Role of Altered Metabolites and Metabolic Pathways in Major Tuber Crops Under Drought Stress
Maltase Mutanda, Fikile N. Makhubu, Sandiswa Figlan

TL;DR
This study explores how major tuber crops like cassava, potato, and sweet potato respond metabolically to drought stress, identifying key metabolites and pathways involved.
Contribution
The study provides a comprehensive synthesis of drought-induced metabolic responses in major tuber crops, highlighting gaps in cassava research.
Findings
Trehalose and proline are the most commonly studied and affected metabolites across cassava, potato, and sweet potato under drought stress.
Potato and sweet potato have more drought-responsive metabolites due to higher metabolomics research, while cassava lacks sufficient data.
Key enriched pathways include glyoxylate metabolism, citrate cycle, and starch and sucrose metabolism.
Abstract
Drought stress poses a significant challenge to growth and productivity of major tuber crops, particularly cassava ( Manihot esculenta Crantz), potato ( Solanum tuberosum L.) and sweet potato ( Ipomoea batatas (L.) Lam.). These crops are among the most widely cultivated tubers globally and play a critical role in food and nutritional security, especially in drought‐prone regions of sub‐Saharan Africa, Asia and Latin America. Several studies have highlighted that metabolites such as sucrose, proline and arginine contribute to osmotic adjustment, cellular protection and energy balance under drought stressed conditions. However, a comprehensive synthesis of drought‐induced metabolic responses and associated pathways utilized by major tuber crops remains limited. Therefore, this study aimed to identify and evaluate the metabolic responses and pathways altered under drought stress in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
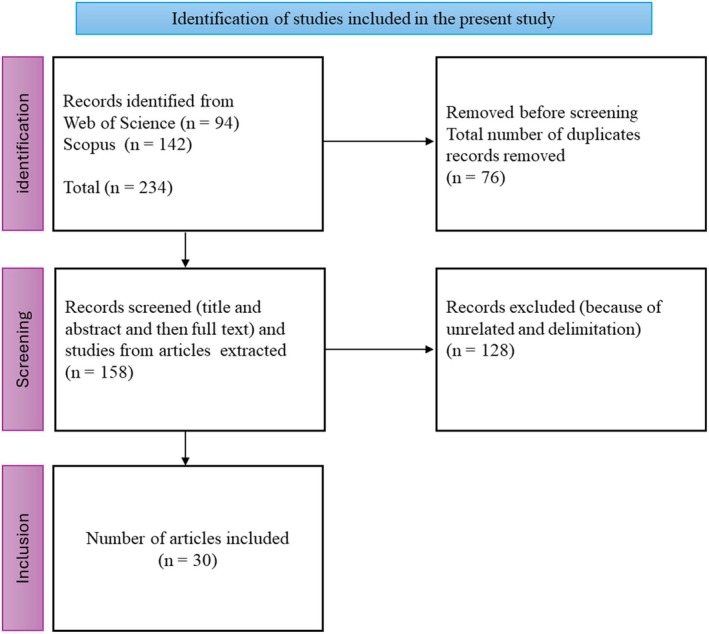 FIGURE 1
FIGURE 1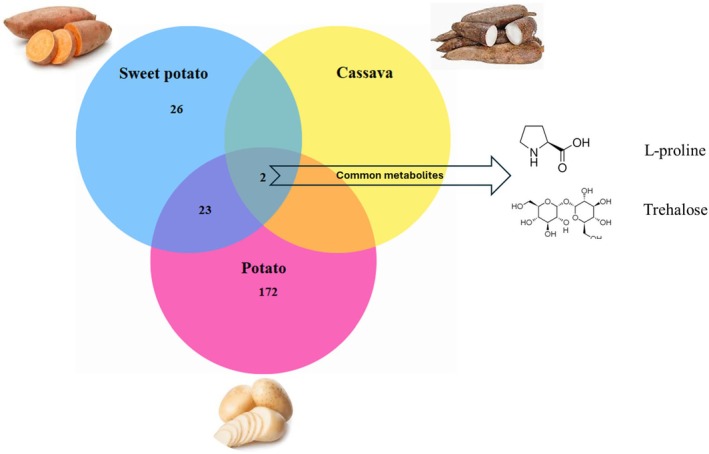 FIGURE 2
FIGURE 2| Pathway name | Assigned metabolites |
| −log( | Holm | FDR | Impact |
|---|---|---|---|---|---|---|
| Glyoxylate and dicarboxylate metabolism | Malate, oxaloacetate, cis‐aconitate, isocitrate, citrate, glutamate, serine, succinate, glycine, 2‐axo‐glutarate, glycerate | 1.37 × 10−8 | 7.86 | 1.25 × 10−6 | 1.20 × 10−6 | 0.48 |
| Citrate cycle (TCA cycle) | Malate, succinate, oxaloacetate, fumarate, citrate, Phosphoenolpyruvate, cis‐aconitate, iso‐citrate, 2‐axo‐glutarate, pyruvate | 2.64 × 10−8 | 7.58 | 2.37 × 10−6 | 1.20 × 10−6 | 0.45 |
| Alanine, aspartate, and glutamate metabolism | Aspartate, Alanine, oxaloacetate, asparagine, pyruvate, 2‐axo‐glutarate, fumarate, glutamate, 4‐aminobutanoate, succinate | 8.39 × 10−8 | 7.08 | 7.47 × 10−6 | 2.55 × 10−6 | 0.70 |
| Galactose metabolism | Stachyose, raffinose, myo‐Inositol, melibiose, sucrose, glucose, fructose, glycerol, mannose, Galactinol | 8.63 × 10−7 | 6.06 | 7.60 × 10−5 | 1.96 × 10−5 | 0.35 |
| Starch and sucrose metabolism | Fructose, starch, sucrose, maltose, cellobiose, glucose, glucose‐6‐phosphate, fructose‐6‐phosphate, alpha‐trehalose | 1.19 × 10−6 | 5.92 | 1.04 × 10−4 | 2.17 × 10−5 | 0.73 |
- —Water Research Commission10.13039/501100004424
- —University of South Africa10.13039/501100008227
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCassava research and cyanide · Potato Plant Research · Plant responses to water stress
Introduction
1
