Holocene man-occupied caves and transformed wetlands as facilitating factors for Leishmania infantum in South America
Sérvio Pontes Ribeiro, Camila de Paula Dias, Rafael Vieira Duarte, Maria Fernanda Brito de Almeida, Luccas Gabriel Ferreira Malta, José Dilermando Andrade, Alexandre Barbosa Reis, Marcos Horácio Pereira, Carlos EV Grelle, Jesus G Valenzuela, Tiago Donatelli Serafim

TL;DR
This study shows that human-modified environments in South America, especially caves, helped spread a disease-causing parasite by creating habitats for its insect vector and animal hosts.
Contribution
The study links Holocene human activity in caves to the spread of Leishmania infantum through altered ecosystems.
Findings
Caves are present in 18% of Brazilian municipalities and correlate with higher presence of the sandfly vector Lu. longipalpis.
Human and fox populations co-occurred with Lu. longipalpis in cave ecosystems during the Holocene.
Visceral leishmaniasis is shown to be a long-term human-related disease linked to modified environments.
Abstract
In the Holocene, South American humans transformed large extensions of the continent, especially in cave ecosystems. Such transformations produced predictable eutrophic habitats that could have attracted foxes and further favored the adaptation of Lutzomyia longipalpis, insect vector of American Visceral Leishmaniosis (AVL), to human-contaminated habitats. Here we present spatial analyses on the distribution of caves, Holocene human populations, the present-days main wild reservoirs of Leishmania infantum, Cerdocyon thous and Lycalopex vetulus, and the vector Lu. longipalpis in Brazil. The presence or absence of Lu. longipalpis in function of cave abundance, based on coordinates of all recorded samples (Fiocruz and GBIF database and literature), and cave locations taken from ICMBio/CECAV database, were tested by contingency table. The overlap in the distribution of Lu. longipalpis, C.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
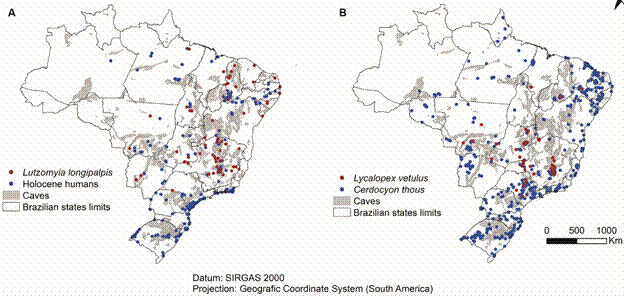 Figure 1
Figure 1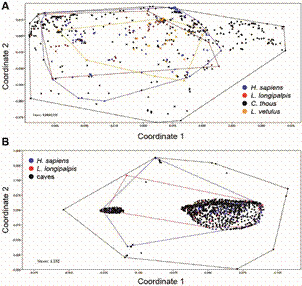 Figure 2
Figure 2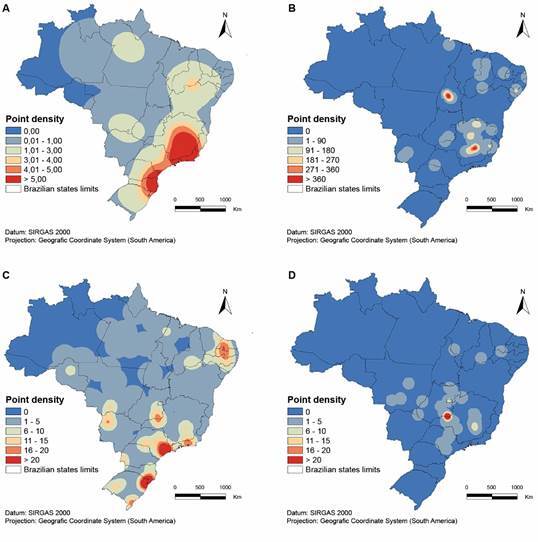 Figure 3
Figure 3 Figure 4
Figure 4- —CNPq
- —FAPEMIG
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsResearch on Leishmaniasis Studies · Environmental Sustainability and Education · Parasites and Host Interactions
Human infectious diseases and parasitism have travelled with people, but with different chances to invade new human populations and habitats. The key to a successful environmental spillover depends on the probability of transmission and host susceptibility,1 but also on how favorable the new environment might be to accommodate ecological demands of a likely invasive parasite. The European human invasion of the Americas introduced a substantial number of new parasites and pathogens to the continent, and some became part of the global pattern of disease distribution.2 However, the ecological conditions that were most favorable for successful parasitism colonization have never been properly explored.
