DEC1 regulates human β cell functional maturation and circadian rhythm
Sam Preza, Bliss Zheng, Zihan Gao, Akshaya Biju, Mai Liu, Zhihui Cheng, Matthew Choi, Juan R. Alvarez-Dominguez

TL;DR
DEC1, a circadian regulator, is essential for the maturation of human β cells in islet organoids, improving their glucose response and function.
Contribution
DEC1 is identified as a key circadian factor that links circadian rhythms to β cell maturation in human islet organoids.
Findings
DEC1 is specifically active in maturing β cells and regulates glucose-responsive insulin secretion.
DEC1 deficiency impairs insulin release and glucose metabolism in islet organoids.
DEC1 synchronizes circadian rhythms and enhances islet organoid functionality.
Abstract
Stem cell-derived islet (SC-islet) organoids offer hope for cell replacement therapy in diabetes, but their immature function remains a challenge. Mature islet function requires the β cell circadian clock, yet how the clock regulates maturation is unclear. Here, we show that a circadian transcription factor specific to maturing SC-β cells, DEC1, regulates insulin responsiveness to glucose. SC-islet organoids form normally from DEC1-ablated human pluripotent stem cells, but their insulin release capacity and glucose threshold fail to increase during in vitro culture and upon transplant. This deficit reflects the downregulation of maturity-linked effectors of glucose utilization and insulin exocytosis, blunting glycolytic and oxidative metabolism, and is rescued by increasing metabolic flux. Moreover, DEC1 is needed to boost SC-islet maturity by synchronizing circadian glucose-responsive…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4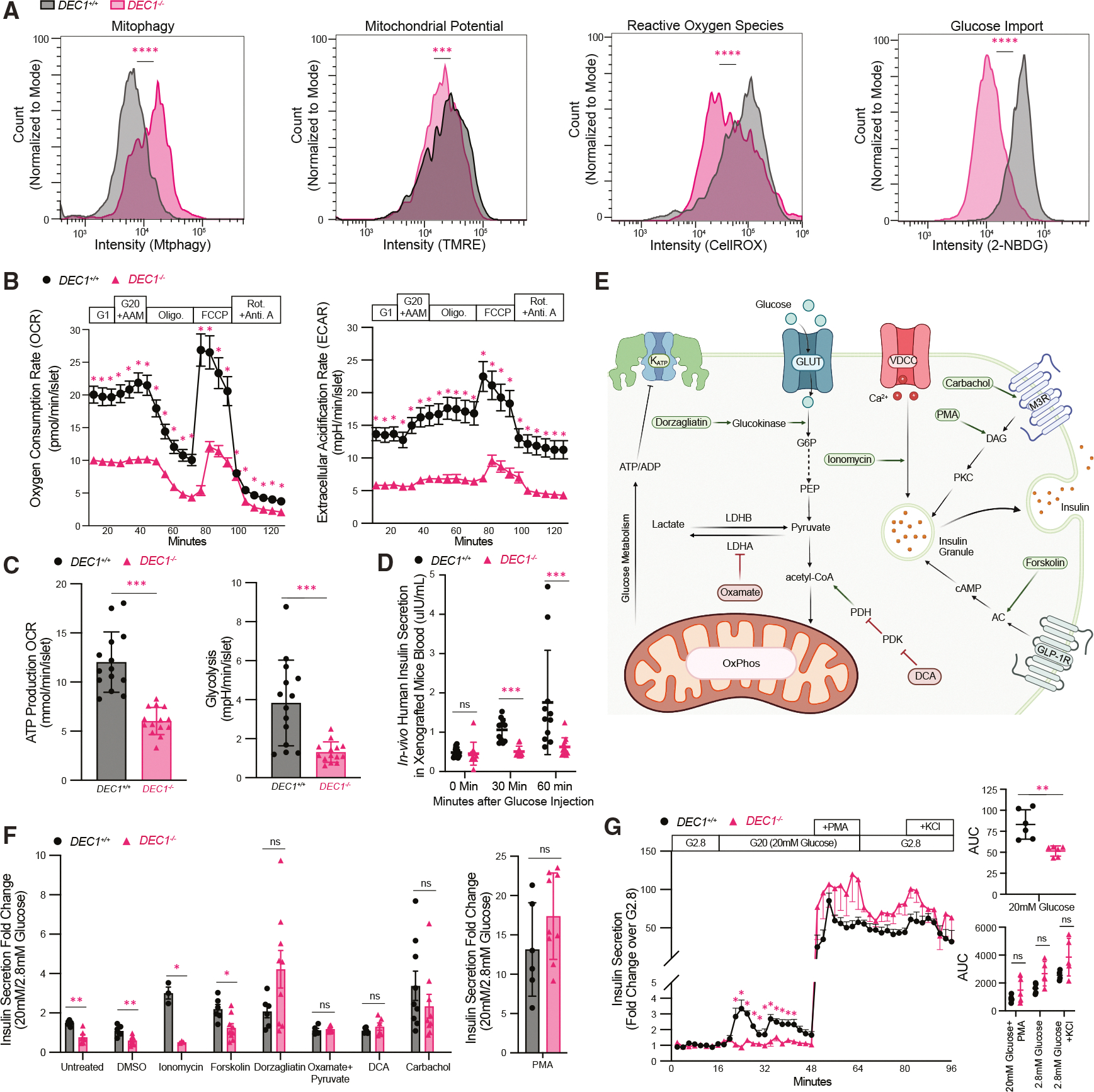 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPancreatic function and diabetes · Diabetes and associated disorders · Adipose Tissue and Metabolism
INTRODUCTION
The promise of broad cell therapy for diabetes has grown closer to reality with the advent of human pluripotent stem cell (hPSC)-derived islet organoids (SC-islets)^1–3^ Islet transplants can cure insulin-dependent diabetics^4^ but are limited by a scarcity of acceptable islets. SC-islets offer limitless cells for transplantation therapy, which can render human recipients free of daily insulin injections.^5,6^ Yet, in vitro SC-islets are molecularly and functionally less mature than transplanted ones,^7–9^ and both lack the kinetics, precision, and magnitude of glucose-stimulated insulin secretion (GSIS) of adult islets.^10,11^ This delays the therapeutic benefit from SC-islet transplants, posing a key challenge to their broad application.