Tuber crops are ranked third among food crops based on global production after cereal and legume crops (Chauhan et al. 2022). They form a substantial part of the world's food supply, contributing significantly to the nutritional needs of populations globally (Chandrasekara and Josheph Kumar 2016). Tuber crops, including potato ( Solanum tuberosum ), cassava ( Manihot esculenta ) and sweet potato ( Ipomoea batatas L.) are widely cultivated due to their ability to thrive in diverse environmental conditions, producing optimal yields even in suboptimal climates (Nanbol and Namo 2019). Despite their significance and resilience, tuber crops are still vulnerable to the adverse effects of drought stress, which can severely impact their growth and yield (Begna 2021; Makhubu et al. 2025; Mutanda, Amelework, et al. 2025). Under drought‐stressed conditions, plants develop a range of strategies of adaptations at different levels of organization including morphological, physiological, biochemical and metabolic adjustments to cope with water scarcity (Shah and Satti 2023; Khamis et al. 2025; Mutanda, Shimelis, et al. 2025). Therefore, understanding these mechanisms at the metabolic and biochemical levels is crucial for developing drought‐tolerant varieties that can maintain high productivity under limited water conditions.
Building on their inherent adaptability and resilience, recent advances in plant metabolomics have significantly expanded scientists' understanding of how tuber crops respond to drought stress conditions. Unlike genomics and transcriptomics, which mainly analyze gene expression patterns, metabolomics offers a direct snapshot of the plant's physiological state under drought‐stressed conditions (Almeida et al. 2020; Qiu et al. 2023). Metabolomics, which involves the comprehensive study of metabolites, provides valuable insights into the molecular mechanisms that drive drought response. This approach focuses on analyzing the metabolites such as sugars, amino acids, organic acids and secondary metabolites (such as phenolic acids and flavonoids). The changes in these metabolites can influence critical processes, including osmotic regulation and maintenance of cellular integrity (Mutanda, Amelework, et al. 2025; Mutanda, Shimelis, et al. 2025). However, specific directional changes (e.g., accumulation of osmolytes or antioxidants) are associated with enhanced drought tolerance (Arbona et al. 2013; Drapal et al. 2017), but the effect depends on tissue, timing and overall plant physiological. Importantly, drought‐induced shifts in metabolite levels can indicate either adaptive processes or stress‐related damage such as increased malondialdehyde (MDA) showing lipid peroxidation and membrane injury (Zhang, Yan, et al. 2021; Zhang, Luan, et al. 2021).
Tuber crops' metabolomics has proven instrumental in identifying and characterizing a diverse array of metabolites that contribute to plant health and resilience (Evers et al. 2010; Barnaby et al. 2015; Yang et al. 2015; Drapal et al. 2017; Zhou et al. 2022). The application of metabolomics in drought research has provided deeper insights into how tuber crops regulate their metabolic networks to cope with water scarcity. Furthermore, other studies have shown that tuber crops exhibit shifts in carbohydrate metabolism, alterations in organic acid and nitrogen metabolism, and the activation of pathways that help maintain energy balance during periods of water scarcity (Rosado‐Souza et al. 2019; Obata et al. 2020; Zhou et al. 2022; Yin, Chen, et al. 2024; Yin, Qiao, et al. 2024). These metabolic adjustments enhance the plant's ability to conserve water, sustain cellular functions, and maintain growth under drought‐stressed conditions.
