Resting-state brain function and its modulation by intranasal oxytocin in antisocial personality disorder with and without psychopathy
Julia Griem, Daniel Martins, John Tully, Declan Murphy, Yannis Paloyelis, Nigel Blackwood

TL;DR
This study explores how brain function differs in people with antisocial personality disorder with and without psychopathy and how oxytocin affects these differences.
Contribution
The study reveals distinct resting-state brain function and differential effects of oxytocin in ASPD subtypes.
Findings
ASPD individuals showed reduced blood flow in frontotemporal regions compared to non-offenders.
ASPD+P showed greater blood flow increases in posterior default mode network regions than ASPD-P.
Oxytocin decreased blood flow in the left basal ganglia only in ASPD-P individuals.
Abstract
Behavioural, structural, and functional neuroimaging differences exist between individuals with antisocial personality disorder with (ASPD + P) or without psychopathy (ASPD-P). However, the aetiological mechanisms underpinning such differences remain unclear, hindering treatment development. Intranasal oxytocin (OT) has shown modulatory effects on social brain function in healthy and antisocial populations. We investigated the effects of OT on resting-state brain function in individuals with violent offending histories with ASPD+/-P using arterial spin labelling to measure regional cerebral blood flow (rCBF). A double-blind, placebo-controlled, crossover design was employed with males with ASPD (ASPD + P: N = 17, ASPD-P: N = 14) and healthy male non-offenders (N = 22). Both ASPD subtypes exhibited reduced rCBF in frontotemporal regions compared to non-offenders. Individuals with ASPD +…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
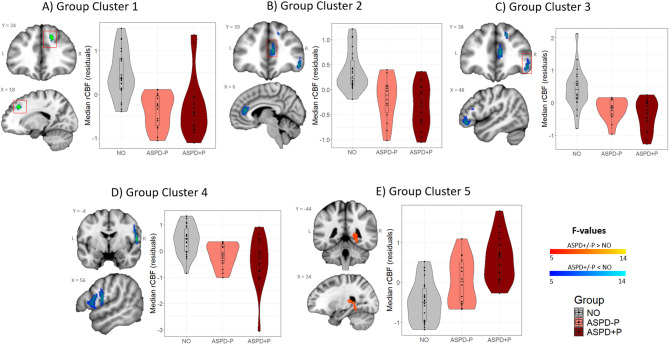 Figure 1
Figure 1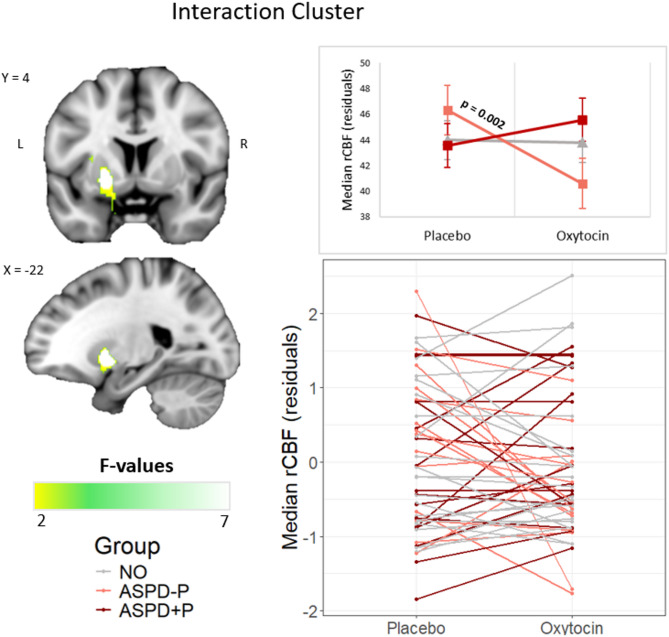 Figure 2
Figure 2- —https://doi.org/10.13039/501100000272National Institute for Health and Care Research
- —https://doi.org/10.13039/100010269Wellcome Trust
- —https://doi.org/10.13039/501100000269Economic and Social Research Council
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeuroendocrine regulation and behavior · Primate Behavior and Ecology · Human-Animal Interaction Studies
Introduction
Antisocial personality disorder (ASPD) is characterized by impulsivity, irresponsibility and aggression^1^. Approximately one third of individuals with ASPD additionally meet categorical criteria for psychopathy (ASPD+P)^2^, characterized by callous unemotional behaviours and a lack of remorse^3^. Antisocial individuals with (ASPD+P) and without (ASPD-P) psychopathy share features such as life-course persistent offending and heightened reactive aggression^4,5^. Those individuals with ASPD+P demonstrate an earlier onset and greater density of offending behaviours, increased use of proactive aggression, and a poorer response to treatment interventions than those with ASPD-P^6–10^.