Host species spatial distribution, abundance and habitat fidelity are key factors in host-parasite ecological dynamics, affecting transmissibility and evolution of virulence.3 In scenarios where tolerance (being infected without getting sick) is more likely to evolve than resistance (not getting or maintaining the infection), would have a positive effect on a parasite population that has spilled over, allowing it to spread silently. In cases of a recurrent and predictable interaction between host and parasite, tolerance of a proportion of the host genotypes may boost parasite specialization.4 ^,^ 5 Such a pattern is typically associated with plant-pathogen systems, but animals with low vagility and high habitat fidelity could produce a similar evolutionary scenario.5 As settled and landscape-attached human communities, we became a long-term predictable host, and this may have been a driver of parasite spillovers, namely of those parasites introduced by migrant humans into native humans and animals.
The probability of encountering a susceptible host might favor generalism in an ecological community dominated by either spatially unpredictable, highly vagile or rare host species.5 ^,^ 6 Conversely, the predictability of successfully finding a susceptible host could result in specialization to one or a few hosts species.7 Applying this concept to vector-borne diseases, the prevalence among humans depends, first, on the distribution of the vector species. Hematophagous insect dispersal is driven by habitat suitability and host density and distribution.8 Possible trade-offs between surviving in certain environments (especially for immatures) and optimizing egg production (depending on finding predictable blood sources) might affect oviposition choices, dissemination behaviors, as well as the evolution of host species range.8 ^,^ 9 This combination of optimal habitats and the predictability of human settlements may have been shaping the evolution of human parasites since we abandoned nomadism.
A vertebrate animal species regularly associated with a place, such as dens and territorial dwellings, leaves environmental signals to hematophagous insects about their location. In the case of humans, predictability could be a main driver for insect vector of diseases to specialize in anthropomorphic environments and domestic animals.7 ^,^ 10 Further, a disease transmitted by insect vectors with a highly specialized infection route, and with a substantial proportion of asymptomatic hosts, is likely a well-established specialization.3 ^,^ 11 The above description fits the leishmaniasis, especially the visceral form found in wild canids, domestic dogs and humans, in both the Old and New World.12 ^,^ 13
An aspect rarely discussed in visceral leishmaniasis ecology is that human and other large animals’ interactions might have created richer and more predictable microhabitats for sandfly immatures. However, a limited number of sandfly species can transmit visceral leishmaniasis in both Old and New World, compared to cutaneous leishmaniasis.13 These are the same few sandfly species associated with modern densely human populated suburbs. Ecological filters such as highly toxic or polluted environments found around our settlements could reduce the chance of many species associating with us.14
Many distinct canid species have experienced convergent domestication processes throughout the history of humanity, dating back 40,000 years ago (y.a. hereafter) in North America,15 ^,^ 16 but also in Egypt (7,000 y.a.), Iraq (8,000 y.a.), Cyprus (5,500 y.a.), Greece (6,000 y.a.)17 ^,^ 18 and also in South America (2,000-3,000 y.a.19 ^,^ 20 In South America, the 10 living canid species are spread out along the whole continent, and are descendants of a sole ancestral species, which invaded the continent between 3.9 million years ago (Mya hereafter) and 3.5 Mya,21 just before the sandfly genus Lutzomyia is supposed to have arrived, both through the Panama Isthmus.13 ^,^ 21 In South America, American visceral leishmaniasis (AVL hereafter) is caused mostly by Leishmania infantum, and transmitted by Lutzomyia longipalpis (Psychodidae; Phlebotominae). Two fox species in South America, Cerdocyon thous and Lycalopex vetulus are listed as reservoirs of L. infantum.22 ^,^ 23 Regardless ongoing debate, the most accepted hypothesis is that at least part of AVL is caused by a sub-population of L. infantum emerged from a Portuguese population, likely arrived around the colonization time, 500 y.a.24
Considering a spillover of L. infantum from Old World dogs to South American fox species implies that the common vector, Lu. longipalpis, could have blood-fed on recent invaded domestic dogs, as well as associated to their domestic environment. A recent study has shown that domestic dog feces are a suitable substrate for the full development of Lu. longipalpis.25 Canid dens are composed of feces and prey corpses under distinct decaying stages, resulting is a very special organic matter-rich microhabitat.26
On the other hand, Forattini27 described Lu. longipalpis as a species constrained to rocky-shelters, caves or deep creek valleys within the savanna or dry forest Biomes. Hence, this species is constrained to very wet and predictable microhabitats in ecosystems submitted to a seasonal climate, with very dry winters, when low relative air humidity predominates. Whether a cave also provided shelter for foxes and humans in the past, it became a wet but also nutrient-rich environment. Therefore, a resource and climatic favorable and predictable microhabitat.