Furthermore, drought stress triggers the production of reactive oxygen species (ROS), which can damage vital cellular components leading to oxidative stress and potential cellular dysfunction (Cruz de Carvalho 2008; Mutanda, Amelework, et al. 2025; Mutanda, Shimelis, et al. 2025). However, plants including tuber crops use antioxidant defense mechanisms to mitigate ROS‐induced damage. These defenses include the accumulation of metabolites such as sucrose, proline, ascorbate and glutathione, which help maintain cellular turgor and protect cellular membranes from dehydration under water stress (Sachdev et al. 2021). Notably, proline, a well‐known osmolyte, plays a crucial role in stabilizing proteins and cellular structures while also acting as an antioxidant to scavenge ROS (Ghosh et al. 2022). The accumulation of proline and other compatible solutes helps preserve cellular membrane integrity and sustain essential physiological functions under drought‐stressed conditions (Mutanda et al. 2024). Therefore, these metabolite‐driven defense strategies enable tuber crops to survive and maintain productivity in drought‐prone environments.
Despite these significant advances in understanding the metabolic responses to drought stress in crops, substantial gaps remain in understanding the full spectrum of metabolic changes that occur under drought‐stressed conditions. While much is known about the role of specific metabolites associated with drought tolerance, the metabolic responses and pathways in major tuber crops (such as cassava, potato and sweet potato) are largely unexplored. The metabolic responses to drought stress are highly dynamic and can vary between different crops, genotype, developmental stage, tissue type, drought severity, duration and the method of drought stress imposition (Burnett et al. 2021; Makhubu et al. 2024; Mutanda et al. 2024; Khamis et al. 2025; Makhubu et al. 2025). This variability highlights the need for a more comparative approach to studying drought tolerance of tuber crops at their metabolic level. Therefore, this study aimed to evaluate the metabolic responses and pathways under drought stress in major tuber crops (cassava, potato and sweet potato). The findings will provide valuable insights to future breeding programs by identifying the key metabolites and pathways that could serve as species‐non‐specific biomarkers to enhance drought resilience and productivity of tuber crops for sustainable agriculture.
Materials and Methods
2
Literature Search
2.1
The Preferred Reporting Items for Systematic Reviews and Meta‐Analyses (PRISMA) guidelines were strictly followed in designing, conducting, and reporting this review. This systematic narrative review adhered to the PRISMA 2020 statement, ensuring transparent identification, screening, eligibility assessment and inclusion of studies. The PRISMA 2020 checklist and flow diagram (as recommended by Page et al. 2021) were used to guide the reporting, and because this review used published literature only, ethical approval was not required. A systematic search of literature was conducted from two bibliographic databases [Web of Science (https://www.webofscience.com/) and Scopus (https://www.scopus.com/)]. The search for articles from Google Scholar was avoided due to the high number of irrelevant results retrieved. The studies were reported in English by peer‐reviewed journals in the period between 2010 and 2024 (inclusive). The period from 2010 to 2024 ensures the inclusion of recent research, reflecting the latest advancements, methodologies and technologies in the field. This timeframe ensures the studies remain relevant to current scientific understanding and practices. The search on academic databases was performed on November 5, 2024. The keywords that facilitated the search of articles were “cassava”, “sweet potato”, “potato” “drought stress”, “limited water conditions”, “water stress”, “metabolite”, “metabolite profiling” and “metabolic responses”. We included peer‐reviewed research studies conducted under open‐field and controlled environmental conditions. The study selection procedure is summarized in the PRISMA flow diagram (Figure 1), and all publications included in the review as well as the metabolites altered by drought stress are listed in Table S1. The present study only included metabolites that showed statistically significant alterations under drought‐stressed compared to non‐stress conditions in major tuber crops. For pathway analysis, metabolite names reported across the selected studies were standardized by matching them to their corresponding entries in the Human Metabolome Database (HMDB, https://hmdb.ca/ accessed January 20, 2025) and the Kyoto Encyclopedia of Genes and Genomes (KEGG, https://www.genome.jp/kegg/ accessed January 20, 2025) using default database settings. The list of metabolites was then imported into MetaboAnalyst 5.0 for pathway enrichment and topology analysis. The pathway enrichment was performed using Fisher's exact test, and pathway impact was assessed using relative‐betweenness centrality, following the default parameters in MetaboAnalyst. All steps were performed using default settings.
Articles selection process for the present study.
Results
3
Literature Search
3.1
After checking the relevance according to the titles, the initial search identified a total of 234 studies published from 2010 to 2024. A detailed review of titles, abstracts and full text according to the selection criteria resulted in a total of 30 peer‐reviewed research articles reporting original experimental data (reviews, editorials, and book chapters were excluded), and all of them were reported in English from all continents. There was a drastic increase in publications from 2010 to 2024, with many of them focusing on potato drought stress responses. A total of 223 metabolites were identified as being influenced by drought stress and they were extracted from a total of 30 peer‐reviewed research articles. Of these, 25 metabolites were found in two or more crops and reported in more than three studies each. Notably, two metabolites (L‐proline and trehalose) exhibited significant changes under drought stress responses across three crops reviewed in the present study (Figure 2). Table S1 provides an overview of the selected studies and metabolites significantly altered under drought‐stressed conditions.