Behavioural similarities between the two antisocial subgroups are underpinned by convergent neurobiological abnormalities. Meta-analytic evidence indicates that functional abnormalities in the orbitofrontal cortex, anterior cingulate, amygdala, and insula are observed in men with ASPD undertaking social and affective cognitive tasks^11^. Imbalances in the striatal excitation/inhibition ratio, which may underpin shared problems with impulsive decision-making^12^, have been observed in those with ASPD+P and ASPD-P in comparison to healthy individuals^13^. A small number of studies also suggest that the behavioural differences between the two groups are underpinned by neurobiological differences. Antisocial men with ASPD+P have reduced grey matter volumes in the anterior prefrontal cortex and the bilateral temporal poles^14^ and abnormal frontotemporal activity during empathic and reward processing tasks^15–17^ in comparison to antisocial men without psychopathy and healthy individuals.
While prior functional neuroimaging (fMRI) studies in these populations have predominantly focused on task-related brain function in ASPD, resting-state fMRI analysis offers unique insights into baseline neural activity independent of specific behavioural demands^18–21^. Resting-state function is linked to, and may even predict, general cognitive propensities that underpin behaviour such as decision-making^22–24^. Neuroimaging studies investigating resting-state brain activity in large samples of incarcerated individuals with varying degrees of psychopathy have suggested abnormalities in the form of aberrant functional connectivity or topology in frontotemporal and limbic regions including the anterior cingulate, insula, and amygdala, as well as in large-scale brain networks including the default mode network (DMN)^25–27^. Two studies of young men meeting criteria for ASPD have also implicated these regions as well as the frontoparietal attention network^28,29^. However, no study to date has directly compared resting-state brain activity between ASPD+P and ASPD-P populations.
In addition, resting-state imaging studies in these populations have typically relied on measurements of the blood-oxygen-level-dependent (BOLD) contrast, a proxy measure of neural activity based on the complex interaction between blood flow, volume and oxygen metabolism^30,31^. A closely related physiological measure, regional cerebral blood flow (rCBF) provides a more direct proxy of neural activity by indexing perfusion from cerebral capillaries into brain tissue. A small number of studies have measured rCBF using single photon emission computed tomography (SPECT) or positron emission tomography (PET) in antisocial populations, typically demonstrating decreased perfusion in anterior and medial (cingulate) frontal regions, lateral temporal regions, the amygdala and insula^32,33^, and increased perfusion in posterior DMN regions^34^. However, these studies were constrained by the recruitment of heterogenous clinical and non-clinical samples, including individuals characterised by aggression, antisocial behaviours or substance misuse disorders, but without detailed phenotypic characterization of ASPD+/-P. Furthermore, these studies used large, manually defined regions of interest and non-automated analytic approaches (e.g., visual judgment) to perfusion data. These limitations can be addressed by using arterial spin labelling (ASL), a non-invasive measure of rCBF with improved spatial resolution and excellent test-retest reliability^35,36^, to assess resting-state rCBF in individuals with carefully phenotyped ASPD+/-P.
Measurement of resting-state brain activity in clinical populations also offers a context-free measure of the brain’s responsivity to pharmacological modulation. One pharmacological agent of particular interest to the ASPD+/-P populations is oxytocin, a neuropeptide central to the regulation of complex social behaviours such as empathy^37,38^. Two recent task-based fMRI studies revealed that a single dose of intranasal oxytocin (OT) normalised aberrant neural responsivity to fearful faces in the anterior cingulate and anterior insula in the ASPD+P (but not ASPD-P) group^17^ and angry faces in the amygdala in a broadly defined ASPD group^39^. However, whether OT also modulates resting-state brain function in ASPD with or without psychopathy remains unknown. Gaining an understanding of the effect of OT on resting-state brain function in ASPD would provide valuable insight into the ‘shiftability’ (i.e., the responsivity to pharmacological modulation) of baseline brain dysfunction in the disorder^40^.
ASL imaging is more sensitive to the effects of pharmacological challenges than BOLD imaging because rCBF is less susceptible to non-specific drug effects than the BOLD signal^31,41–43^. ASL may thus be preferred for investigating OT effects on resting-state brain function. Its ability to capture the time-, method-, and dose-dependent changes in rCBF in response to oxytocin has been demonstrated^44^. In healthy individuals, OT modulates rCBF in regions including the amygdala and the anterior insula^44–46^. Importantly, these are areas where dysregulated neural activity has been demonstrated in ASPD+/-P populations^15,16^ and where task-related brain activity in individuals with ASPD+P was modulated with OT^17,39^.