South America had a fast and diverse dispersion of humans from 13,000 y.a.,28 ^,^ 29 followed by a stable, relatively low population around a carrying capacity from 9,000 to 5,500 y.a.30 Most of these stable occupations were along the coast, in the Amazon or in the cave-rich landscapes of central highlands.30 ^,^ 31 Later, from 5,5000 to 2,0000 y.a., new waves of migrations, combining a complex movement of linguistic groups spread and mixed with original groups. First, Jê-speaking groups, which mixed and changed Sambaqui-builders’ traditions in the coast.31 After, the Tupi-Guarani populations, which occupied and transformed most of the large riverbeds and wetlands of the continent.30 ^,^ 31 ^,^ 32 An unknown aspect is whether such largely human-transformed landscapes could favor population growth of both vectors and reservoirs of parasitic human diseases.
The fast change in the carrying capacity from 5,500 y.a., followed by an exponential growth of continental and coastal populations, could only be possible by technological changes in the management of the landscape, most likely related to a new wave of Tupi-Guarani and related groups around the continent.30 ^,^ 33 Such land management with fast population growth was most likely related to sedentarism, and overlapping with previously existent cave people.28 Such possible scenario must have caused sensitive transformations in the habitats, making them more eutrophic, as usually happen to human settlements. Therefore, the environment also may have become attractive to commensal canids,16 including South American foxes.20 ^,^ 34 Whether habitat enrichment and the constant presence of these interactive hosts might have driven Lu. longipalpis to adapt for living near humans is currently under investigation.
Neither the existence of pre-European complex societies, densely distributed human populations,28 ^,^ 30 nor the indigenous likely domestication (or at least taming) of foxes20 were taken into consideration to describe the natural history of visceral leishmaniasis in South America. Assuming the most likely scenario, that L. infantum invaded the continent with Portuguese infected dogs, there is a lack of studies on how such spillover to native foxes and people may have happened. To understand whether there was pre-colonial human-made niche for this parasite to invade could help to understand how this disease later migrated to suburbs of modern cities in the continent.
Here we explored the hypothesis that Lu. Longipalpis’ distribution may overlap with the distribution of caves occupied by human populations from 9,000 y.a., being its present-days distribution a trace left from a pre-historic ecological association. Further, we also evaluated whether fox species which are implicated as wild reservoirs of AVL, overlap with Lu. longipalpis and early South America human distribution. We further discuss the present distribution of the disease and Lu. longipalpis in wild areas that were, until recently, the home of large native human populations and explored ecological paths for this sandfly to have invaded urban ecosystems.
MATERIALS AND METHODS
Study area - South America has a diverse and abundant number of caves.35 According to governmental survey, Brazil has documented caves in 995 municipalities (Available from: https://www.gov.br/icmbio/pt-br/assuntos/centros-de-pesquisa/cecav/), which correspond to 18% nationwide number of municipalities. Rocha et al.36 pointed out a total of 16,089 known caves, 68% of them are limestone caves (Brazilian Karst), and 49.7% found in the savanna (Brazilian Cerrado) vegetation, where Lu. longipalpis predominates, along with the two fox species (C. thous and L. vetulus) incriminated as reservoirs for AVL. Likewise, according to Goldberg et al.30 kernel density map of humans during the Holocene in South America (13,000 to 4,000 BP), their occurrence overlapped quite substantially with biogeographic regions with an abundance of caves.