Venn diagram showing the significantly affected metabolites in three major tuber crops (sweet potato, cassava and potato) under drought‐stressed conditions. The numbers within each section represent the count of unique or shared metabolites among the three crops. Sweet potato exhibits 26 unique metabolites, cassava has no unique metabolite, and potato contains 172 distinct metabolites. Twenty‐three metabolites are shared between sweet potato and potato, while only two metabolites are common across all three crops. The common metabolites, depicted with their chemical structures, include proline and trehalose based on screened studies.
Metabolic Responses
3.2
Table S1 summarizes the presence of various metabolites identified in cassava, potato, and sweet potato, along with supporting references. A diverse range of metabolites, including sugars, amino acids, organic acids and phenolic acids were identified across the three crops. Trehalose was found to be a common sugar in all three crops. Sweet potato exhibited a unique profile with the presence of xylose, which was not detected in cassava or potato. On the other hand, raffinose was identified in potato plants only. Potato exhibited the most extensive metabolite profile, with a significantly larger number of unique metabolites detected compared to cassava and sweet potato. Sweet potato showed a distinct metabolite composition, characterized by the presence of several compounds not found in cassava or potato, highlighting specialized metabolic pathways.
Pathway Analysis
3.3
The present analysis revealed the involvement of multiple metabolic pathways associated with drought stress adaptation in tuber crops which are highlighted in Table 1 and Figure S1. Glyoxylate and dicarboxylate metabolism (GDM) was highly enriched, with key metabolites including malate, oxaloacetate, citrate and succinate, indicating its role in carbon assimilation and energy conservation. The citrate cycle (TCA cycle) was also significantly represented, with malate, succinate, oxaloacetate, fumarate and citrate detected. The identification of phosphoenolpyruvate and pyruvate suggests an active link between glycolysis and the TCA cycle, supporting carbon flux regulation. The alanine, aspartate, and glutamate metabolism pathway showed enrichment with aspartate, alanine, oxaloacetate, asparagine and 2‐oxo‐glutarate. The carbohydrate metabolism pathways were prominently represented, with galactose metabolism showing the accumulation of stachyose, raffinose, sucrose, glucose and myo‐inositol; as well as starch and sucrose metabolism enriched with glucose, fructose, maltose, starch and trehalose.
TABLE 1: Significant metabolic pathways (with an impact value > 0.1 and p value < 0.05) detected in major tuber crops evaluated in the present study.
Discussion
4
Metabolic Changes in Tuber Crops Under Drought Stress
4.1
Tuber crops exhibit a wide range of metabolic responses to drought stress, with numerous studies providing valuable insights into these adaptations. In the current study, 223 metabolites were identified as being influenced by drought stress in cassava, potato and sweet potato. Some metabolites demonstrated conserved responses across the three crops, while others exhibited species‐specific changes. Notably, two metabolites (L‐proline and trehalose) were consistently reported as responsive to drought stress across the three crops reviewed in the present study.
Furthermore, the studies reviewed indicate that the metabolites accumulated in these tuber crops contribute to drought‐tolerance mechanisms, including osmotic adjustment (Fang and Xiong 2015; Zia et al. 2021). This phenomenon enables cells to maintain positive turgor pressure despite the water deficit (Zivcak et al. 2016). Osmotic adjustment involves the accumulation of osmolytes that stabilize cell structures, maintain osmotic balance, and mitigate the effects of stress (Fang and Xiong 2015; Guru et al. 2025). These osmolytes include a variety of sugars (such as glucose, fructose, sucrose and trehalose); amino acids (such as proline and gamma‐aminobutyric acid); and polyamines (such as putrescine and spermidine) (Fang and Xiong 2015). Therefore, it is important to note that metabolites also play additional roles beyond osmotic adjustment, such as antioxidant defense, signaling, energy metabolism, and membrane stabilization (Mutanda et al. 2024).
Sugars
4.1.1
Sugars are among the most important osmolytes, serving dual roles as osmoprotectants and signaling molecules (Dutta et al. 2019; Ejaz et al. 2020). The comparative metabolomic analysis of cassava, potato and sweet potato revealed distinct sugars involved in plant's survival under drought‐stressed conditions (Rosa et al. 2009). The consistent detection of trehalose across all three crops suggest that it is important in their physiology. Trehalose is a well‐known stress protectant, stabilizing proteins and membranes under drought‐stressed conditions (Evers et al. 2010; Barnaby et al. 2015; Yang et al. 2015; Ren et al. 2017; Zhou et al. 2022). This suggests that all three tuber crops utilize trehalose as a core mechanism to mitigate the impact of drought stress conditions. The relatively high levels of trehalose previously observed in drought‐tolerant cassava varieties further support this hypothesis (Ren et al. 2017; Li et al. 2022). The presence of melibiose, stachyose and raffinose, which are raffinose family oligosaccharides (RFOs) involved in drought tolerance (Evers et al. 2010; Juhász et al. 2014; Yang et al. 2015; Sprenger et al. 2018; Barnaby et al. 2019), suggests that potato plants might utilize these sugars to enhance their resilience to drought stress conditions. Furthermore, the identification of maltose and starch aligns with the role of potato tubers as a major starch storage organ, with maltose being a key intermediate in starch degradation during drought stress. These findings are consistent with previous research demonstrating significant fluctuations in sugar content during potato tuber development and storage (Kumar 2011; Haider et al. 2023). Furthermore, the unique presence of lyxose, gentiobiose and erythrose in sweet potato points to distinct sugar metabolism pathways compared to cassava and potato. While the specific functions of lyxose and gentiobiose in sweet potato could be linked to cell wall metabolism. The absence of many of the potato‐specific sugars in cassava and sweet potato suggests that these crops rely on alternative mechanisms for carbohydrate storage and stress adaptation. For instance, cassava primarily stores carbohydrates as starch but may utilize different types of oligosaccharides or polyols for stress protection. Further research is needed in breeding of genotypes with manipulated genes involved in trehalose or raffinose synthesis to enhance drought tolerance in potato or cassava.
