Draft genome sequence of Actinopolyspora saharensis DSM 46666, a rare actinomycete isolated from the Algerian Sahara
Shuangqing Zhou, Rafika Saker, Noureddine Bouras, Guendouz Dif, Yvonne Mast, Imen Nouioui

TL;DR
This paper presents the draft genome sequence of a rare soil bacterium from the Sahara desert, revealing its potential to produce useful secondary metabolites.
Contribution
The paper provides a new draft genome sequence for Actinopolyspora saharensis DSM 46666 and identifies its biosynthetic gene clusters.
Findings
The genome is 4.67 Mbp in size with a G+C content of 69.5%.
It contains 14 biosynthetic gene clusters for secondary metabolite production.
Carbohydrate metabolism is supported by carbohydrate-active enzyme gene repertoires.
Abstract
Actinopolyspora saharensis DSM 46666 (=H244), a rare actinomycete, was isolated from Palm grove in the Oasis of Inghid in the Mzab region of the Algerian Sahara and deposited in the German Collection of Microorganisms and Cell Cultures (DSMZ). Here, we report the draft genome sequence of strain DSM 46666, with a size of 4.67 Mbp and a G+C content of 69.5 %. Bioinformatic analysis of the genomic sequence revealed the carbohydrate-active enzyme gene repertoires involved in carbohydrate metabolism and the occurrence of 14 biosynthetic gene clusters disclosing the secondary metabolite capacity of strain DSM 46666.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
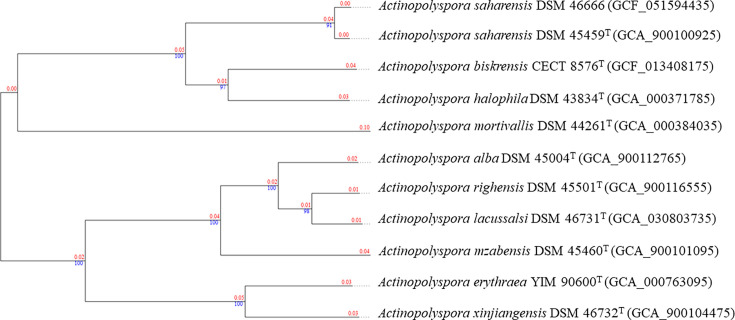 Fig. 1
Fig. 1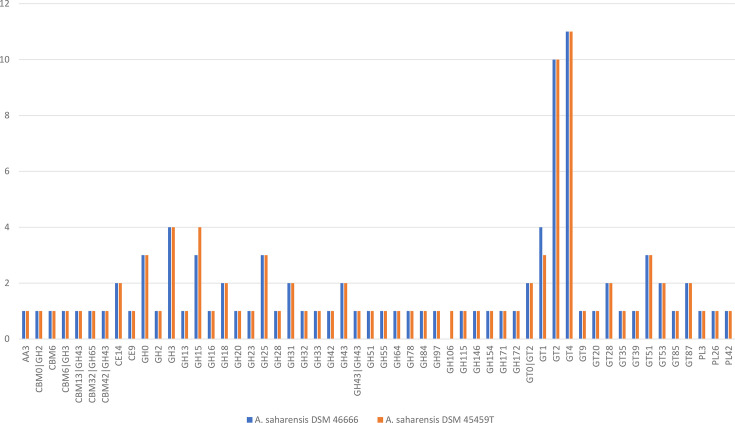 Fig. 2
Fig. 2| Strain DSM 46666 | Strain DSM 45459 | |
|---|---|---|
| Genome size (Mb) | 4.7 | 4.7 |
| G+C content (mol%) | 69.5 | 69.5 |
| CDSs | 4,058 | 4,052 |
| RNA numbers | 73 | 72 |
| N50 | 247.4 kb | 2.6 Mb |
| L50 | 7 | 1 |
| Contigs | 70 | 2 |
| Coverage | 53× | 356× |
| BioSample | SAMN49880177 | SAMN04489718 |
| BioProject | PRJNA1289121 | PRJEB15893 |
| Completeness (%) | 97.98 | 96.91 |
| Contamination (%) | 1.88 | 1.65 |
| Accession number |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Microbial Natural Products and Biosynthesis · Enzyme Production and Characterization
Data Summary
This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession number JBPQHB000000000. The version described in this paper is version JBPQHB000000000. The raw sequence reads have been submitted to Sequence Read Archive at NCBI (SRX31383543). The 16S rRNA gene sequence of Actinopolyspora saharensis DSM 46666 has been deposited at GenBank under the accession number KJ574187.
Announcement
The genus Actinopolyspora of the family Pseudonocardiaceae comprises 12 validly named species, with Actinopolyspora halophila as the type species (https://lpsn.dsmz.de/genus/actinopolyspora) [1]. Members of this genus are considered rare actinomycetes and have been isolated from saline, hypersaline and alkaline environments and are known to produce bioactive compounds with antimicrobial, antitumour and biofortification properties [24]. However, little is known about their ecological diversity and functional capabilities. Comparative genomic analysis reveals that the enzymatic activity associated with dienelactone hydrolase is highly conserved within the species of Actinopolyspora saharensis, despite the different geographical and ecological origins of the strains [5]. In view of investigating the German Culture Collection (DSMZ) for a rare halophilic actinobacterial strain, the genome of A. saharensis DSM 46666 was sequenced and its biosynthetic potential was analysed based on genome mining appraoch.