Although in modern times most Lu. longipalpis samples are found in urban areas,37 we hypothesize them to also happen in urban locations with similar ecological conditions of cave hidden den-like habitats, i.e., city locations with low to no sanitary infrastructure with spots of soil and water contaminated with rich organic matter, from dog feces to open air sewage. Although any urban’s neglected, wet, dirty, and sheltered dwelling spaces can provide ideal breeding grounds for this species, low-income neighborhoods in the vicinities of cave-rich ecosystems might have greater chances of having long-term endemicity of AVL.
To test this hypothesis, we first verified whether Lu. longipalpis distribution might be restricted to those biogeographic regions rich in caves, testing presence or absence of the sandfly in function of cave abundance. We took the coordinates of all recorded samples of Lu. longipalpis in the Fiocruz database (Available from: https://specieslink.net/search/ samples from 1956 to 2015), double checked in the free and open access to Biodiversity database (Available from: https://www.gbif.org/). These two databases comprise the majority of scientific sampling of Lu. longipalpis in the country. Other samples from health service were not used as identification precision may vary excessively. On the other hand, cities with confirmed occurrence of the species in the literature and not recorded by the Fiocruz’s or GBIF database were also added (taking the core coordinate of the city) to decrease database record bias. The known cave locations and estimated locations were taken from ICMBio/CECAV database (Available from: https://www.gov.br/icmbio/pt-br/assuntos/centros-de-pesquisa/cecav/).
We assumed municipality as territory unit, but also, we corrected by neighbor caves. For each occurrence of Lu. longipalpis, we associated the number of caves in that municipality, and then checked by actual cave proximity, matching each sampling location with any actual cave occurrence within a radius of 100 Km from the center in the coordinates of Lu. longipalpis sample point, using Google Earth distance tools. This area size was defined arbitrarily, but in order to deal with the wide range of municipality sizes in the Country [from 3.5 to 60,431 Km^2^ in Southeast and Northeast regions (Available from: https://www.ibge.gov.br/geociencias/)]. However, this correction was made only to prevent any underestimation of cave density in situations in which a Lu. longipalpis sample could have been taken in a municipality out of core distribution of caves, but still close enough to cavities. Then, caves found in neighboring municipalities within this radius were summed up for that sandfly occurrence.
In order to avoid inflating the zeros, we excluded the municipalities belonging to Biomes unlikely to have Lu. longipalpis, namely the wet Amazon forests and subtropical South of Brazil. These are also regions with irrelevant public records of AVL. All remaining Brazilian municipalities with no sampling or confirmed records of Lu. longipalpis were added as a “zero” occurrence. Because a reasonable proportion of zeros might be due to lack of research rather than actual absence of the species, we validated the distribution of Lu. longipalpis, and verify the effect of possible under sampling, by overlapping the distribution of the insect with the distribution of AVL incidence of cases in the Country, taken from the Information System of Injuries Notification (SINAN), Ministry of Health, from 2001 to 2021.
We then tested the overlap in the distribution of Lu. longipalpis, C. thous and L. vetulus with humans from Holocene. Holocene human distribution came from data made available in Goldberg et al.30 The foxes species data were obtained from ICMBio/Sistema de Avaliação do Risco de Extinção da Biodiversidade - SALVE (Available from: https://salve.icmbio.gov.br/) - accessed in 28/04/2023.
Kernel density maps were produced for each species based on the distribution in the database mentioned above, and on the Goldberg et al.30 published database. All distributions were overlap with projections of high potential for the occurrence of caves in Brazil, found in Jansen et al.35 Maps were produced in Arcmap 10.3 using default search radius for sandfly and foxes, and a search radius of 660 Km for Holocene humans.