Amino Acids
4.1.2
The consistent detection of proline in all three tuber crops highlights its crucial role in plant physiology, especially under drought stressed conditions (Evers et al. 2010; Yang et al. 2015; Barnaby et al. 2015; Sprenger et al. 2016; Drapal et al. 2017; Barnaby et al. 2019; Haas et al. 2020; Demirel et al. 2020; Da Ros et al. 2020; Toubiana et al. 2020; Yin, Chen, et al. 2024; Yin, Qiao, et al. 2024). This is because proline functions as an osmoprotectant which stabilizes proteins and membranes. The elevated levels of proline observed in drought‐stressed plants (Singh et al. 2015; Meena et al. 2019) further support its role in drought stress adaptation. Some studies reported a broad array of amino acids in potato, including arginine, cysteine, glutamate and lysine, highlighting their metabolic diversity under drought stress. These amino acids contribute to protein synthesis, enzyme activity and signaling processes during the period of drought stress (Ali et al. 2024). The presence of γ‐aminobutyric acid, a non‐protein amino acid with signaling functions, further enhances drought tolerance in potato cultivars (Juhász et al. 2014). The exclusive detection of L‐tyrosine, 3‐hydroxy‐L‐proline and alpha‐aminoadipic acid in sweet potato suggests distinct amino acid metabolism pathways compared to cassava and potato. The presence of these metabolites may be linked to the unique biochemical pathway enhancing the drought tolerance of sweet potato genotypes. The absence of many potato‐specific amino acids in cassava and sweet potato suggests that these crops may have lower protein content (Chandrasekara and Josheph Kumar 2016) or rely on different mechanisms for amino acid synthesis and regulation. Therefore, future research should focus on analyzing the amino acid profiles especially on different varieties of cassava to identify genetic factors that influence amino acid metabolism.
Phenolic Acids
4.1.3
Phenolic acids are a class of plant secondary metabolites with diverse biological activities, including antioxidant properties. They contribute to plant defense mechanisms against oxidative stress caused by drought‐stressed conditions. The presence of 3‐phenylacetic acid and 4‐vinylphenol dimer in sweet potato points to distinct phenolic acid metabolism pathways compared to cassava and potato (Zhou et al. 2022). The presence of p‐coumaric acid, caffeic acid, ferulic acid, and sinapic acid only in potato suggests that potato may have stronger antioxidant activities than other crops (Drapal et al. 2017). Drought stress induces the production of ROS, leading to oxidative damage in plant cells. The phenolic acids can act as antioxidants that can scavenge ROS and protect cellular components from oxidative stress (Drapal et al. 2017; Sprenger et al. 2016). The presence of compounds/metabolites like 3‐phenylacetic acid in sweet potato and caffeic acid, p‐coumaric acid, ferulic acid and sinapic acid in potato suggests an active antioxidant defense mechanism in these crops. Additionally, compounds like benzoic acid, 3‐hydroxybenzoic acid and 3,4‐dihydroxybenzoic acid found in potato further enhance ROS scavenging capacity. Therefore, the identification and characterization of the phenolic acids provide valuable insights into developing crops with improved antioxidant defense mechanisms.