Mature islet function develops after birth, as hormone secretion capacity and the glucose threshold for secretion increase.^12–14^ These changes are intertwined with the postnatal onset of islet circadian clocks.^15^ The clock in islet β cells aligns peak GSIS responsiveness with the active phase of the day,^16^ likely to adapt to postnatal feeding-fasting rhythms.^8^ Inactivating CLOCK or BMAL1—the master clock activators—renders mouse islets unable to mount mature GSIS responses, causing diabetes.^17–20^ Further, we find that inducing clock genes, via daily metabolic stimulation and recovery cycles, fosters mature function in human cadaveric/SC-islets by prompting circadian GSIS responses with a raised glucose threshold.^9^ How the clock coordinates metabolic maturation of human β cells, however, remains poorly understood.^10,21^
To elucidate the regulation of human SC-β cell maturation by the circadian network, we focused on the transcription factor DEC1 (also called BHLHE40 or SHARP2). DEC1 entrains circadian rhythms to environmental cues, including light and feeding,^22,23^ by competing with CLOCK:BMAL1 for DNA E-box binding.^24^ Recently, we found that mouse DEC1 coordinates islet GSIS by synchronizing energy metabolism and exocytic gene rhythms and that Dec1 knockout renders mouse islets immature, causing lifelong glucose intolerance due to insufficient insulin responses.^25^ We previously predicted that DEC1 partakes in the core regulatory circuit that defines human β cell identity.^9^ However, DEC1’s role in human β cell development, functional maturation, and metabolism remains unexplored.
Here, we report that DEC1 is essential for SC-β cell maturation in vitro and in vivo. SC-islet organoids differentiate normally from DEC1-ablated hPSCs in scalable suspension culture^26^ but exhibit impaired GSIS in both static and dynamic assays. RNA sequencing (RNA-seq) of purified SC-β cells links this defect to the downregulation of maturity-associated genes for glycolysis, mitochondrial respiration, and insulin exocytosis. Accordingly, DEC1^−/−^ SC-islet organoids exhibit an energy deficit, prompted by lower glucose uptake, that hinders glycolysis and oxidative respiration and incites mitophagy. This renders DEC1^−/−^ organoids unable to mature even after transplantation into the kidney capsule of immunodeficient mice. We further find that DEC1 is required for enhancing SC-islet maturity through synchronization of circadian clock machinery and GSIS rhythms. Importantly, the GSIS deficit of DEC1^−/−^ organoids can be rescued pharmacologically, by insulinotropic agents that bypass metabolism or increase metabolic flux. These findings establish DEC1 as a key link between the circadian clock and human β cell metabolic maturation, highlighting the potential of leveraging circadian control to generate SC-islets with adult-like GSIS for improved transplantation therapy.
RESULTS
DEC1 is required for mature insulin responses to glucose in human SC-islets
To evaluate DEC1’s role in human islet development, we tracked its expression at each stage of our scalable protocol for differentiating 3D SC-islet organoids in suspension bioreactors (Figure 1A).^26^ DEC1 is minimally detectable in differentiating progenitors but enriched in SC-islet β cells, present in ~97% of β cells and induced 12-fold (Figures 1B and S1F). We thus disrupted DEC1 in independent hPSC lines using an inducible CRISPR-Cas9 system (Figure S1A).^27^ DEC1^−/−^ lines differentiated normally, forming 3D SC-islet organoids with intact morphology, distribution of α (glucagon+/C-peptide+), β (C-peptide+/glucagon− ≡ C-peptide+/NKX6.1+), and enterochromaffin-like (SLC18A1+/C-peptide−) cells, and insulin production (Figures S1C–S1G). We verified DEC1 mRNA and protein loss throughout differentiation and in vitro maturation by extended culture (28 days) in serum-free media without added factors (Figures 1C, S1B, and S1F).^28,29^ DEC1 absence impaired the induction of IAPP, a marker of SC-β cell functional maturation^30^ (Figures 1D and S1H). Strikingly, DEC1^−/−^ SC-islets show markedly diminished GSIS responses in both serial static and dynamic assays (Figures 1E–1G). Insulin secretion was significantly higher under static non-stimulatory (2.8 mM) glucose incubations, while subsequent responses to saturating (20 mM) glucose were impaired, despite normal insulin release following chemical depolarization with KCl (Figure 1E). Thus, DEC1^−/−^ SC-islet organoids show elevated insulin secretion under non-stimulatory glucose and lower GSIS capacity—hallmarks of immature β cell function.^12–14^
Longitudinal assays over 4 weeks of extended in vitro culture further revealed that DEC1^−/−^ SC-islets fail to expand their GSIS capacity, as evidenced by a consistently lower insulin secretion fold change from 2.8 to 20 mM glucose (Figure 1F). In dynamic assays, although DEC1^−/−^ SC-islets retain a biphasic pattern of insulin secretion, they show significantly reduced peak stimulation (4.14 ± 1.4-fold decrease) under saturating glucose, leading to diminished cumulative secretion (Figure 1G).
DEC1 both regulates and is regulated by the circadian clock.^22,31^ Given DEC1’s requirement for mature GSIS responses in SC-islets, we asked if the clock contributes to SC-islet maturity. To test this, we used small interfering RNAs (siRNAs) to deplete BMAL1 and PER2 in day 14 SC-islets, core transcriptional regulators of the clock’s activating and repressing arms, respectively (Figure S1I). PER1- and BMAL1-knockdown SC-islets showed diminished GSIS responses in serial static and dynamic assays, reminiscent of neonatal/functionally immature islets.
Collectively, our findings show that DEC1 is critical for refining GSIS responses during SC-islet maturation in vitro, including suppressing basal insulin secretion and expanding GSIS capacity.