Lutzomyia longipalpis distribution in function of proximity of caves was tested with a Contingency Table using MINITAB v21.3. The co-occurrence of the four species (humans, the two foxes and Lu. longipalpis) and the caves were tested by permutational multivariate analysis of variance (PERMANOVA). For such, we built a nonmetric multidimensional scaling (NMDS) analysis for all four species together and also for Lu. longipalpis and humans separate, using PAST v4.13. Using a simple dissimilarity matrix-based and non-metric method, it is possible to take coordinates as environmental variables and so scale up species by similarity in their location.38 ^,^ 39 The PERMANOVA compares groups of species and tests whether their distribution centroids and data scattering in the multidimensional space were significantly different.38 ^,^ 39
RESULTS
Lutzomyia longipalpis was found in only 65 municipalities. Regardless apparent under sampling, these sites were widespread all over the country. The species was significantly more recorded in municipalities with caves or at least 100 Km near a cave in a neighboring municipality (Chi-Square = 21.58, p < 0.0001, d.f. = 1). Within the biogeographic range of present days AVL incidence, which is mostly in the Biomes of Cerrado, Atlantic Rainforest, and Dry Forests, municipalities with mapped caves were 767, summing up 18% of total. On the other hand, 86% of database-deposited records of Lu. longipalpis came from these same 767 municipalities with caves.
Conversely, from all municipalities with caves (882), 70.2% of them had recent records of AVL cases, which composed 21.3% of the total municipalities with AVL (2910 out of 5565 Brazilian municipalities). Still, the majority of the 52% Brazilian municipalities which had AVL in present days happened within the Biomes above cited.
Also, this sandfly species showed a significant overlap distribution with Holocene humans and the two foxes species incriminated as reservoirs of L. infantum, C. thous and L. vetulus (Fig. 1). Both foxes´ distribution were far broader than Lu. longipalpis or Holocene humans, as they occupy a much larger territory, besides the cave-rich biogeographic regions (PERMANOVA F = 3.34, p <0.007, Fig. 2).
Fig. 1:distribution of Holocene humans, Lutzomyia longipalpis (A), Cerdocyon thous and Lycalopex. vetulus (B) overlap with the expected distribution of caves in Brazil. Coordinates of all recorded samples of Lu. longipalpis in the Fiocruz database (Available from: https://specieslink.net/search/ samples from 1956 to 2015), double checked in the free and open access to Biodiversity database (Available from: https://www.gbif.org/), and the known cave location and estimated locations from Instituto Chico Mendes de Conservação da Biodiversidade/Centro Nacional de Pesquisa e Conservação de Cavernas (ICMBio/CECAV) database (Available from: https://www.gov.br/icmbio/pt-br/assuntos/centros-de-pesquisa/cecav/). The foxes data were obtained from ICMBio/Sistema de Avaliação do Risco de Extinção da Biodiversidade - SALVE (Available from: https://salve.icmbio.gov.br/), accessed in 28/04/2023. Holocene human distribution came from available data in Goldberg et al.30
Fig. 2:non-metric multidimensional scaling (NMDS) from coordinates of occurrence of Holocene humans, Lutzomyia longipalpis, Cerdocyon thous and Lycalopex vetulus (A) and occurrence of Holocene humans, Lu. longipalpis and caves (B). The coordinate axes are collapsed information from original multivariate factors, in this case, the geographical actual coordinates for each location where species were recorded. These combined coordinates created by rank orders allow statistical comparison of co-occurrence.
Then, the exclusion of the foxes from the NMDS/PERMANOVA analyses exposed a non-significant difference in the distributions of present days Lu. longipalpis and Holocene humans and caves (PERMANOVA 0.51, p > 0.5, Fig. 2). Still, the distribution of all four species strongly suggest that coexistence must have happened among them in many locations in the central Brazil, with two especially strong hotspots, in interior Northeast Region and Central Minas Gerais State, and for quite a long time (Fig. 3).
Fig. 3:kennel maps for the Holocene humans (A), Lutzomyia longipalpis (B), Cerdocyon thous (C) and Lycalopex vetulus (D) overlap with the expected distribution of caves in Brazil. Point densities show the regions with the highest to the lowest occurrence of each studied event.