Organic Acids
4.1.4
Organic acids are essential metabolites that play pivotal roles in plant's response to drought‐stressed conditions. In tuber crops, organic acids such as malic acid, citric acid and quinic acid accumulate under drought‐stressed conditions, contributing to osmotic adjustment and cellular homeostasis. These acids help maintain cell turgor pressure by balancing the osmotic potential within the cell, thereby preventing dehydration (Barnaby et al. 2015; Drapal et al. 2017; Haas et al. 2020). In potato, α‐ketoglutarate and quinic acid are particularly significant for their involvement in osmotic regulation, while in sweet potato, maleic acid and citraconic acid play vital roles in maintaining osmotic balance (Barnaby et al. 2015; Drapal et al. 2017; Barnaby et al. 2019; Haas et al. 2020). Beyond osmotic regulation, organic acids help buffer ion imbalances and stabilize cellular pH under drought‐stressed conditions, ensuring the optimal functioning of cellular processes (Barnaby et al. 2015; Drapal et al. 2017; Guo et al. 2018). They also serve as metabolic intermediates, acting as precursors for the biosynthesis of other essential metabolites. For instance, malic acid can be converted into sugars through gluconeogenesis, providing an alternative energy source during water scarcity (Barnaby et al. 2015; Drapal et al. 2017). In sweet potato, maleic acid and citraconic acid contribute to osmotic adjustment and modulate the activity of stress‐related enzymes, further enhancing drought tolerance (Barnaby et al. 2019; Haas et al. 2020). The presence of shared organic acids such as malic acid and citric acid in both potato and sweet potato indicates that these crops utilize common mechanisms to withstand drought stress conditions. This highlights the universal importance of organic acids in drought tolerance across different tuber crops (Barnaby et al. 2015; Drapal et al. 2017; Haas et al. 2020). Therefore, understanding the roles of organic acids and their biosynthetic pathways could provide valuable insights for breeding drought‐tolerant genotypes of cassava, potato and sweet potato.
Polyamines
4.1.5
Polyamines are low‐molecular‐weight aliphatic amines that play crucial roles in various plants' physiological processes, including responses to drought‐stressed conditions (Hasan et al. 2021). The metabolites under the polyamines group (including putrescine and spermidine) stabilize the phospholipids and proteins in the cell membrane (Kapoor 2023). In addition, these metabolites are considered to have antioxidant properties (Sprenger et al. 2016) that promote the activity of antioxidant enzymes to reduce oxidative stress (Weiss and Landauer 2000). This protective role helps maintain plant growth and productivity as well as regulating stomatal conductance to reduce transpiration rate. Furthermore, polyamines play a significant role in enhancing root system architecture, which is crucial for plants to effectively absorb water and nutrients when experiencing drought stress. Research has shown that the upregulation of polyamines such as putrescine and spermidine can enhance root length, biomass, and lateral root proliferation in plants (Couée et al. 2004; Tang and Newton 2005; Tyagi et al. 2023), leading to better soil moisture utilization. Therefore, plant breeders should focus on targeting and utilizing polyamines as biomarkers for improving tuber crop root morphology, especially in cassava.
Metabolic Pathways Utilized to Improve Drought Tolerance of Tuber Crops
4.2
Drought stress is a major environmental constraint affecting the growth, development, and yield of tuber crops such as cassava, potato and sweet potato. These crops have developed complex metabolic strategies to cope with water stress, ensuring both survival and productivity under drought conditions (Kapoor 2023). Key adaptive mechanisms involve regulation of metabolic pathways controlling osmotic balance and stress signaling (Wasaya et al. 2021; Lal et al. 2022). Therefore, understanding how these metabolic pathways function under drought stress is essential for improving the drought resilience of tuber crops. The following sections explore the specific metabolic pathways activated in response to drought stress, highlighting the key metabolites involved and their roles in enhancing drought tolerance.
Glyoxylate and Dicarboxylate Metabolism
4.2.1
The GDM pathway plays a critical role in plant survival under drought stress by facilitating energy conservation and carbon assimilation. It allows plants to use stored fatty acids as carbon sources when photosynthesis is compromised, producing essential metabolites for survival (Kunze et al. 2006; Wang, Li, and Dong 2024; Wang, Zhang, et al. 2024). This ability to generate energy from alternative sources supports plant survival and productivity, making the GDM pathway a key target for improving drought tolerance in crops. Furthermore, in plant metabolism, the glyoxylate and dicarboxylate pathway significantly upregulates the production of metabolites like malate and succinate. These metabolites act as osmolytes to maintain cell turgor and balance metabolic disruptions caused by reduced photosynthesis. The enzymes (isocitrate lyase and malate synthase) involved in this pathway are crucial for the conversion of acetyl‐CoA into succinate, a precursor for malate synthesis (Yuenyong et al. 2019) to maintain cell turgor under drought stress (Ahn et al. 2016). Therefore, these insights suggest that metabolic engineering of GDM could improve drought resilience and yield stability under water‐limited environments.
TCA Cycle
4.2.2
The TCA cycle plays a vital role in enhancing drought tolerance in plants by sustaining energy production and ensuring metabolic flexibility under drought‐stressed conditions (Li, Yang, et al. 2021; Li, Shao, et al. 2021). Limited water availability reduces photosynthesis and energy supply, but tuber crops can maintain ATP production by increasing TCA cycle flux. Key intermediates including malate and succinate support mitochondrial respiration, enabling continuous energy generation. Additionally, citrate regulates carbon flow and prevents metabolic imbalances that affect growth under drought stress (Tahjib‐Ul‐Arif et al. 2021). The elevated levels of iso‐citrate and 2‐oxo‐glutarate promote the synthesis of proline and glutamate (Gai et al. 2020), which help mitigate oxidative stress induced by drought‐related ROS. Moreover, the phosphoenolpyruvate and pyruvate nodes of the TCA cycle contribute to drought resilience (Diniz et al. 2020) by facilitating carbon partitioning and metabolic adjustments. Phosphoenolpyruvate supports carbon metabolism, aiding gluconeogenesis and energy redistribution (Shi et al. 2015; Hu et al. 2025), while pyruvate connects glycolysis to the TCA cycle (Zhang and Fernie 2023), ensuring a steady supply of carbon skeletons for energy production and biosynthesis under drought. Therefore, these findings highlight the potential for targeted metabolic interventions to optimize TCA cycle flux and support drought tolerance in tuber crops.