DEC1 regulates β cell maturity-linked effectors of glucose metabolism and insulin secretion
To study DEC1’s molecular role in SC-islet maturation, we performed longitudinal RNA-seq of DEC1^+/+^ and DEC1^−/−^ organoids at in vitro maturation days 7, 14, 21, and 28 (Figures S2A and S2B). 2,556 genes were differentially expressed, including the suppression of IAPP and effectors of GSIS and metabolic signaling (PCSK1, GLP1R, and SYT13^32^) and induction of lipid metabolism regulators (ACSL1 and ELOVL2). To investigate DEC1’s role specifically in SC-β cells, we performed RNA-seq in CD49a-purified cells from both day 1 and day 14 SC-islets (Figures 2A, 2B, and S2C).^29^ We identified a total of 2,622 genes significantly affected by DEC1 loss (Figures 2C–2F, S2D, and S2E). These comprise downregulated genes impacting β cell function through roles as regulators or effectors of glucose import (POU4F2),^33^ sensing (HK2),^34^ glycolysis (LDHB),^35^ mitochondrial oxidative phosphorylation (NDUFA4),^36^ redox homeostasis (TXNRD2),^37,38^ and insulin secretion (ADCY5).^39^ Genes upregulated in DEC1^−/−^ SC-β cells, by contrast, include liver genes (ALB and AFP); disallowed glycolytic enzymes (LDHA and PDK3); mitochondrial depolarization (TSPO and RACK1)^40,41^ and autophagy (PINK1, MMP9, and BNIP3)^42–45^ machinery; and fatty acid metabolism enzymes (ACSL1, ACSL6, and ELOVL2).^46–48^ Interestingly, core circadian clock genes (BMAL1, CLOCK, and PER2) lack differential expression (Figure 2B). Lastly, we confirmed the RNA-seq results by qPCR for a subset of genes (LDHA, LDHB, SYT13, and GCK) in enriched SC-β cells (Figure 2E).
Our qPCR validation showed that in addition to IAPP, the key maturity markers MAFA and SIX2 are also depleted in DEC1^−/−^ SC-β cells (Figure 2E). To assess this comprehensively, we examined a set of 2,218 genes differentially expressed across human β cell maturation pseudotime^49^ and found that they are globally depleted, with IAPP and PAX4 among the leading-edge subset of genes driving the downregulated β cell maturity signature (Figures 2C–2F).
Together, these findings implicate DEC1 in the control of GSIS maturation through the regulation of glucose import and metabolism, mitochondrial homeostasis, and insulin release.
DEC1 controls circadian insulin secretion and clock gene rhythms in human SC-islets
The circadian clock attunes insulin secretion to daily feeding-fasting cycles^16^ and is needed for mature β cell function.^16,50^ Previously, we reported that SC-islet organoids also display circadian GSIS responsiveness rhythms.^9^ Given DEC1’s ability to entrain circadian rhythms to environmental cues,^22,23^ we investigated its requirement for GSIS rhythms. After synchronizing DEC1^+/+^ and DEC1^−/−^ SC-islets with 1-h forskolin^20,51^ or dexamethasone^52^ pulses, we performed sequential static GSIS and collected RNA over two 24-h cycles (Figure 3A). DEC1^+/+^ SC-islets showed an autonomous 24-h GSIS rhythm (p = 7.6E–07 from Jonckheere-Terpstra-Kendall [JTK] algorithm and 9.70E–10 by Rhythmicity Analysis Incorporating Nonparametric methods [RAIN]), first peaking at 36 h post-synchronization (Figure 3B). At these peak times, GSIS capacity was, on average, 2-fold greater than in mock-treated SC-islets, which did not exhibit a GSIS rhythm (Figures 3B and 3C). This greater GSIS responsiveness was driven by lower insulin secretion under non-stimulatory (2.8 mM) glucose (Figure S3A), as reported.^9^ Strikingly, DEC1^−/−^ SC-islets failed to show circadian GSIS responsiveness or suppressed basal insulin secretion upon synchronization, with no improvement in GSIS capacity (Figures 3B, 3C, and S3A). We verified circadian DEC1 expression in synchronized but not mock-treated DEC1^+/+^ SC-islets (Figure 3D). We also validated antiphasic expression of the core clock activator BMAL1 and the PER2 and NR1D1 repressors in synchronized DEC1^+/+^ SC-islets but not in mock-treated or DEC1^−/−^ SC-islets (Figures 3D and S3B). Since DEC1 regulates β cell maturity and mitochondrial fission genes, we also examined expression patterns for MAFA, a key driver of mature β cells,^53,54^ and DRP1, which mediates mitochondrial fission.^55^ Remarkably, DEC1 loss abrogates circadian MAFA and DRP1 expression (Figure 3E).
Together, these data demonstrate that DEC1 is essential for proper circadian rhythms in GSIS and for the rhythmic expression of core clock, mitochondrial dynamics, and β cell maturity effectors.
DEC1 sustains bioenergetics and mitochondrial integrity in maturing SC-β cells
Given DEC1’s role in regulating genes essential for respiration and redox homeostasis, we probed its influence on SC-β cell energetics and mitochondrial integrity. We find that DEC1^−/−^ SC-β cells show diminished mitochondrial membrane potential, along with elevated mitophagy (Figure 4A), consistent with mitochondrial dysfunction.^45,56,57^ Analyzing oxygen consumption rates in day 14 DEC1^−/−^ SC-islet organoids reveals severe reductions in basal, glucose-stimulated, ATP-linked, maximal, and non-mitochondrial respiration (Figures 4B, 4C, and S4A). Cellular reactive oxygen species levels are also significantly lower in DEC1^−/−^ SC-β cells (Figure 4A), in line with diminished respiration. Thus, DEC1 is vital to bolster the mitochondrial oxidative activity driving glucose-coupled energetics.
Reduced mitochondrial activity upon DEC1 loss could reflect adaptations to reduced glucose utilization. Extracellular acidification rate analyses in day 14 SC-islet organoids reveal that glycolysis and glycolytic capacity are indeed hampered by DEC1 loss (Figures 4B, 4C, and S4A). Further, we trace such glucose utilization deficits to stifled glucose import, as shown by the severely impaired uptake of a fluorescent glucose analog in DEC1^−/−^ SC-β cells (Figures 4A and S4D). Taken together with dulled glycolysis and mitochondrial oxidative activity,^58^ these data argue that DEC1 promotes glucose import and utilization, stimulating the energy flux that underpins insulin secretion.