DISCUSSION
Visceral leishmaniosis is a disease that evolved through an ecological path completely different from the cutaneous form. While cutaneous leishmaniasis is caused by more than 20 Leishmania species (Available from: https://www.cdc.gov/dpdx/leishmaniasis/index.html)[13](#B13) VL is caused by only a few species. The most common are L. infantum, L. donovani 13 ^,^ 40 and, sporadically, L. tropica 41 and L. amazonensis.42 There are also quite a few sandfly species capable of transmitting L. infantum or L. donovani. In the New World: Lu. longipalpis, secondarily L. cruzi,43 and sporadically found in other species such as Migonemyia migonei, L. evansi, Nyssomyia whitmani or species of Evandromyia.13 ^,^ 44 ^,^ 45 ^,^ 46 ^,^ 47 In the Old World: Phlebotomus perniciosus, P. major, P. chinensis, P. longicuspis and few other sporadic species.48 ^,^ 49
The geographic distribution of Lu. longipalpis and AVL reservoir foxes in Brazil showed a clear association of these species with cave habitats where, in the past, human had a consistent occupation. For instance, the oldest South America skeleton, Luzia, 10,000-year-old women, was found in a cave in Lagoa Santa Municipality, Minas Gerais State, a region abundant in caves and endemic for AVL. Fox dens are found in the crevices and holes around and in the entrances of caves. Nearby Luzia’s cave (Lapinha cave), we sampled Lu. longipalpis females around feces in a fox den (Fig. 4). For the crab-eating fox, C. thous, the proximity to humans seemed not to be a problem even in present days, but an opportunity of resources, as described for Indian free ranging dogs.50
Fig. 4:surrounds of Lapinha Limestone cave, showing several fox dens or latrines, which are open to outside.
Lutzomyia longipalpis invaded South America by the same time the ancestor canid, likely a direct ancestor to the Lycalopex genus or a Cerdocyon species.13 ^,^ 21 The accepted scenario is that canids spread thru the continent following xeric habitats, as savannas (and, thus, an ecological filter), by the time the Panamanian connection was formed between North and South America.21 A savanna route could have been decisive for the widespread and radiation of most South American canids around 1.0 Mya, when dry Pleistocene periods made savannas predominant in the whole continent.21 ^,^ 51 The existence of some level of mesic microhabitats around dens, eventually within gallery/deep valley forests, would have been a very favorable habitat for Lu. longipalpis immature development.27 ^,^ 52
Humans arrived much later, and likely due to climate unpredictability during the Younger Dryas (from 13,074 to 11,775 y.a.), started to occupy sheltered habitats as caves.28 ^,^ 53 Peopling around caves were intensified in early Holocene, from 10,000 to 9,000 y.a.28 Thus, our hypothesis is that humans stepped into this fox-sandfly habitat, transformed it into a much more eutrophic, plenty of predictable organic matter spots, to which Lu. longipalpis may have quickly adapted.
As humans and foxes are carnivores, an environment abundant in iron-rich feces and prey corpses under distinct decaying stages are a likely output for this interaction, as already recorded by other hunter gathering populations and canid dens.16 ^,^ 20 ^,^ 26 Thus, they might produce a blood-rich organic matter, highly contaminated by iron, especially due to Heme group decomposition, which releases Fe^+3^, a more toxic form of this element. Hence, human and fox interaction could produce an environmental filter for decomposers to overcome, a likely explanation for the existence of so few sandfly species associated with them. Ribeiro54 showed that Lu. longipalpis has oviposition preference for decaying rat corpses and fresh dog feces, if compared with usual substrates tested in the literature. Also, it was found that this species can complete its life cycle on such substrates, showing an eco-evolutionary basis for this sandfly proximity with city suburbs.
Tupi-Guarani populations expanding towards cave-rich biogeographic regions, coming from mid-Amazonia, and then overlapped in time and space with original cave populations,32 may have created ecological conditions for Lu. longipalpis to follow humans up to a new, technologically transformed habitat. Intense human constructions for ritualistic burial, such as the coastal Sambaquis, must have created an extra habitat very rich in decaying corpses and wet microhabitats. Most of these burials were sequential, resulting in a great number of bodies close together in decomposition, summed up to offerings of dead animals,55 ^,^ 56 which must have resulted in organic matter-rich habitats.