Alanine, Aspartate and Glutamate Metabolism
4.2.3
The metabolic pathway involving alanine, aspartate and glutamate is pivotal in bolstering drought tolerance in tuber crops by facilitating crucial physiological and biochemical adaptations. This pathway is central to nitrogen assimilation and redistribution processes that are indispensable for maintaining metabolic equilibrium under water‐limited conditions (Raza et al. 2025). In addition, these amino acids enhance the plant's capacity to adapt to drought stress by optimizing nitrogen utilization for protein synthesis and other vital metabolic processes. Notably, glutamate serves as a precursor for proline synthesis (Stolarz and Hanaka 2025), a key osmolyte that helps maintain cellular turgor pressure and protects proteins from denaturation under drought stress. Aspartate and alanine mitigate oxidative damage by supporting pathways linked to cellular homeostasis under drought conditions (Wang, Li, and Dong 2024; Wang, Zhang, et al. 2024). Furthermore, these amino acids act as signaling molecules, modulating stress responses and adaptive mechanisms in plants (Raza et al. 2024). Therefore, these amino acids reinforce plant drought tolerance through both biochemical and signaling roles.
Galactose Metabolism
4.2.4
Drought stress significantly activates the galactose metabolism in plants to enhance survival and productivity (Zhang, Yan, et al. 2021; Zhang, Luan, et al. 2021). Under drought stressed conditions, tuber crops accumulate metabolites including stachyose, raffinose and galactinol, which act as osmoprotectants to prevent oxidative damage (Nishizawa et al. 2008; Sanyal et al. 2023). The myo‐inositol is an important metabolite in this pathway, which enhances drought tolerance by regulating stomatal closure and helps maintain water balance during drought stress (Hu et al. 2022). These metabolic adjustments collectively aid in preserving cell turgor and mitigating the physiological stress associated with water deficit. The galactose metabolism integrates with carbohydrate metabolism to help plants adapt to drought stress (Gundaraniya et al. 2020). Furthermore, when water availability declines, sucrose (from the galactose metabolism) is hydrolysed into glucose and fructose (Bagherikia et al. 2019), providing immediate energy to sustain cellular activities ensuring tuber crop survival during drought stress. The specific metabolites within galactose metabolism, such as melibiose and glycerol, enhance osmoprotection, which allows tuber crops to efficiently adapt to water stress. These adaptations highlight the critical role of galactose metabolism in improving drought resilience and offering potential for metabolic engineering aimed at enhancing drought tolerance in tuber crops.
Starch and Sucrose Metabolism
4.2.5
Drought stress significantly impacts the starch and sucrose metabolism of tuber crops, leading to the alteration of sugars in tuber crops (Geigenberger et al. 1997). During drought stress, plants tend upregulate sugars such as sucrose in tuber crops (Kaur et al. 2021) due to reduced activities of sucrose synthase, which converts sucrose into glucose and fructose. This results in decreased starch synthesis as enzymes crucial for starch biosynthesis, like ADP‐glucose pyrophosphorylase and starch synthase. Consequently, starch storage in tubers is impaired lead to poor tuber development and tuber yield (Ahmad et al. 2024). The intermediate metabolites such as glucose‐6‐phosphate and fructose‐6‐phosphate essential for starch biosynthesis decline under drought stressed conditions due to reduced photosynthetic rate (Wang, Liu, et al. 2022; Wang, Yang, et al. 2022; Thomas and Beena 2024). Furthermore, the accumulation of sucrose and its derivatives, such as alpha‐trehalose, plays a crucial role in protecting cell membranes and proteins from dehydration‐induced damage (Benaroudj and Goldberg 2001). These integrated metabolic responses optimize carbohydrate utilization, reinforcing tuber crop resilience under drought conditions. Therefore, a deeper understanding of this metabolic pathway will assist in enhancing drought tolerance tuber crop cultivars.
Summary
5
The study identified a wide range of metabolites including sugars (trehalose and sucrose), amino acids (proline and glutamate) and organic acids (malic acid and citric acid) which were altered under drought stressed conditions. These metabolites play crucial roles in osmotic regulation, oxidative stress management and energy conservation, enhancing drought tolerance mechanisms in tuber crops. The review highlighted the importance of specific metabolic pathways, including the GDM, citrate cycle and galactose metabolism, which contribute to the adaptive responses of tuber crops to water stress conditions. While the current understanding of the metabolomic responses to drought stress in these crops is significant, it remains fragmented. Further studies are necessary to address the inconsistencies in metabolite profiles across different studies and crops. In particular, the regulatory mechanisms of cassava need to be thoroughly dissected to understand its ability to withstand harsh climatic conditions including drought stress, as well as the adeptness to thrive in marginal soils. The new understanding of the crop's resilience can be adopted and help breeders in developing a wide range of climate‐smart crops for agricultural sustainability and achieving food security.