A metabolic flux deficit underlies impaired insulin secretion upon DEC1 loss
To test the extent to which the maturation defect of DEC1^−/−^ organoids can be rescued by the in vivo milieu, we transplanted day 21 SC-islets under the kidney capsule of immunocompromised mice. Strikingly, DEC1^−/−^ SC-islets show blunted GSIS responses relative to DEC1^+/+^ SC-islets even 19 weeks post-transplant (Figure 4D), as indicated by significantly reduced human insulin secretion following an intraperitoneal glucose injection. We thus conducted a targeted screen of insulinotropic agents to rescue the GSIS defect of DEC1^−/−^ SC-islets in vitro (Figures 4E–4G). Stimulation with forskolin, an adenylyl cyclase agonist, failed to restore GSIS, suggesting defects in steps upstream of cAMP-mediated secretory amplification. Ionomycin, a Ca^2+^ ionophore, also failed to rescue GSIS, consistent with normal Ca^2+^ influx dynamics in the absence of DEC1 (Figure S4B). Conversely, insulin secretion is restored to DEC1^+/+^ levels by glucose-independent stimulation with either the diacylglycerol mimetic phorbol 12-myristate 13-acetate (PMA) or with carbachol, a muscarinic receptor agonist (Figures 4E–4G). Thus, we sought to rescue the glucose coupling defect of insulin secretion in DEC1^−/−^ organoids by increasing metabolic flux. Raising the pyruvate pool via methyl-pyruvate supplementation combined with inhibition of lactate dehydrogenase A using oxamic acid or increasing the acetyl-coenzyme A (CoA) pool via inhibition of pyruvate dehydrogenase kinase with dichloroacetate (DCA) successfully restores GSIS to wild-type levels (Figure 4F). Similarly, enhancing glycolytic flux via the glucokinase activator dorzagliatin drastically improved GSIS to DEC1^+/+^ levels (Figure 4F). Thus, enhancing metabolic flux rescues the GSIS defect of DEC1^−/−^ SC-islets, demonstrating that DEC1 promotes glucose import and utilization to stimulate the coupling of insulin secretion to glucose metabolism in mature β cells.
DISCUSSION
Transplantable SC-islet organoids are likely to become a therapeutic option for severe insulin-dependent diabetes. Yet, human SC-islet transplants today exhibit delayed therapeutic benefit, with 4 months needed for >85% of recipients to reach <7% hemoglobin A1C (HbA1c) levels,^6^ in contrast to 75 days or less for cadaveric islet transplants.^59^ This delay likely reflects a need for SC-islets to complete functional maturation in vivo, in line with gradual molecular changes observed after transplant.^9,60^ Common among these changes is the induction of circadian clock controllers,^7,49^ and we previously showed that recreating circadian rhythms in vitro enhances SC-islet maturity.^9^ How the circadian clock coordinates SC-islet maturation, however, has remained elusive.
Here, we show that a circadian transcriptional regulator, DEC1 (SHARP2 or BHLHE40), is vital for metabolic maturation of SC-β cells in vitro and in vivo. We previously predicted DEC1 to be the most interconnected transcription factor in the core regulatory circuit defining mature human β cell identity.^9^ Consistently, DEC1 is specifically induced in maturing SC-β cells, and its absence thwarts the induction of maturity-linked gene circuits. The disrupted circuits control glucose import and metabolism, mitochondrial dynamics, and insulin exocytosis. Accordingly, DEC1^−/−^ SC-islets show impaired glucose uptake, glycolysis, and oxidative metabolism. These changes stunt the ability of DEC1^−/−^ organoids to mount circadian GSIS responses, which fail to mature even after in vivo transplant but are rescued by pharmacologically increasing metabolic flux. Thus, our findings demonstrate that DEC1 orchestrates the coupling of insulin secretion to glucose sensing, linking the clock to the acquisition of the mature β cell phenotype.
Our report of a circadian islet maturation factor offers a distinct paradigm for how the clock programs metabolic specialization. Mature β cell physiology is thought to be orchestrated by the CLOCK/BMAL1 complex, as inactivating Clock or Bmal1 renders mouse islets immature.^17–20^ However, roles for CLOCK/BMAL1 on human islet maturation are not well defined. Three lines of evidence argue that such regulation is indirect. First, CLOCK and BMAL1 are already present early in human β cell development, before maturation.^61^ Second, they are not part of the core regulatory circuit of mature human β cells.^9^ Third, we find here that DEC1 loss suffices to render SC-islets immature, despite intact CLOCK and BMAL1. Our data thus support DEC1 as the direct arm of the clock imparting circadian GSIS control to drive β cell maturation.
Mechanistically, our results indicate that DEC1 mediates maturation in SC-β cells by coordinating glucose utilization downstream of core clock genes. DEC1-lacking β cells exhibit reduced glucose influx and compensatory responses. These responses include induction of lactate dehydrogenase A, which diverts pyruvate away from mitochondrial metabolism toward lactate production, and of lipid utilization genes, likely signaling a shift to alternative energy substrates. These adaptations recall the metabolic flexibility of neonatal β cells, which lack strict glucose dependence for insulin release and show higher lipid utilization.^10,14,62^ Notably, stimulating glucose utilization via glucokinase activation rescues GSIS to wild-type SC-β cell levels, ruling out glycolytic defects. Dorzagliatin stimulated insulin secretion more than inhibiting lactate dehydrogenase A or pyruvate dehydrogenase kinase—interventions expected to increase pyruvate and acetyl-CoA entry into the tricarboxylic acid (TCA) cycle, respectively. These data provide further evidence for an intrinsic SC-islet bottleneck in glucose metabolism to pyruvate, occurring downstream of glucokinase but upstream of the TCA cycle, possibly through diversion into serine/glycine shunt.^49,63^
DEC1’s role in glucose metabolism is consistent with its ability to synchronize circadian rhythms with feeding, possibly through ChREBP-mediated nutrient-sensing mechanisms, although these mechanisms are not well defined in islets.^22,23^ Since islet circadian rhythms emerge postnatally,^15^ along with the onset of daily feeding-fasting, they likely foster maturation, at least in part, through DEC1-mediated optimization of glucose metabolism to daily energetics. We further found, in separate mouse studies,^25^ that DEC1 orchestrates mature GSIS responses by directly binding genes regulating the integration of energy metabolism and insulin exocytosis to synchronize their expression and thus energy and secretory rhythms. Taken together, our human and mouse studies reveal an evolutionarily conserved mechanism whereby DEC1 links circadian clockwork to β cell energetics and metabolic specialization.