A location with a well understood native civilization history, and where AVL cycle was properly studied is the Marajo Island.12 The Marajo is an immense 40,100 Km^2^ Island formed by the delta of the Amazonia River. It is a floodplain covered by savanna vegetation, and it was originally occupied by a complex native society, quickly exposed to European invaders in the early XVI Century, due to the proximity to Belém city.
Lainson & Rangel,12 after Lainson et al.,23 supported the hypothesis that C. thous was the sylvatic reservoir of the AVL, which infected dogs, and then humans, in small rural farms close to primary forests. This was a well-accepted hypothesis, built from a sample in the Marajo Island, in Salvaterra village, a location near a so-called primary forest. However, being the Island highly populated by indigenous before Europeans colonization, the possibility of a pristine, primary forest is remote.57 Here we offer an alternative interpretation to Lainson’s pioneering and important findings.
In 5,500 y.a., the ancestral of Jê and Tupi-guarani groups, especially the latter, were able to deal with agriculture in the semi-arid, seasonal environments, and occupied wetlands from the Pantanal Matogrossense to Marajo Island.30 ^,^ 32 ^,^ 58 These people built mounds of large sizes to perform symbolic burial, as well as to manage water flow.59 Their migration transformed the landscape in the whole South America, even the old, occupied lowlands around central plateau caves, and civilizations overlap is likely to have happened.31 ^,^ 32
The Marajo Island was probably colonized once, around 3,100 y.a. At the Marajoara phase the island could have reached up to 1 million people,60 with the summing up of villages and cities at the denser regions capable of accommodating up to 100,000 people.57 The Marajoara phase was a complex society of mound constructers and pottery makers, with intense landscape management and urban structures that lasted from 1625 to 668 y.a. (400 to 1350 AD).59 These human-modified, and organic matter-rich habitats could have become a substitute for the human-foxes cave habitats for Lu. longipalpis. Therefore, a recent adaptation of Lu. longipalpis to peri-urban and suburb habitats may never have happened from populations coming from the wild, but from insects ecologically adjusted to primitive human-modified habitats, previously found in the same regions where the species has been spreading now-a-days.
The system visceral leishmaniasis-canines-humans may have evolved separately and in the north Africa and Mediterranean Europe,24 but a convergent phenomenon is possible to have happened in the Neotropical region.61 In any case, for this host-parasite system to thrive, habitat-specific insect vector species also should have to adapt to the habitats created by humans, as well as by canine packs. In both cases, heavily eutrophic dens, caves, and primitive human-made litter and ceremonial mounds, rich in feces and carcasses, may have shaped the environments around these vertebrate species (and also, may have been a main driver of canid domestication),16 imposing specific selection on insects associated to such extreme conditions. There are reports on different Nations of Guarani ethnic groups who used to bury domesticated foxes of different species, highlighting the human-canid domestication relationship, which might happen to suffer hybridizations with European dogs later on.15 ^,^ 34
Ongoing research of our group has shown that Lu. longipalpis immatures fed on dog feces exhibit high ferritin (a decontaminating protein) transcription levels, similar to immatures exposed to iron-contaminated substrates. Hence, iron and other heavy metals found in organic matter such as carnivore feces and carcasses, act as a strong ecological filter, limiting sandfly diversity associated with canids. This is an avenue for future research.
In 1620, the Marajo Island was still heavily populated by native communities, but also threatened by a combination of Portuguese, Dutch and English invaders, based on both official forts and settlements, as well as pirates.62 Mostly important, the Marajoara society (Nheengaíba nation) survived and resisted for 20 years a war promoted by the Portuguese.62 ^,^ 63 Such war finished only when Jesuit missionaries were allowed into the island, promoting a partially pacific annexation, in 1659.63
The prolonged conflict likely created opportunities for VL infected domestic dogs to reach the island and get in contact with native foxes, in case introduction was indeed the pathway for the disease to spillover to native hosts. On the other hand, in the possibility of a native origin of L. infantum,61 it is still possible to consider an evolutionary scenario of a four-species interaction, based on expected L. infantum’s niche demand. The conditions into which L. infantum spread and keep viable populations seemed to depend on: 1) highly contaminated and humid microhabitats prone to the insect vector domestication (likely to have been colonized by Lu. longipalpis, after adaptation to heavy metal contaminated organic matter); 2) densely human occupied and transformed landscapes.
