyces biseptatus, the only species with a sexual morph, produces torpedo-shaped ascospores with two nearly central septa and appendages, while the eleven other species represent asexual morphs, producing hyaline, septate conidiophores with sympodial, denticulate conidiogenous cells and solitary, hyaline, clavate or fusoid-ellipsoid conidia. This integration of sexual and asexual morphs within a single lineage highlights the evolutionary significance of the family and provides a robust morphological framework for family-level delimitation. When compared with Nothodactylariaceae, morphological evidence corroborates the close relationship between the two families, yet the overall conidial range in Funiliomycetaceae is considerably broader, encompassing species with distinctly larger conidia (Table 4). A more definitive diagnostic feature is the diversity of conidiogenous cells, which bear large cylindrical to geniculate denticles or lack denticles entirely, in sharp contrast to the uniform, pimple-like denticles of Nothodactylariaceae (Crous et al. 2019b, Zhang et al. 2025). Future studies will require additional species sampling and isolation to uncover sexual morphs, explore ecological adaptations, and expand molecular datasets for a more comprehensive understanding of Funiliomycetaceae.
Species within Funiliomycetaceae exhibit considerable ecological diversity in lifestyle, host range, and geographical distribution. The family includes endophytic, saprobic, and epiphytic species, colonizing hosts from a broad spectrum of plant families such as Fabaceae, Ericaceae, Poaceae, and Rutaceae, as well as decaying unidentified leaves or ornamentals, indicating no strict host specificity. Geographically, Funiliomycetaceae species are widely distributed across multiple continents, including North America, Asia, and South America, and occur in diverse climatic zones ranging from temperate to tropical regions (Table 4). By contrast, most Nothodactylariaceae species are saprobic, with the exception of Nothodactylaria nephrolepidis, which is epiphytic. They show a strong preference for ferns (e.g., Blechnaceae, Lomariopsidaceae) and are known only from China and South Africa, mostly in tropical and subtropical climates (Zhang et al. 2025). These marked differences in host range, ecological strategy, geographical distribution, and climatic preference underscore the distinct ecological niche occupied by Funiliomycetaceae. With the addition of Funiliomycetaceae, the order now comprises 21 families, highlighting both the considerable diversity within Amphisphaeriales and the presence of previously unrecognized lineages.
The new family comprises Funiliomyces biseptatus, ten previously described “Dactylaria” species, and our newly obtained isolates. As confirmed by phylogenetic analyses (Fig. 1), these ten “Dactylaria” species are not congeneric with the type of Dactylaria (D. purpurella) but form a coherent, monophyletic group with F. biseptatus. This supports their recognition as a single genus, consistent with the “One Fungus, One Name” principle (Hawksworth 2012) and the general taxonomic guideline of recognizing monophyletic genera with coherent morphological characters. Accordingly, we transfer the ten previously described “Dactylaria” species into the genus Funiliomyces as new combinations (F. acaciae, F. bisepatus, F. calliandrae, F. fragilis, F. hwasunensis, F. mavisleverae, F. monticola, F. retrophylli, F. sparsus, and F. zapatensis). Furthermore, Funiliomyces is designated as the type genus of Funiliomycetaceae. The genus was described by Aptroot (2004), containing only its type species, F. biseptatus, which was collected from dead leaves of a Bromeliaceae species in a rock field. This species differs from the other newly transferred species in the genus by possessing only a sexual morph. In the present study, we summarize the asexual morphological features, lifestyle, host associations, and distribution of Funiliomyces species (Table 4), providing a comprehensive framework for future identification, ecological studies, and taxonomic research of the genus.