How can DEC1’s pathway be harnessed to improve SC-islet organoid maturity? Unlike global manipulation of metabolic cycles or the entire circadian network, the precise tuning of DEC1 levels offers a path to co-opt bioenergetic rhythms to control SC-β cell maturity. This may become possible by identifying DEC1’s nutrient-sensing mechanisms that could be targeted for external modulation. The persistent functional defect of transplanted DEC1^−/−^ SC-islets suggests that such modulation would be relevant in vivo. Thus, further delineating how the circadian system underpins SC-β cell maturity heralds opportunities to attain fully functional SC-islets to improve islet replacement therapies.
Limitations of the study
The experimental systems that were used in this study have limitations to consider. We used SC-islet organoids differentiated from the HUES 8 hPSC line, so genotype-specific effects were not evaluated. Our in vitro SC-islet differentiation protocol inherently produces some batch-to-batch differences in organoid functionality. Therefore, we assayed at least three independent SC-islet differentiations in each experiment. To assess in vivo function, we transplanted SC-islet organoids under the kidney capsule of immune-deficient mice, so DEC1’s impact on SC-islet function under diabetic conditions was not investigated. Mechanistic links between DEC1 and target gene programs are inferred from perturbation studies; direct gene occupancy by DEC1 was not determined. Finally, our findings derive from an SC-islet organoid system that lacks the vascular, immune, and endocrine-neural inputs of native islets, so potential modulatory effects of blood perfusion, immune surveillance, and tissue crosstalk on DEC1-dependent maturation were not assessed.
RESOURCE AVAILABILITY
Lead contact
Inquiries related to additional information, resources, or reagents should be directed to the lead contact, Juan R. Alvarez-Dominguez ([email protected] ).
Materials availability
All cell lines and oligonucleotides generated in this study are available from the lead contact with a completed materials transfer agreement.
Data and code availability
Raw and processed RNA-seq data generated in this study have been deposited at the Gene Expression Omnibus (GEO) and are publicly available through the accession number GEO: GSE302939, as reported in the key resources table.No original code was generated in this work.Any additional information required to reanalyze the data reported in this study is available from the lead contact upon request.
STAR★METHODS
EXPERIMENTAL MODEL AND SUBJECT DETAILS
Human cell lines
The HUES8 (NIH hESC registry #09–0021; male) line was used for directed differentiation, and lines derived from HUES8 were used for CRISPR-Cas9 genome editing followed by directed differentiation. Undifferentiated cells were maintained as aggregates in supplemented mTeSR1 medium (StemCell Technologies) using spinner flasks (Corning) set at a 70rpm rotation rate in a 37°C 5% CO_2_ incubator.
METHOD DETAILS
SC-islet differentiation in 3D suspension culture
Directed differentiation of human pluripotent stem cells into pancreatic islets was performed as previously described in 30-mL stirred suspension bioreactors (REPROCELL; ABBWVS03).^26^ All experiments were conducted using the HUES 8 (NIH registry #NIHhESC-09–0021) line maintained in mTeSR1 (StemCell Technologies, 85850) media on a rotator stir plate (Chemglass) at 70 RPM in a humidified 37°C, 5% CO_2_ tissue culture incubator. Maintenance cultures were passaged every 3 days using a sequential filtration approach: 300 μm strainers were used to remove large aggregates, followed by 37 μm strainers (StemCell Technologies, 27250) to eliminate small debris. Cells were then incubated with Gentle Cell Dissociation Reagent (GCDR, StemCell Technologies, 100–0485) for 6 min, counted for viability, and reseeded at 0.6 × 10^6^ cells/mL in mTeSR1 supplemented with 10 μM ROCK inhibitor (Y27632, Abcam). Cells underwent at least three passages after thaw from cryopreservation before differentiation, with each passage achieving a minimum 3-fold expansion. A six-stage differentiation protocol was used to generate SC-islets. Cells were seeded at 0.6 × 10^6^ cells/mL in mTeSR1 with Y27632 on day 0, typically using 18M cells in 30 mL total volume. A half-media change was performed at 24 h, replacing with fresh mTeSR1 (without Y27632), followed by a full media change on day 2. Differentiation was initiated on day 3 (Stage 1) and proceeded through Stage 6, with media changes performed on a 24-h feeding cycle as described.^26^
Reverse transcription-quantitative PCR (RT-qPCR)
RNA extraction and purification was done using the DirectZol RNA miniPrep (Zymo Research, R2050), which included a DNase treatment to eliminate genomic DNA contamination. Complementary DNA (cDNA) synthesis was carried out using the High-Capacity cDNA reverse transcription kit (Invitrogen, 18080051), according to manufacturer instructions. qPCR reactions were performed using SYBR green master mix (Applied biosystems, A25742) on a StepOnePlus Real-Time PCR system (Applied biosystems). All reactions were done in triplicate, and melting curves were run to establish specificity of amplification. The primer sequences used for amplification are listed in the key resources table. All qPCR CT values are normalized to 18S rRNA (ΔCT).