We thus employed ASL imaging to examine potential differences in resting-state brain function and the impact of a single acute intranasal administration of OT on such resting-state rCBF in males with a history of violent offending with ASPD+P (N = 17) and ASPD-P (N = 22) and healthy male non-offenders (N = 22) using a double-blind, placebo-controlled, randomised crossover design. We conducted region-of-interest (ROIs: amygdala/anterior insula) and exploratory whole-brain analyses. The ROIs were selected a priori due to consistent evidence of structural and functional abnormalities in these areas in the disorder^11,47,48^ together with their demonstrated responsivity to OT in functional imaging paradigms^17,39^. For the ROI analysis, we hypothesized that: (1) both ASPD groups would show reduced amygdala and anterior insula rCBF in comparison to the non-offender group; and (2) such differences would be more marked in the ASPD+P group in comparison to the ASPD-P group. Considering the distinct normalising effects of OT on fearful face processing in ASPD+P^17^, we further hypothesized that (3) OT would restore rCBF in these ROIs in individuals with ASPD+P but not ASPD-P. For the whole-brain analyses, we further expected to find that (4) both ASPD groups would show reduced anterior frontal, anterior cingulate and lateral temporal lobe rCBF, together with increased rCBF in posterior DMN regions such as posterior cingulate and precuneus in comparison to the non-offender group and that (5) such differences would be more marked in the ASPD+P group in comparison to the ASPD-P group.
Results
Sample characteristics
Table 1 shows the demographic and clinical characteristics of the three participant groups. They did not differ significantly in age or IQ. As expected, the three groups differed significantly in years of education (the ASPD groups had fewer years of education than the non-offender group), PCL-R scores, and the aggression subscales (the ASPD groups scored higher than the non-offender group, and the ASPD+P group scored higher than the ASPD-P group). Also as expected, the ASPD+P group had a higher frequency of comorbid cluster A personality disorders than the ASPD-P group (see supplementary Table 1 for a breakdown of the comorbidities of individual personality disorders). This is in keeping with the normal range of variation in clinical profiles of ASPD+/-P^49^ and we did not adjust our analyses based on these findings. The two ASPD groups did not differ significantly in comorbid substance use disorder prevalence or positive urine drug screening tests on the days of scanning. To avoid over-correcting for phenotypic variance inherent to ASPD^50^, substance use was not included as a covariate in our main analyses. However, we note that the ASPD+P group had a significantly higher rate of cocaine use as compared with the non-offender group. Therefore, we conducted supplementary analyses which included substance use as a covariate (see supplementary Table 4 for the details about drug use across groups and supplementary results for covariate analyses). Global CBF did not differ between groups or treatment conditions (see supplementary Table 2).
Table 1Demographic and clinical characteristics of participants. Data are mean (standard deviation) unless otherwise stated. Non-offender participants did not have any diagnosis of personality disorder or mental illness and no history of conviction, therefore only the two ASPD groups were compared on these variables. Some participants did not complete the RPQ (final ASPD+P N = 13, ASPD-P N = 13, NO N = 20). RPQ = reactive proactive aggression questionnaire, PD = personality disorder, SUD = substance use disorder within past 12 months. F-statistic = ANOVA with Tukey post-hoc, H-statistic = Kruskal Wallis with Mann Whitney U post hoc, ^†^Fisher’s exact test with fisher’s exact post-hoc, χ^2^ = chi-squared test of independence with chi-squared test of independence as post hoc, U-statistic = Mann Whitney U. ◊ pairwise comparison did not survive a Sidak correction for multiple comparisons (adjusted α = 1 – (1-0.05.05)^1/3^ = 0.017).DemographicASPD+P(N = 17)ASPD-P(N = 14)NO(N = 22)Group comparisonPost hoc tests (p-values)NO vs. ASDP+PNO vs. ASPD-PASPD+P vs. ASPD-PAge (years)40.56 (9.95)45.21 (10.05)37.73 (9.56)H_(2)_ = 4.44, p = 0.11...IQ91.56 (12.45)99.14 (15.24)98.95 (10.77)H_(2)_ = 3.44, p = 0.18...Duration of education (years)9.88 (1.82)10.64 (2.21)13.77 (3.27)H_(2)_ = 19.40, p < 0.001p < 0.001p = 0.002p = 0.28Age at first violent conviction20.38 (5.45)21.43 (5.03).t = 0.55, p = 0.59...Violent convictions4.31 (2.87)3.86 (2.93).U = 112.00, p = 0.78...Reconviction within 3 years, N (%)8 (47%)6 (43%).0.06, p = 1.00 ^†^...RPQ reactive aggression15.77 (4.69)12.46 (5.77)5.90 (3.54)F_(2)_ = 19.90, p < 0.001p < 0.001p < 0.001p = 0.22RPQ proactive aggression13.85 (6.39)7.38 (6.41)0.75 (1.16)H_(2)_ = 29.30, p < 0.001p < 0.001p < 0.001p = 0.03^ns◊^RPQ total aggression29.62 (9.91)19.85 (11.46)6.65 (4.03)H_(2)_ = 27.56, p < 0.001p < 0.001p < 0.001p = 0.04^ns◊^PCL-R Factor 19.24 (3.05)4.81 (3.03)1.09 (1.63)H_(2)_ = 35.41, p < 0.001p < 0.001p < 0.001p < 0.001PCL-R Facet 1 (interpersonal)4.24 (1.82)1.81 (1.63)0.64 (0.95)H_(2)_ = 27.80, p < 0.001p < 0.001p = 0.02^ns◊^p < 0.001PCL-R Facet 2 (affective)5.00 (1.80)3.00 (1.80)0.46 (0.80)H_(2)_ = 34.47, p < 0.001p < 0.001p < 0.001p = 0.01PCL-R Factor 216.17 (1.69)11.43 (3.08)1.09 (1.44)H_(2)_ = 43.40, p < 0.001p < 0.001p < 0.001p < 0.001PCL-R Facet 3 (lifestyle)7.59 (1.23)5.36 (1.69)1.00 (1.23)H_(2)_ = 41.07, p < 0.001p < 0.001p < 0.001p < 0.001PCL-R Facet 4 (antisocial)8.47 (1.33)6.07 (2.13)0.50 (1.14)H_(2)_ = 41.84, p < 0.001p < 0.001p < 0.001p = 0.002PCL-R Total28.21 (3.22)17.51 (4.55)2.64 (2.97)H_(2)_ = 45.55, p < 0.001p < 0.001p < 0.001p < 0.001PD other than ASPD, N (%)Cluster A5 (29%)0 (0).4.91, p = 0.05^†^...Cluster B8 (47%)2 (14%).3.77, p = 0.07^†^...Cluster C1 (6%)2 (14%).0.62, p = 0.58^†^...SUD, N (%)3 (18%)4 (29%).0.52, p = 0.67^†^...ADHD, N (%)2 (12%)1 (7%).0.19, p = 1.00^†^...Positive urine drug test, N (%)Placebo scan11 (65%)3 (23%)5 (23%)8.13, p = 0.02^†^p = 0.01p = 1.00p = 0.03^ns◊^Oxytocin scan12 (71%)5 (36%)6 (27%)χ2 = 7.78, p = 0.02p = 0.01p = 0.72p = 0.08
Group, treatment, and interaction effects on rCBF
The a priori ROI analysis in the amygdala and anterior insula did not reveal significant group, treatment, or interaction effects (see supplementary Table 3). The additional Bayesian linear mixed models revealed strong evidence in favour of the null hypothesis (see supplementary materials). This indicated that there is a very low likelihood for group differences or treatment effects (or their interaction) on rCBF in the four ROIs.
The whole-brain analysis revealed a significant main effect of group in five clusters (Table 2; Fig. 1). Post-hoc pairwise comparisons showed that both ASPD+P and ASPD-P groups had reduced rCBF relative to the non-offender group in four of these clusters. These four clusters spanned the right hemisphere frontal and temporal areas (medial superior frontal gyrus, anterior cingulate cortex, orbitofrontal cortex, Rolandic operculum, pre-/post-central gyrus, and superior temporal gyrus). By contrast, in the fifth cluster, which encompassed core areas of the posterior DMN (right hemisphere posterior cingulate, precuneus, hippocampus), both ASPD groups had increased rCBF relative to the non-offender group, and individuals with ASPD+P had further increased rCBF compared to individuals with ASPD-P. There were no significant main effects of treatment. However, a significant group by treatment interaction effect was found in one cluster spanning the left basal ganglia, specifically the globus pallidus, putamen and caudate (Table 2; Fig. 2). Simple main effects tests revealed that this was driven by a significant decrease in rCBF after OT in the ASPD-P group only. These results were largely unchanged after adding substance use (presence of a positive drug screen) as a covariate (see supplementary materials).
Table 2 Clusters with significant group and interaction effects on rCBF. Cluster-wise analysisPairwise ComparisonsCluster descriptionHemisphereKP _FWE_FPeak coordinatesEstimated Marginal Means (SE)Sidak-corrected p-valuesxyzMain effect of group (F-contrast)ASPD+PASPD-PNOASPD+P vs. NOASPD-P vs. NOASPD+P vs. ASPD-PCluster 1: Medial superior frontal gyrusRight410< 0.00118.1218344039.74 (0.91)38.68 (1.03)44.70 (0.81)p < 0.001p < 0.001p = 0.83Cluster 2: Anterior cingulate cortexRight2180.00710.776361657.55 (0.75)58.48 (0.85)62.77 (0.66)p < 0.001p < 0.001p = 0.80Cluster 3: Pars orbitalis, orbitofrontal cortexRight1630.0410.324640−1258.22 (1.07)59.66 (1.22)65.53 (0.95)p < 0.001p = 0.002p = 0.77Cluster 4: Rolandic operculum, pre- and postcentral gyrus, superior temporal gyrusRight490< 0.00113.2658−41249.77 (1.66)51.75 (1.88)57.75 (1.47)p = 0.002p = 0.05p = 0.82Cluster 5: Posterior (isthmus) cingulate cortex, precuneus, hippocampusRight2700.0019.9022−42835.14 (0.71)32.31 (0.81)29.49 (0.63)p < 0.001p = 0.03p = 0.04Group x treatment interaction effect (F-contrast)PL vs. OTASPD + PASDP-PNOGlobus pallidus, putamen, caudateLeft2580.029.32−222−6PL: 43.57 (1.78)OT: 45.57 (1.78)PL: 46.33 (2.05)OT: 40.62 (2.05)PL: 43.99 (1.62)OT: 43.80 (1.62)p = 0.20p = 0.002p = 0.89Main effects were measured with F-contrasts. *PL = placebo scan, OT = *oxytocin scan. Clusters labelled according to Automated Anatomical Labelling (AAL3) atlas built into SPM12 and confirmed by mapping MNI peak coordinates to Talairach space in BioImage Suite (https://bioimagesuiteweb.github.io/bisweb-manual/tools/mni2tal.html). SE = standard error.