Way Forward
6
- Breeding programs should focus on enhancing the production of key metabolites such as proline, trehalose and polyamines, which play critical roles in drought tolerance.
- The key metabolic pathways involved in drought tolerance, such as the GDM, citrate cycle and galactose metabolism should be explored further through metabolic engineering to improve drought resilience in tuber crops.
- Future research should focus on standardized methodologies and experimental conditions to ensure the reproducibility of results across different studies.
- Given their protective roles in osmotic regulation, antioxidant activity and root architecture, polyamines should be explored as potential biomarkers for improving root system efficiency and water utilization under drought stress conditions.
- Integrating metabolomics with transcriptomics and proteomics could provide a more comprehensive overview of tuber crop responses to drought stress.
Funding
We would like to thank the Water Research Commission for funding this research. This paper is based on research under the project titled “An integrated multi‐omics approach to uncover drought tolerance biomarkers in two underutilised crops: sweet potato ( Ipomoea batatas L.) and cassava ( Manihot esculenta Crantz)” funded by the Water Research Commission (Project No. C2023/2024–01262), with grant funds awarded and managed by the University of South Africa (UNISA). The contents of the article are the authors' sole responsibility and should not be regarded as reflecting the views and position of the WRC and UNISA.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Table S1: List of significantly changed metabolites in selected three major tuber crops under drought stress conditions. Figure S1: Pathway analysis using all identified metabolites in major tuber crops showing metabolic pathways represented as nodes. The graph presents a view of all the matched pathways arranged by p values on the y‐axis and the pathway impact values on the x‐axis. The node color (beige to red) is based on the node's p value, and the node radius is defined by the pathway impact values. A pathway impact value > 0.1 and p < 0.05 was considered a target.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmad, D. , Y. Ying , and J. Bao . 2024. “Understanding Starch Biosynthesis in Potatoes for Metabolic Engineering to Improve Starch Quality: A Detailed Review.” Carbohydrate Polymers 346: 122592.39245484 10.1016/j.carbpol.2024.122592 · doi ↗ · pubmed ↗
- 2Ahn, S. , J. Jung , I. A. Jang , E. L. Madsen , and W. Park . 2016. “Role of Glyoxylate Shunt in Oxidative Stress Response.” Journal of Biological Chemistry 291, no. 22: 11928–11938.27036942 10.1074/jbc.M 115.708149 PMC 4882458 · doi ↗ · pubmed ↗
- 3Ali, H. , I. Mahmood , M. F. Ali , et al. 2024. “Individual and Interactive Effects of Amino Acid and Paracetamol on Growth, Physiological and Biochemical Aspects of Brassica napus L. Under Drought Conditions.” Heliyon 10, no. 11: e 31544.38882271 10.1016/j.heliyon.2024.e 31544 PMC 11176763 · doi ↗ · pubmed ↗
- 4Almeida, T. , G. Pinto , B. Correia , S. Gonçalves , M. Meijon , and M. Escandón . 2020. “In‐Depth Analysis of the Quercus suber Metabolome Under Drought Stress and Recovery Reveals Potential Key Metabolic Players.” Plant Science 299: 110606.32900444 10.1016/j.plantsci.2020.110606 · doi ↗ · pubmed ↗
- 5Arbona, V. , M. Manzi , C. de Ollas , and A. Gómez‐Cadenas . 2013. “Metabolomics as a Tool to Investigate Abiotic Stress Tolerance in Plants.” International Journal of Molecular Sciences 14, no. 3: 4885–4911.23455464 10.3390/ijms 14034885 PMC 3634444 · doi ↗ · pubmed ↗
- 6Bagherikia, S. , M. Pahlevani , A. Yamchi , K. Zaynalinezhad , and A. Mostafaie . 2019. “Transcript Profiling of Genes Encoding Fructan and Sucrose Metabolism in Wheat Under Terminal Drought Stress.” Journal of Plant Growth Regulation 38: 148–163.
- 7Barnaby, J. Y. , D. Fleisher , V. Reddy , and R. Sicher . 2015. “Combined Effects of CO 2 Enrichment, Diurnal Light Levels and Water Stress on Foliar Metabolites of Potato Plants Grown in Naturally Sunlit Controlled Environment Chambers.” Physiologia Plantarum 153, no. 2: 243–252.24888746 10.1111/ppl.12238 · doi ↗ · pubmed ↗
- 8Barnaby, J. Y. , D. H. Fleisher , S. K. Singh , R. C. Sicher , and V. R. Reddy . 2019. “Combined Effects of Drought and CO 2 Enrichment on Foliar Metabolites of Potato (Solanum tuberosum L.) Cultivars.” Journal of Plant Interactions 14, no. 1: 110–118.