Static glucose-stimulated insulin secretion
Static glucose-stimulated insulin secretion (GSIS) assays were performed on Stage 6 clusters.^26^ Clusters were first washed and equilibrated in KREBs buffer (128 mM NaCl, 5 mM KCl, 2.7 mM CaCl2, 1.2 mM MgSO4, 1 mM Na2HPO4, 1.2 mM KH2PO4, 5 mM NaHCO3, 10 mM HEPES, 0.1% BSA) containing 2.8 mM glucose for 1 h. Then, we used 12 μm polycarbonate Millicell inserts (Sigma-Aldrich, PIXP01250) in 24-well plates to sequentially incubate clusters in low glucose (2.8 mM, 1 h), high glucose (20 mM, 1 h), and KCl (30 mM in 2.8 mM glucose, 1 h). Media samples were collected after each incubation period. Following the assay, clusters were dispersed into single cells using TrypLE (Fisher Scientific, 50–591-419), which were counted to normalize secretion data. Insulin concentrations in the collected samples were measured using an ultrasensitive human insulin ELISA (ALPCO, 80-INSHUU-E01.1), with results expressed as μIU/mL/1000 cells. For insulin content measurements, cluster lysates were prepared using M-PER (Thermo Fisher, 78501) and analyzed using the same ELISA protocol. All assays were performed in triplicate.
Dynamic glucose-stimulated insulin secretion
Dynamic glucose-stimulated insulin secretion assays were conducted using an automated perifusion system (BioRep). Size-matched islets were carefully selected and placed in perifusion chambers. The chambers were first equilibrated with KREBs buffer containing basal glucose (2.8 mM) for 60 min at a constant flow rate of 100 μL/min. Following equilibration, islets were sequentially exposed to low glucose (2.8 mM) for 16 min, then high glucose (20 mM) for 32 min. For dynamic GSIS with insulinotropic agent, (10μM) PMA an additional 20 mM glucose step incorporating PMA was done for 16 min, followed by 16 min 2.8mM glucose, and 16 min of 2.8 mM glucose + 30 mM KCl. Insulin secretion data were normalized to the mean insulin release during the initial basal incubation.
Flow cytometry for differentiation cell markers
Suspension culture Samples were treated with TrypLE to dissociate clusters to single cells, which were then fixed in 4% Paraformaldehyde for 30 min at 4°C. The fixed cells were then blocked at 4°C for 30 min in a blocking buffer (5% Donkey serum [Jackson Immunoresearch], 0.1% Saponin [Sigma Aldrich] in PBS) to permeabilize cell membranes and minimize nonspecific binding. Primary antibody incubations were conducted overnight at 4°C, with antibodies selected based on the differentiation stage of the cells. The following day samples were incubated at room temperature for 1 h with the appropriate fluorophore-conjugated secondary antibodies. The cells were resuspended in a washing buffer (5% donkey serum, PBS) and analyzed using an Accuri C6 Plus flow cytometer (BD Biosciences). Data acquisition and analysis were performed using FlowJo software. The antibodies used are listed in key resources table and are as previously reported.^26^
Immunofluorescence staining
Measurements were performed based on previously established protocols.^9,26^ SC-islet organoids were collected and fixed in 4% paraformaldehyde for 15 min at room temperature and washed with PBS. After fixation organoids were resuspended in embedding gel (2.5% agar, 2.5% gelatin). A droplet of the solution was transferred into an embedding cassette, stored in 70% ethanol, and then submitted to the University of Pennsylvania Pathology and Imaging Core (RRID: SCR_022420) for paraffin embedding. For deparaffinization the slides were treated with xylene, followed by serial ethanol and water washes. Antigen retrieval was done by incubating the slides in warm 1X Citrate buffer (Sigma Aldrich, C9999–1000ML) using a crockpot water bath at “high” temperature for 5 min and then “warm” for 30 min. The slides were subsequently blocked and permeabilized using a blocking buffer consisting of 5% donkey serum, 0.3% Triton and PBS solution for 1 h to minimize nonspecific binding. Primary antibody staining was conducted overnight in 4°C for C-PEPTIDE, DEC1, Glucagon and SLC18A1 (key resources table). After incubation with primary antibodies the slides were incubated with the appropriate secondary antibodies for an hour at room temperature and then counterstained with DAPI (Thermo Fisher, P36935). After staining, fluorescent imaging was performed using a fluorescent microscope.
SC-β cell enrichment with CD49a
Fluorescence-activated cell sorting (FACS) was performed as previously reported^29^ to isolate live, marker-defined β-cell populations from Stage 6 cultures. Cells were first dissociated into a single-cell suspension using TrypLE enzymatic digestion. Aggregates were dispersed mechanically through repeated pipetting with a P1000 micropipette, ensuring uniform suspension while minimizing cellular stress. The digestion reaction was quenched with S3 medium, followed by centrifugation at 300 × g for 5 min. The pellet was resuspended in sorting buffer (PBS + 1% BSA + 2 mM EDTA), filtered through a 35 μm strainer, and counted to determine cell concentration. For immunostaining, cells were aliquoted at 10 × 10^6^ cells/mL and incubated with anti-human CD49a-PE (1:11 dilution) (Miltenyi Biotec, 130–101-397) antibody for 20 min at room temperature, protected from light with gentle agitation every 3–4 min. After incubation, cells were washed twice by centrifugation (300 × g, 5 min) and resuspended in sorting buffer at a final concentration of 10 × 10^6^ cells/mL. FACS sorting was performed using a 100 μm nozzle at a rate of ~10,000 events per second. CD49a^+^ cells were selectively sorted to enrich for insulin-producing β-cells within the Stage 6 population. Cells were collected into FACS tubes containing 1 mL sorting buffer supplemented with 1:1000 ROCK inhibitor (Y27632) to enhance post-sort survival. Sorted β-cells were kept on ice and subsequently transferred to pre-warmed media for further functional characterization or RNA extraction for downstream RNA-sequencing.
RNA-sequencing and differential gene expression
RNA-sequencing was performed as previously described.^9^ RNA from CD49a enriched SC-β cells (~300k cells) or SC-islets (~300k cells) was isolated for RNA-sequencing. Sequenced reads were STAR^65^ aligned using default parameters. Read counts per gene were quantified using HTSeq^66^ and normalized using DESeq.^67^ The expression levels were normalized to counts per million (CPM), with genes considered expressed if CPM was greater than 1.
Gene set enrichment analysis
Gene lists ranked by log_2_fold change (DEC1^−/−^/DEC1^+/+^) were analyzed for enrichment of genes using GSEA^68^ with default parameters and “-metric log2_Ratio_of_Classes.” Pathway annotations were derived from Hallmark and C5 (Gene Ontology) databases to investigate the overrepresentation of curated pathways and biological processes.