Fig. 1Clusters with significant group differences in rCBF. The violin plots (with box plots inside) show the marginal mean and individual datapoints (z-standardized residuals) to depict spread of the median rCBF values within each significant cluster, and to clarify post-hoc what was driving the F-effects identified in the whole-brain analysis. For D), the main effect of group and the pairwise comparisons remained significant (p = 0.008) after removing the ASPD+P outlier (z-standardized residual > |3.0|), so this was kept in the analysis to increase power. Blue shaded clusters indicate reductions, red shaded cluster indicates increase. A red box visually highlights the cluster that the plot is referring to (in case other clusters are also visible in that slice).
Fig. 2Cluster with a significant group by treatment interaction effect on rCBF. The top line plot shows the marginal means (EMMs) of the median rCBF values for each group under each treatment condition, after accounting for the effect of global median CBF, age, and minutes since dose. The bottom spaghetti plot shows individual participants’ responsivity to OT. The interaction effect and simple main effect remain significant (p = 0.01) after excluding the single ASPD-P participant with the steepest slope.
Discussion
This study provides novel evidence that individuals with a history of violent offending with ASPD+P or ASPD-P exhibit shared and distinct resting-state neurobiological features. We did not establish significant differences in resting-state rCBF between groups, or in response to OT, in either the amygdala or the anterior insula (our a priori ROIs; hypotheses 1–3). However, we did reveal the expected reductions in resting-state rCBF in frontotemporal regions in both antisocial groups compared to non-offenders, and the increased rCBF in posterior DMN regions in the ASPD+P group compared to the ASPD-P and non-offender groups (hypotheses 4 and 5). Moreover, OT selectively reduced resting-state rCBF in the left basal ganglia of the ASPD-P group but had no significant effect in the ASPD+P or non-offender groups.
The a priori ROI analysis did not reveal the expected resting-state rCBF abnormalities in the anterior insula or amygdala in the ASPD+/-P groups, supporting a recent suggestion that observed functional impairments in these brain regions in the disorder are task-dependent and context-evoked^51^, i.e., they may not have been detected in our resting-state paradigm. Equally, OT did not exert a significant modulatory effect on resting-state rCBF in these pre-specified ROIs in any group. The latter was surprising given that previous research had demonstrated resting-state rCBF changes in the anterior insula and amygdala in healthy individuals^44–46^. However, differences in the dose and timing of oxytocin administration could explain the discrepant findings. Thus, maximal resting-state effects appear to be associated with lower OT doses such as 9–18 IU^46^. OT effects in the amygdala, insula, or anterior cingulate of healthy individuals have typically been captured at an earlier time point (25–78 min post-administration^45^). However, our study was adequately powered to detect between-group rCBF differences in a crossover design^52^ and the Bayesian linear mixed models provided compelling evidence in favour of the null hypothesis in the context of this OT dose (40 IU) and time frame of post-dose assessment (85 min). Ultimately, confidence in such null findings will be best derived from mega-analytic data from collaborative neuroimaging consortia such as the ENIGMA-Antisocial Behaviour group (https://enigma.ini.usc.edu/ongoing/enigma-antisocial-behavior/). Such consortia benefit from access to within-subject multi-modal data from multiple centres, enabling improved statistical power to reveal true effects.
The exploratory whole-brain analysis did reveal the expected reductions in rCBF in both ASPD+P and ASPD-P groups compared to the non-offender group in predominantly frontal, and to a lesser extent temporal regions. These findings are consistent with (but more spatially precise than) earlier SPECT^33^ and PET^32^ findings in adult antisocial populations. These results (except for the reduced rCBF in the ASPD-P group within cluster 4) remained significant after covarying for the presence of recent substance use (see supplementary materials), suggesting that the demonstrated differences between the antisocial and non-offender groups are not simply attributable to the potential confound of comorbid substance use.