A. saharensis DSM 46666 (=H244) was isolated from a desert soil sample collected from Palm grove in the Oasis of Inghid, Béni-Isguen, Ghardaia, Mzab region, Algeria, as described by Meklat et al. [6] and deposited at the DSMZ culture collection. The strain was cultivated in DSMZ 1504 medium supplemented with 15% of NaCl and incubated at 28 °C for 14 days. Under this growth condition, it had white aerial mycelium and beige substrate mycelium, as shown in the DSMZ online catalogue (https://www.dsmz.de/collection/catalogue/details/culture/DSM-46666). DNA extraction and whole-genome sequencing of the strain were carried out by MicrobesNG service [Birmingham, UK, (https://microbesng.com/)]. In total, 50 mg of wet biomass was suspended in 500 µl of DNA/RNA Shield buffer (Zymo Research, California, USA) and sent to the sequencing service. The bacterial suspension was lysed with 120 µl of Tris/EDTA buffer supplemented with proteinase K (VWR Chemicals, Ohio, USA) and SDS (Sigma-Aldrich, Missouri, USA) at final concentrations of 0.1 mg ml^−1^ and 0.5% (v/v), respectively. The lysate was incubated for 5 min at 65 °C. An equal volume of solid-phase reversible immobilization beads was used for the purification of genomic DNA, which was resuspended in EB buffer (10 mM Tris/HCl, pH 8.0). DNA quantification was measured using the Quant-iT dsDNA HS assay (Thermo Fisher Scientific) on an Eppendorf AF2200 plate reader (Eppendorf UK Ltd., UK). Genomic libraries were prepared with the Nextera XT Library Prep Kit (Illumina, San Diego, California, USA) according to the manufacturer’s instructions with the following modifications: the DNA was increased twofold and the PCR elongation time was increased to 45 s. A Hamilton Microlab STAR automated liquid handling system (Hamilton Bonaduz AG, Switzerland) was used for library preparation. Genome sequencing was carried out on an Illumina NovaSeq 6000 platform (Illumina, San Diego, USA) with 2×250 bp paired-end reads, achieving ~53× coverage. Read trimming was carried out with Trimmomatic (v0.3) [7] with a window quality cutoff of Q15. De novo assembly was carried out using SPAdes v3.7 [8]. Prokka v1.11 was used for contig annotation [9]. The authenticity of the strain was confirmed by comparing the 16S rRNA gene sequence obtained by PCR with the corresponding sequence extracted from the assembled genome. The genome sequence was submitted to GenBank under accession number JBPQHB000000000 and automatically annotated via NCBI Prokaryotic Genome Annotation Pipeline (PGAP) [1012].
The final assembly of strain DSM 46666 revealed a genome size of 4.7 Mbp, with a G+C content of 69.5 mol%, 4,058 CDSs, 73 RNAs, an N50 of 247.4 kb and an L50 of 7 (Table 1). Taxonomic assignment placed the strain within the genus Actinopolyspora, based on 16S rRNA gene sequence, which showed 99.8% identity to A. saharensis DSM 45459^T^, as determined using the EzBioCloud server [13]. The phylogenetic relatedness of DSM 46666 to validly named Actinopolyspora species was further assessed through taxogenomic analyses using the type strain genome server (TYGS) [14]. Strain DSM 46666 formed a well-supported subclade with A. saharensis DSM 45459^T^, adjacent to a subclade containing Actinopolyspora biskrensis and A. halophila (Fig. 1). Digital DNA–DNA hybridization (dDDH) and average nucleotide identity (ANI) values between strain DSM 46666 and its close phylogenomic neighbour, A. saharensis DSM 45459^T^, were determined using the TYGS and the EzBioCloud servers [15], respectively. The average aligned length for ANI between the studied genomes was 3,194,858 pb. The dDDH (95.5%) and ANI (99.1%) values were above the established species delineation thresholds of 70% (dDDH) and 95–96% (ANI), confirming that strain DSM 46666 belongs to the A. saharensis species [1619].
Genome Blast Distance Phylogeny (GBDP) tree inferred with FastME 2.1.6.1 from GBDP distances calculated from genome sequences, illustrating the evolutionary relationship between A. saharensis DSM 46666 and Actinopolyspora species validly named. The branch lengths are scaled according to the GBDP distance formula d5. The blue numbers are GBDP pseudo-bootstrap support values >60% from 100 replications, with an average branch support of 98.0%. The red numbers are the branch length values.