In addition to transferring ten “Dactylaria” species to Funiliomyces, we describe a new species (F. jiangxiensis) that further enriches the diversity of this genus. Multi-gene phylogenetic analyses also indicate that the new isolates, Funiliomyces jiangxiensis, occupies a distinct position within Funiliomyces, supported by high statistical values (SH-aLRT/UFB/PP = 98.8/100/0.90). Morphologically, F. jiangxiensis can be distinguished from its closely related species (F. hwasunensis and F. calliandrae) by differences in conidiophore structure, conidiogenous cell denticulation, and conidial characteristics (Table 4). Additionally, Funiliomyces jiangxiensis was isolated from the medicinal plant Tetradium ruticarpum. This plant, commonly known as Evodia, is renowned for its therapeutic properties, including anti-inflammatory, analgesic, and antimicrobial activities (Xiao et al. 2023). Endophytes have been studied and are known to produce novel biologically active compounds (Liao et al. 2025). For example, Funiliomyces hwasunensis CMML 20-35, isolated from the roots of Zoysia japonica, inhibited the mycelial growth of Rhizoctonia solani AG2-2(IIIB) by 37.97% in dual culture (Liu et al. 2025). To date, endophytic fungal studies on this host are limited, with only a few taxa reported recently, including Cyphellophora guangxiensis and Pseudokeissleriella tetradii (Mi et al. 2025a, 2025b). These findings emphasize the need for comprehensive surveys and further functional studies to explore the diversity and antimicrobial capabilities of endophytic fungi associated with this medicinal plant.
Supplementary Material
XML Treatment for Funiliomycetaceae
XML Treatment for Funiliomyces
XML Treatment for Funiliomyces jiangxiensis
XML Treatment for Funiliomyces acaciae
XML Treatment for Funiliomyces bisepatus
XML Treatment for Funiliomyces calliandrae
XML Treatment for Funiliomyces fragilis
XML Treatment for Funiliomyces hwasunensis
XML Treatment for Funiliomyces mavisleverae
XML Treatment for Funiliomyces monticola
XML Treatment for Funiliomyces retrophylli
XML Treatment for Funiliomyces sparsus
XML Treatment for Funiliomyces zapatensis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barbosa FR, Raja HA, Shearer CA et al. (2013) Some freshwater fungi from the Brazilian semi-arid region, including two new species of hyphomycetes. Cryptogamie, Mycologie 34(3): 243–258. 10.7872/crym.v 34.iss 2.2013.243 · doi ↗
- 2Becerra-Hernández CI, González D, De Luna E et al. (2016) First report of pleoanamorphy in Gyrothrix verticiclada with an Idriella-like synanamorph. Cryptogamie, Mycologie 37(2): 241–252. 10.7872/crym/v 37.iss 2.2016.241 · doi ↗
- 3Beimforde C, Schmidt AR, Rikkinen J et al. (2020) Sareomycetes cl. nov.: A new proposal for placement of the resinicolous genus Sarea (Ascomycota, Pezizomycotina). Fungal Systematics and Evolution 6: 25–37. 10.3114/fuse.2020.06.02PMC 745177632904095 · doi ↗ · pubmed ↗
- 4Bhatt G, Kendrick B (2011) The generic concept of Diplorhinotrichum and Dactylaria, and a new species of Dactylaria from soil. Canadian Journal of Botany 46: 1253–1257. 10.1139/b 68-167 · doi ↗
- 5Bhattacharya D, Lutzoni F, Reeb V et al. (2000) Widespread occurrence of spliceosomal introns in the r DNA genes of ascomycetes. Molecular Biology and Evolution 17(12): 1971–1984. 10.1093/oxfordjournals.molbev.a 02629811110913 · doi ↗ · pubmed ↗
- 6Bussaban B, Lumyong S, Lumyong P et al. (2005) Molecular and morphological characterization of Pyricularia and allied genera. Mycologia 97: 1002–1011. 10.3852/mycologia.97.5.100216596952 · doi ↗ · pubmed ↗
- 7Castañeda Ruíz RF (1988) Fungi cubenses III. Instituto de Investigaciones Fundamentales en Agricultura Tropical “Alejandro de Humboldt”, 1–27. http://www.cybertruffle.org.uk/cyberliber/03416/cfo_.htm
- 8Chandra H, Yadav A, Prasad R et al. (2024) Fungal endophytes from medicinal plants acting as natural therapeutic reservoir. The Microbe 3: 100073. 10.1016/j.microb.2024.100073 · doi ↗