Forskolin and dexamethasone synchronization experiments
Forskolin and dexamethasone was used to synchronize circadian rhythms^20,51^ prior to downstream assays. Stage 6 SC-islet organoids were incubated in Stage 3 media supplemented with 10μM forskolin (Stemgent, 04–0025) or 10μM dexamethasone (Sigma, D4902–500mg) for 1 h. Following the 1-h forskolin or dexamethasone shock, SC-islets were returned to culture and allowed to recover in fresh Stage 3 media for an additional 15 h. After the 15-h post-shock incubation period, SC-islets were harvested for downstream analyses including static GSIS and RT-qPCR. Insulin secretion fold changes and relative expression of genes of interest were analyzed for circadian rhythmicity using both the JTK_CYCLE and RAIN algorithms implemented in Nitecap61.^64^ JTK_CYCLE employs nonparametric rank-based testing to detect periodic signals by comparing time-series data against idealized symmetric waveforms and assesses rhythmicity significance by fitting the data across multiple candidate periods. This process yields empirical p values and estimates of the peak phase within a 24-h cycle.
In parallel, RAIN (Rhythmicity Analysis Incorporating Nonparametric Methods) was applied as a complementary nonparametric approach. It tests rhythmicity by evaluating whether the data exhibit a significant rising trend followed by a falling trend within each cycle, allowing the peak to occur at any phase. The method uses exact null distributions derived from randomization principles to compute p values, offering enhanced robustness to asymmetric waveforms and outliers. Both methods were used to test a 24-h period.
The MetaCycle meta2d() function was used with default parameters to calculate rhythmicity metrics, including Benjamini-Hochberg corrected p values and phase estimates. For visualization, time-series data was normalized to max expression.
Knockdown of PER1 and BMAL1 using siRNA
SC-islets at day-14 were cultured in 6-well ultra-low attachment plates at a density of 1×10^6^ cells per well in complete medium. For each well, 9 μL of Lipofectamine RNAiMAX was diluted in 150 μL of Opti-MEM I Reduced-Serum Medium (Thermo Fisher Scientific). Separately, 3 μL of siRNA (20 μM stock; see key resources table for product number) was diluted in 150 μL of Opti-MEM Medium. The diluted siRNA was combined with the diluted Lipofectamine RNAiMAX reagent at a 1:1 ratio (v/v), mixed gently, and incubated for 5 min at room temperature to allow complex formation. The resulting siRNA-lipid complexes (300 μL total volume) were added drop-wise to each well.
Following transfection, plates were placed on an orbital shaker rotating at 300 rpm in incubator for 72 h. The knockdown efficiency was validated via quantitative reverse transcription polymerase chain reaction (qRT-PCR). Functional assessment of insulin secretion was subsequently performed using static and dynamic GSIS assay.
Cellular and mitochondrial energetics analysis
Flow cytometry analyses of mitochondrial autophagy, mitochondrial membrane potential, cellular reactive oxygen species, and glucose analogue uptake were conducted in day 14 SC-islet organoids. Clusters were dissociated into single-cell suspensions using TrypLE and resuspended in Stage 3 media. Single-cell suspensions were incubated with a freshly prepared staining solution containing 50nM TMRE (Abcam; ab113852), 5μM CellROX Deep Red (Invitrogen, C10422), 20μM 2-NBDG (Thermo Fisher, N13195), or 100nM Mtphagy dye (Dojindo, Mt048) in Stage 3 medium. CD49a antibody (1:50) was included during incubation to gate-select for SC-β cells. Cells were incubated at 37°C for 30 min in a humidified 5% CO_2_ incubator. After staining, cells were washed twice with 1% BSA in PBS and filtered through a 35 μm nylon mesh into flow cytometry tubes (STELLAR SCIENTIFIC, FSC-9005). Flow cytometry data was acquired using a BD Accuri C6 Plus flow cytometer (BD Biosciences) and analyzed using FlowJo v10.8.1 (BD Life Sciences). Fluorescence intensities were normalized to unstained controls and data were gated based on forward and side cell scatter parameters to exclude debris and doublets.
Metabolic flux analysis
Oxygen consumption rate (OCR) measurements were performed using the Seahorse XFe96 Analyzer and Spheroid FluxPak (Agilent, 102905–100) to assess mitochondrial function in SC-islets at day 14 of in vitro maturation. One day prior to the assay, sensor cartridge wells were hydrated by incubating with 200 μL of Seahorse calibrant solution (Agilent) at 37°C overnight in a non-CO_2_ incubator. Spheroid plate wells were coated with 30 μL of 100 ng/mL poly-D-lysine (Sigma, P7280). Injection basal media (Agilent; 103335–100), wash media (basal media with 0.5% FBS), and assay media (basal media with 0.5% FBS and 1mM glucose) were freshly prepared and stored at 4°C protected from light. On the day of the experiment, all media were adjusted to pH 7.4 and warmed and kept at 37°C. SC-islets were hand-selected and washed twice in wash for 15 min at room temperature. After washing, 8–10 size-matched SC-islets were transferred per well in a total volume of 30 μL using an islet-loading mold and gently overlaid with 175 μL of pre-warmed assay media. The Spheroid plate was incubated for 1 h at 37°C in a CO_2_-free incubator to allow for metabolic equilibration. Prior to the assay, the sensor cartridge was calibrated on the Seahorse XFe96 Analyzer for 15 min. Mitochondrial stress testing was then performed by sequential injection of 20 μL the following compounds, prepared in injection basal media and loaded into the cartridge injection ports: 20 mM glucose with 0.5mM glutamine and 3.5mM amino acid mix, 5 μM oligomycin (Sigma, O4876), 2 μM FCCP (Sigma, C2920), and a combination of 5 μM rotenone (Sigma, 557368) and 5 μM antimycin A (Sigma, A8674). Assay cycles consisted of 2 min of mixing followed by 3 min of measurement. The number of cycles performed was as follows: 6 for basal OCR, 4 for 20 mM glucose with 0.5mM glutamine and 3.5mM amino acid mix, 5 for oligomycin, 4 for FCCP, and 6 for rotenone + antimycin A.