The human-made habitats in South America during the late Holocene likely are convergent and similar to those in the Old World´s hotspots for visceral leishmaniasis, namely, North Africa-South Europe, Mid-East and Southeast Asia.17 All these regions are expected to have been densely populated and transformed in the late Holocene.28 ^,^ 32 Such worldwide similarities in that stage of humanity evolution are also described as a baseline for diversification and dispersion of parasitic diseases, as well as canid domestication.18 ^,^ 19
In conclusion, our findings indicate that Lu. longipalpis may have adapted to living near humans long before European colonization. Recent archeological evidence suggests that original South American societies transformed dramatically the landscape, creating vast anthropomorphic environments,64 likely to favor insect hematophagous as well as to create a landscape prone-to-transmission of diseases. The proximity of foxes to indigenous settlements may have further enhanced the creation of hyper-wet and nutrient-rich microhabitats, which are suitable to Lu. longipalpis immature development, around human occupied caves, as well as in primitive constructions in wetlands. Our species distribution data showed a very clear habitat-related co-occurrence of human-foxes-Lu. Longipalpis, providing solid, statistically supported, evidence for our conclusion.
The most relevant implication of our findings is that AVL, and also Old World VL, should be considered a long-term human related disease, associated with few sandfly species well adapted to our modified, and heavily contaminated, environments. Hence, and contrary to cutaneous leishmaniasis, this is not a sylvatic disease spilling-over to human societies. The narrative that the AVL invaded cities from wild ecosystems, under this perspective, should be revisited. Lu. longipalpis (as well as the C. thous fox), may have kept associated from old to new human settlements, especially in cave-rich biogeographic regions. Further studies on Lu. longipalpis immature development on habitats related to human-canid contaminated organic matter, as well as the identification of neglected urban habitats capable of retaining similar humidity and dirtiness like those in primitive caves, should be put in priority for disease control.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Karesh WB Dobson A Lloyd-Smith JO Lubroth J Dixon MA Bennett M Ecology of zoonoses natural and unnatural histories Lancet 201238098571936194510.1016/S 0140-6736(12)61678-X 23200502 PMC 7138068 · doi ↗ · pubmed ↗
- 2Murray KA Preston N Allen T Zambrano-Torrellio C Hosseini PR Daszak P Global biogeography of human infectious diseases Proc Natl Acad Sci USA 201511241127461275110.1073/pnas.150744211226417098 PMC 4611664 · doi ↗ · pubmed ↗
- 3Dutra DA Poulin R Ferreira FC Evolutionary consequences of vector-borne transmission how using vectors shapes host, vector and pathogen evolution Parasitology 2022149131667167810.1017/S 003118202200137836200511 PMC 10090782 · doi ↗ · pubmed ↗
- 4Antonovics J Thrall P Jarosz A Stratton D Real L Ecological genetics Ecological genetics of metapopulations: the Silene-Ustilago plant-pathogen system 1994 Princeton Princeton University Press 146170
- 5Höckerstedt L Numminen E Ashby B Boots M Norberg A Laine AL Spatially structured eco-evolutionary dynamics in a host-pathogen interaction render isolated populations vulnerable to disease Nat Commun 2022136018601810.1038/s 41467-022-33665-336229442 PMC 9561709 · doi ↗ · pubmed ↗
- 6Leggett HC Buckling A Long GH Boots M Generalism and the evolution of parasite virulence Trends Ecol Evol 2013281059259610.1016/j.tree.2013.07.00223968968 · doi ↗ · pubmed ↗
- 7Powell JR Gloria-Soria A Kotsakiozi P Recent history of Aedes aegypti vector genomics and epidemiology records Bio Science 2018681185486010.1093/biosci/biy 11930464351 PMC 6238964 · doi ↗ · pubmed ↗
- 8Rose NH Sylla M Badolo A Lutomiah J Ayala D Aribodor OB Climate and urbanization drive mosquito preference for humans Curr Biol 202030183570357910.1016/j.cub.2020.06.092.932707056 PMC 7511451 · doi ↗ · pubmed ↗