These results could have potential implications for understanding the neurobiological mechanisms underpinning the shared behavioural characteristics of individuals with ASPD+P and ASPD-P. It is important to consider that the brain’s intrinsic network architecture that is present during the resting-state shapes the brain’s functional network architecture during task performance^53^. In other words, resting-state brain function influences task-related brain function. The observed resting-state abnormalities in our study may contribute to an impaired recruitment of these areas during functional neuroimaging tasks. For example, we found reduced resting-state rCBF in both antisocial groups in the anterior cingulate cortex (cluster two) and the pars orbitalis/orbitofrontal cortex (cluster three). Functionally, these two regions play important roles in learning from punishment cues^54,55^, changing behaviour in the face of changing contingencies^56^, and decision-making under conditions of uncertainty^57,58^ – processes which appear to be similarly impaired in those with ASPD with and without psychopathy^12^. However, such potential resting-state–functional links remain speculative in this study due to the absence of additional perfusion data obtained during functional tasks. To further disentangle which specific rCBF mechanisms underpin behavioural deficits, future studies could integrate rCBF and measurements of cerebral metabolic rate of oxygen consumption (CMRO_2_)^59^ or oxygen extraction function (OEF)^60^ during behavioural tasks to determine whether these rCBF reductions reflect metabolic deficits in ASPD subtypes beyond the resting-state.
Our precise phenotyping further enabled us to demonstrate that the ASPD+P group had significantly increased rCBF in a medial parietal cluster compared to the ASPD-P and non-offender groups. Previous studies of individuals with ASPD+P have consistently found structural^14,61^ and functional^15,17,62^ activity abnormalities in this region, especially in the posterior cingulate and precuneus. Altered grey matter volume, surface area and increased BOLD demands may underpin the observed changes in rCBF. Yet there may also be important functional explanations. Typically, this region has heightened perfusion in the resting state in healthy individuals, reflecting its role as a ‘rich club’ hub in the brain’s information processing network^21,63^. However, alterations in network topology in psychopathy^27^ could make increased function in areas like the precuneus a compensatory mechanism which underpins information integration in the disorder. This is further supported by findings that the posterior DMN does not appropriately deactivate during task engagement in people with offending history and psychopathy^64^. This region also contributes to self-/other-referential processing (including mentalizing and perspective-taking), autobiographical episodic memory, and subjective reward representation^63,65–68^. Evidence suggests that automatic mentalizing and subjective reward representational processes which draw on such areas are compromised in individuals with ASPD+P^15,69–74^. Taken together, these findings suggest that the structure and function of the posterior DMN in ASPD+P is examined in more detail in future research.
Resting-state rCBF responsivity to oxytocin also differed between the ASPD groups. Thus, OT significantly reduced rCBF in the left basal ganglia, specifically the globus pallidus and dorsal striatum (putamen and caudate), when compared to placebo, in the ASPD-P but not the ASPD+P group. Importantly, this result is unlikely to reflect non-specific vascular effects, as OT (10–40 IU) does not disrupt cerebrovascular reactivity^44^.
The basal ganglia, rich in oxytocin receptors^75^, are crucial for reinforcement learning, reward processing, habit formation, and goal-directed action selection and control^76–78^. The dorsal striatum predominantly mediates choice impulsivity, evaluating action-contingent outcomes to better select future goal-directed actions^79^. Functional MRI studies have provided evidence for dorsal striatum abnormalities in antisocial groups in childhood^80^. Youths with disruptive behaviour disorders (precursors of ASPD in adulthood) show reduced responsiveness to positive prediction errors and increased responsiveness to negative prediction errors within the dorsal striatum during feedback^81^ and reduced dorsal striatal response to early stimulus-reinforcement exposure^82^. Therefore, striatal dysfunction may underpin dysfunctional learning and decision-making in antisocial populations. Given the modulatory effect of OT on dorsal striatum function in this and other research^45,83^, and evidence for a beneficial effect of OT on reinforcement learning^84–86^, further studies investigating the potential therapeutic relevance of OT for reinforcement learning and decision-making in adults with ASPD are warranted.
We observed no significant OT effects on resting-state function in the ASPD+P group. This may be due to neurochemical differences between individuals with ASPD+P and ASPD-P. One possibility is that individuals with ASPD+P have higher peripheral endogenous OT levels^87,88^, potentially limiting the impact of exogenous administration^89^ (though see^90^. Alternatively, spatial pharmacodynamic studies show differential effects of OT on rCBF at different post-dose intervals^44,45^, and it is possible that effects in ASPD+P^17^ may have been more immediate or transient than in ASPD-P and thus not captured within our time window of 85 min post-dose. From a neurochemical perspective, such differential effects of timing between the groups might be associated with differences in oxytocin receptor genotype or availability^91,92^, or interactions with other neurotransmitters such as serotonin^93,94^. However, these conclusions are tentative and further research exploring the neurochemical underpinnings of ASPD+/-P is required.
We also did not observe significant modulatory effects of OT on resting-state brain function in the healthy non-offender group, despite previous studies demonstrating significant modulatory effects. This could be related to different time points of measurement as detailed above. Further studies utilising differing post-dose sampling time frames and different OT doses are required.