Transplantation studies
Transplantation of SC-islets under the kidney capsule of 8–12 week-old immunodeficient SCID-beige mice was done as previously described.^9^ SC-islets were washed and resuspended in RPMI and submitted to University of Pennsylvania Stem Cell and Xenograft Core (SCXC – RRID: SCR_010035). The procedure was performed under anesthesia. Post-surgery, mice were monitored for pain and distress, with symptomatic mice isolated and observed for 2 days. If conditions did not improve, euthanasia was performed.
Insulinotropic agents screen
SC-islets were prepared and incubated in 2.8mM glucose KREBs buffer in typical static GSIS fashion. Insulinotropic agents were added to KREBs buffer during the high glucose (20 mM glucose) 1 h incubation step at 37°C in a humidified 5% CO_2_ incubator. The following compounds were used: dorzagliatin (10μM) (Selleckchem, S6921), forskolin (10 μM) (Stemolecule, 04–0025), PMA (10μM) (Santa Cruz Biotechnology, SC-3576B), ionomycin (10μM) (StemCell Technologies, 73722), and carbachol (1mM) (Santa Cruz Biotechnology, SC-202092). For select metabolic flux modulators, longer pre-incubation was required: cells were treated with either 1 mM dichloroacetate (DCA) (Selleckchem, S8615) or 1 mM oxamate (MedChemExpress, HY-W013032A) for 24 h prior to GSIS. For oxamate-treated samples, 100μM methyl pyruvate (Sigma Aldrich, 371173) was also added during the 1-h high-glucose stimulation step only. Following treatment, GSIS was performed as described above. Media samples were collected for insulin quantification, and data were normalized to cell counts.
Calcium influx
SC-islets were washed twice, followed by staining with 16.7μM Fura2-AM (Life Technologies; F-F1225) in Krebs buffer supplemented with 5mM glucose for 45 min at 37°C. Islets were then transferred to a perifusion chamber on a homeothermic platform, perifused with 37°C Krebs buffer at a flow rate of 1mL/min and imaged with an inverted Zeiss microscope (Axio Observer.Z1). After imaging in the absence of substrate, a physiological amino acids mixture (0.5 mM glutamine and 3.5mM amino acid mix) alone and with increasing glucose levels (3mM, 16.7mM) were sequentially applied, followed by washing all substrates away and applying KCl (30mM). Intracellular Ca^2+^ was measured by dual-wavelength fluorescence microscopy using the Zeiss AxioVision system.
QUANTIFICATION AND STATISTICAL ANALYSIS
No statistical methods were used to predetermine sample size or remove outliers. To assess the statistical difference between two sets of unpaired data, GraphPad Prism was used to calculate statistical significance for all non-RNA sequencing data. To assess the statistical difference between two sets of unpaired, the Shapiro-Wilk normality test was first performed as column or grouped analysis. A two-sided unpaired t test was then used to assess confidence on the measured difference of their mean values. For unpaired data that did not follow a normal distribution, we used a non-parametric Wilcoxon rank-sum test to determine if they belong to the same parent distribution.
Supplementary Material
1
2
3
4
SUPPLEMENTAL INFORMATION
Supplemental information can be found online at https://doi.org/10.1016/j.celrep.2025.116666.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Melton D (2021). The promise of stem cell-derived islet replacement therapy. Diabetologia 64, 1030–1036. 10.1007/s 00125-020-05367-2.33454830 PMC 8012315 · doi ↗ · pubmed ↗
- 2Sneddon JB, Tang Q, Stock P, Bluestone JA, Roy S, Desai T, and Hebrok M (2018). Stem Cell Therapies for Treating Diabetes: Progress and Remaining Challenges. Cell Stem Cell 22, 810–823. 10.1016/j.stem.2018.05.016.29859172 PMC 6007036 · doi ↗ · pubmed ↗
- 3Hogrebe NJ, Ishahak M, and Millman JR (2023). Developments in stem cell-derived islet replacement therapy for treating type 1 diabetes. Cell Stem Cell 30, 530–548. 10.1016/j.stem.2023.04.002.37146579 PMC 10167558 · doi ↗ · pubmed ↗
- 4Verhoeff K, Marfil-Garza BA, and Shapiro AMJ (2021). Update on islet cell transplantation. Curr. Opin. Organ Transplant. 26, 397–404. 10.1097/MOT.0000000000000891.34148980 · doi ↗ · pubmed ↗
- 5Wang S, Du Y, Zhang B, Meng G, Liu Z, Liew SY, Liang R, Zhang Z, Cai X, Wu S, (2024). Transplantation of chemically induced pluripotent stem-cell-derived islets under abdominal anterior rectus sheath in a type 1 diabetes patient. Cell 187, 6152–6164.e 18. 10.1016/j.cell.2024.09.004.39326417 · doi ↗ · pubmed ↗
- 6Reichman TW, Markmann JF, Odorico J, Witkowski P, Fung JJ, Wijkstrom M, Kandeel F, de Koning EJP, Peters AL, Mathieu C, (2025). Stem Cell-Derived, Fully Differentiated Islets for Type 1 Diabetes. N. Engl. J. Med. 393, 858–868. 10.1056/NEJ Moa 2506549.40544428 · doi ↗ · pubmed ↗
- 7Augsornworawat P, Hogrebe NJ, Ishahak M, Schmidt MD, Marquez E, Maestas MM, Veronese-Paniagua DA, Gale SE, Miller JR, Velazco-Cruz L, and Millman JR (2023). Single-nucleus multiomics of human stem cell-derived islets identifies deficiencies in lineage specification. Nat. Cell Biol. 25, 904–916. 10.1038/s 41556-023-01150-8.37188763 PMC 10264244 · doi ↗ · pubmed ↗
- 8Helman A, Cangelosi AL, Davis JC, Pham Q, Rothman A, Faust AL, Straubhaar JR, Sabatini DM, and Melton DA (2020). A Nutrient-Sensing Transition at Birth Triggers Glucose-Responsive Insulin Secretion. Cell Metab. 31, 1004–1016.e 5. 10.1016/j.cmet.2020.04.004.32375022 PMC 7480404 · doi ↗ · pubmed ↗