The abundance and distribution of carbohydrate-active enzyme (CAZyme) families and their domains in the genome sequence of strain DSM 46666 and DSM 45459^T^ were assessed across the analysed genomes using the dbCAN3 platform (https://bcb.unl.edu/dbCAN2/) [2021]. The outcome of this analysis provides a better understanding of the role of these strains in ecosystems and can be used to further study research aimed at exploiting the biotechnological potential of these strains through enzyme engineering. To ensure reliability, a CAZyme family was considered present only if it was independently confirmed by all three integrated tools, HMMER [2223], DIAMOND [2425] and Hotpep [26], which work in a complementary manner. The annotation with dbCAN was performed using three algorithms with very conservative thresholds: DIAMOND (e-value<1×10⁻¹⁰²), HMMER (e-value<1×10⁻¹⁷ and coverage>0.45) and Hotpep (frequency>2.6 and number of hits>6). HMMER utilizes hidden Markov models to identify conserved sequence profiles, DIAMOND performs fast similarity searches against reference CAZyme databases and Hotpep detects conserved short peptide motifs characteristic of CAZymes. To enhance the reliability of the annotations, only genes confirmed by all three tools were retained, following a consensus-based scoring approach. The CAZyme repertoires of the two A. saharensis strains are highly similar, with a total of 94 CAZyme genes in strain DSM 46666 and 95 in strain DSM 45459^T^ (Fig. 2). This represents ~2% of the total CDSs in each genome, highlighting a consistent investment in carbohydrate metabolism across both strains. The accession numbers for the predicted CAZyme protein sequences of strain DSM 46666 were listed in Table S1 (available in the online Supplementary Material). These findings expand our knowledge of the biological roles and potential applications of CAZymes of this taxon.
Comparison of predicted CAZymes of two strains of A. saharensis (DSM 46666 and DSM 45459T) using dbCAN3 server. Only CAZymes predicted by HMMER, DIAMOND and Hotpep tools were considered. AA: auxiliary activities; CBM: carbohydrate-binding modules; CE: carbohydrate esterases; GH: glycoside hydrolases; PL: polysaccharide lyases; GT: glycosyl transferases.
Rare halophilic actinomycetes, such as Actinopolyspora strains, are known for their significant hydrolytic activities and metabolic adaptations to harsh saline environments [2728], which could reflect their bioremediation potential. Bouras et al. [5] studied the ecological diversity and functional capacities of A. saharensis strains based on comparative genomic analyses, focusing mainly on the genetic ability of the type strain H32^T^ (Algeria) and strain BKK2 (China) to produce enzymes (dienelactone hydrolase) capable of degrading petroleum-based plastics. This study showed that the carbohydrate enzymatic activities of A. saharensis strains are highly conserved, despite their ecological diversity. A. saharensis strains have genetic ability to produce potential valuable biocatalyst that may be useful in tackling ecological plastic pollution [4].
Supplementary material
10.1099/acmi.0.001099.v3Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Parte AC Sardà Carbasse J Meier-Kolthoff JP Reimer LC Göker M List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ Int J Syst Evol Microbiol 2020705607561210.1099/ijsem.0.00433232701423 PMC 7723251 · doi ↗ · pubmed ↗
- 2Xie F Pathom-Aree W Actinobacteria from desert: diversity and biotechnological applications Front Microbiol 20211276553110.3389/fmicb.2021.76553134956128 PMC 8696123 · doi ↗ · pubmed ↗
- 3Sabaou N Boudjella H Bennadji A Mostefaoui A Zitouni A et al Les sols des oasis du Sahara algérien, source d’actinomycètes, rares producteursd’antibiotiques Science et Changements Planétaires/Sécheresse 19989147153
- 4Amin DH Abdallah NA Abolmaaty A Tolba S Wellington EMH Microbiological and molecular insights on rare Actinobacteria harboring bioactive prospective Bull Natl Res Cent 202044510.1186/s 42269-019-0266-8 · doi ↗
- 5Bouras N Bakli M Saker R Dif G Meklat A et al Genomic comparison and enzymatic characterization of dienelactone hydrolase in Actinopolyspora saharensis bacteria for petroleum-based plastic bioremediation Arab J Sci Eng 2025211810.1007/s 13369-025-10743-4 · doi ↗
- 6Meklat A Bouras N Zitouni A Mathieu F Lebrihi A et al Actinopolyspora algeriensis sp. nov., a novel halophilic actinomycete isolated from a Saharan soil Extremophiles 20121677177610.1007/s 00792-012-0473-922872369 · doi ↗ · pubmed ↗
- 7Bolger AM Lohse M Usadel B Trimmomatic: a flexible trimmer for Illumina sequence data Bioinformatics 2014302114212010.1093/bioinformatics/btu 17024695404 PMC 4103590 · doi ↗ · pubmed ↗
- 8Bankevich A Nurk S Antipov D Gurevich AA Dvorkin M et al SP Ades: a new genome assembly algorithm and its applications to single-cell sequencing J Comput Biol 20121945547710.1089/cmb.2012.002122506599 PMC 3342519 · doi ↗ · pubmed ↗