Several limitations warrant consideration. First, our study was limited to male participants, precluding conclusions about potential sex differences in OT responsivity^95^. Second, while we excluded individuals with history of traumatic brain injury (TBI) or loss of consciousness for more than 1 h, we did not exclude individuals with mild TBI. Mild TBI has been suggested to have a transient impact on perfusion^96^. Third, OT effects were measured 85 min post-dose, which is later than most fMRI studies^17,39,44,45^ including those which have demonstrated a functional impact of OT in antisocial populations. Fourth, our use of cluster-level inference and spatial smoothing meant we were only able to interpret the significance of whole clusters, and not individual voxels. While this approach is more powerful, it is also linked with a slightly lower level of spatial specificity^97^. Therefore, the findings of increased rCBF in both ASPD+/-P in the superior temporal lobe should be interpreted cautiously, and future studies may wish to use threshold-free cluster enhancement methods^98^. Finally, while we observed significant OT effects in the ASPD-P group, there was nevertheless individual variability in neural response to OT within and across the groups (Fig. 2), suggesting the need for personalized treatment approaches informed by individual level data. Larger trials incorporating genetic and endocrine markers of OT signalling may help to further enhance precision in ASPD treatment strategies.
In conclusion, we employed ASL to measure rCBF in carefully phenotyped groups of violent men with ASPD with or without psychopathy. Such men showed shared (reduced rCBF in frontotemporal regions) and distinct (increased medial parietal rCBF in the ASPD+P group in comparison to ASPD-P and non-offender groups) resting-state abnormalities. OT exerted a modulatory effect on rCBF in the left basal ganglia of the ASPD-P group. The stratification of ASPD into more biologically homogenous clinical subgroups for both mechanistic and therapeutic studies appears warranted.
Methods
Participants
This study included 53 male participants (31 offenders with ASPD with (N = 17) or without (N = 14) psychopathy and 22 healthy non-offenders) aged 18–60, with normal range IQ according to the Wechsler Abbreviated Scale of Intelligence (WASI-II)^99^ who consented between September 2017 and March 2020. We recruited males with convictions for violent crimes (murder, rape, attempted murder, grievous and actual bodily harm) who met DSM-5 criteria for ASPD via the National Probation Service of England and Wales and local forensic personality disorder services. We recruited healthy individuals without previous convictions from the general population through public and online advertising. All participants completed diagnostic interviews (Structured Clinical Interview for the DSM-5-Clinical and Personality Disorders (SCID-5-CV/SCID-5-PD)^1,100^ and Psychopathy Checklist-Revised (PCL-R)^3^) and authorized access to their criminal records. In line with previous research in UK samples^13–15,17,101^, we used a score of 25 as the threshold for psychopathy in this English population. We excluded participants if they had a documented or self-disclosed history – or met criteria during the diagnostic assessment – of major mental illness (bipolar 1, bipolar 2, major depression or psychotic disorders including schizophrenia, schizoaffective disorder and delusional disorder), neurological disorders, head injury resulting in loss of consciousness for 1 h or longer, severe visual or hearing impairments, or contraindications to MRI.
The study was approved by London City and East Research Ethics Committee (15/LO/1083), as well as the National Offender Management Services Research Committee (2016 − 382). After receiving a complete description of the study, all participants completed signed informed consent and were assigned anonymized subject identifiers. All assessments were conducted by an experienced research psychologist (JG) and forensic psychiatrist (JT). All research was performed in accordance with relevant guidelines and regulations and in line with the peer-reviewed and ethically approved study protocol. This trial was registered at ClinicalTrials.gov (ID NCT05383300, https://clinicaltrials.gov/study/NCT05383300) on 20/05/2022. The CONSORT checklist and flowchart can be found in the supplementary materials. Participants completed the self-report Reactive-Proactive Aggression Questionnaire. On the day of each MRI scan, participants were asked to confirm that they did not misuse substances that day and provided a urine sample to assess for substance use up to 30 days prior.
Study design and procedure
We used a double-blind, placebo-controlled, randomised crossover design. We acquired anatomical and ASL scans as part of a larger imaging protocol conducted at the Centre for Neuroimaging Sciences, IOPPN, King’s College London (see supplementary materials for the full schedule, noting the gap between task-based scans and ASL imaging to minimise carry-over effects). For the two scanning sessions, we randomly and blindly allocated the participant to receive 40 international units (IU) of OT (Syntocinon, Novartis, Switzerland) or placebo (PL; same excipient without the oxytocin). The dose is in line with other research and has been deemed safe with no side effects^44–46,102^. Scans were scheduled at least 3 days apart to ensure complete drug washout. We counterbalanced the order of administration for OT/PL across participants, with half receiving OT first and then PL, and the other half receiving the reverse order. According to the recommendation for standardised administration^103^ and under supervision of the researcher, participants received training and then self-administered the nasal spray by snorting one puff every 30 s through alternating nostrils, for 5 min (10 puffs with 4 IU each). The ASL scans (6:23 min) were acquired on average at 84 (± 9) minutes and 85 (± 12) minutes after administration of PL and OT, respectively. This time delay is referred to as the variable ‘minutes since dose’. Scans were acquired at the same time of the day within and between participants (see supplementary materials for details).
Image acquisition
We used a General Electric MR750 3Tesla MRI scanner and 32-channel C-RMNova head coil for this study. During each session, we acquired a 3-dimensional pseudo-continuous ASL scan (60 slice partitions with thickness and gap = 3 mm, TE = 1109 ms, TR = 5180 ms, flip angle = 111°, FOV = 240 × 240 mm^2^, in-plane resolution = 3.6 mm; radiofrequency inversion pulse = 1825 ms, delay = 2025 ms, control-label image pairs = 5). The acquisition of the final rCBF map for each participant at each session was in line with recommendations^104^.
We also acquired a 3D high-resolution T1-weighted whole-brain anatomical image during each scanning session (MPRAGE, 196 slices with thickness and gap = 1.2 mm, TE = 3.02 ms, TR = 7.31 ms, TI = 400 ms, FA = 11°, FOV = 270 × 270 mm^2^, matrix = 256 × 256, voxel resolution = 1.05 × 1.05 × 1.2 mm^3^).
Image pre-processing
We confirmed good image quality and presence of typical perfusion values ranging between 20 and 110 ml/100 g/minute across the whole brain, indicating accurate computation of CBF maps^104^, using FSLeyes. We pre-processed the scans in the Automatic Software for ASL Processing (ASAP) toolbox, version 4.0 in Matlab 2018b. Steps included (1) co-registration of proton density and T1 images; (2) subject-specific normalization of CBF maps; (3) skull-stripping, segmentation and removal of extra-cerebral signal from the normalized CBF maps; (4) partial volume correction and normalization of the CBF map to MNI152 space; and (5) 8 mm Gaussian spatial smoothing of the CBF map. An explicit grey matter tissue probability mask (20%), derived by thresholding the FSL grey matter template at 0.20, was applied. We calculated global median CBF for each participant and each session and included this in statistical analysis as a covariate of no-interest to improve signal-to-noise ratio and the sensitivity of within-subject changes in local areas. We used median rCBF values throughout due to a skew towards the lower end of the typical range.
Statistical analysis
We compared demographic and clinical characteristics, as well as global median CBF using ANOVA, ANCOVA, and Chi-squared tests, or non-parametric equivalents when normality assumptions were not met, in SPSSv29. We interpreted significant main effects via Sidak-corrected pairwise comparisons.
For the a priori ROI analysis, we extracted median rCBF from the bilateral amygdalae and anterior insulae using Harvard-Oxford atlas masks in MNI space. We assessed main effects of group, treatment, and their interaction with bootstrapped linear mixed models in JASP, covarying for global CBF, age, and minutes since dose. We applied FDR correction for multiple comparisons across the ROIs. To further evaluate the robustness of our findings, we conducted Bayesian linear mixed models post hoc (see supplementary methods).
We conducted whole-brain analyses in SPM12 (www.fil.ion.ucl.ac.uk/spm) using a partitioned errors approach to accommodate repeated-measures design assumptions^105^. We examined the main effect of group using a one-way ANOVA on individual-averaged OT and PL CBF maps. We assessed the main effect of treatment via a one-sample t-test using difference images, subtracting an individual’s PL CBF map from their OT CBF map. We tested the group by treatment interaction effect with a one-way ANOVA on the difference images. We covaried for global CBF, age, and minutes since dose in all models. We applied F-contrasts to assess main and interaction effects, with cluster-level inference using a cluster-forming threshold of p = 0.005 and family-wise error correction at α = 0.05; in accordance with prior ASL studies investigating OT effects^44,45,106^. For significant clusters, we extracted and compared raw median CBF values using post-hoc pairwise comparisons or simple main effects tests with the Sidak correction for multiple comparisons in SPSSv29.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1First, M. B., Williams, J. B. W., Benjamin, L. S. & Spitzer, R. L. User’s Guide for the SCID-5-PD (Structured Clinical Interview for DSM-5 Personality Disorder) (American Psychiatric Association, 2015).
- 2Hare, R. D. Manual for the Hare Psychopathy Checklist-Revised (Guilford, 1991). 10.1007/978-0-387-79948-3_837
- 3Molloy, M. F. et al. Regional, but not brain-wide, graph theoretic measures are robustly and reproducibly linked to general cognitive ability. Cereb. Cortex 35 (2025).10.1093/cercor/bhaf 074PMC 1301751440211548 · doi ↗ · pubmed ↗
- 4Wechsler, D. Wechlser Abbreviated Scale of Intelligence, Second Edition (WASI-II) (NCS Pearson, 2011). 10.1177/0734282912467756
- 5First, M. B., Williams, J. B. W., Karg, R. S. & Spitzer, R. L. User’s Guide To Structured Clinical Interview for DSM-5 Disorders, Clinical Version (American Psychiatric Association, 2016).