Beyond kin killing: Dickeya-derived phage-tail-like bacteriocin P2D1 targets phylogenetically distant Pseudomonas spp
Marcin Borowicz, Jan Styn, Kacper Tomasik, Łukasz Rąbalski, Magdalena Narajczyk, Erwan Gueguen, Sylwia Jafra, Julie Baltenneck, Dorota M Krzyżanowska, Robert Czajkowski

TL;DR
A bacteriocin from Dickeya dadantii can kill distant Pseudomonas species, challenging the idea that such weapons only target close relatives.
Contribution
Demonstrates that the tailocin P2D1 targets phylogenetically distant Pseudomonas species, expanding its ecological relevance.
Findings
Nine Pseudomonas strains from diverse clades were found to be susceptible to P2D1 tailocin.
A D. dadantii mutant lacking tailocin genes lost the ability to kill Pseudomonas isolates.
Some P2D1-susceptible Pseudomonas strains reduced D. dadantii-induced soft rot on potato.
Abstract
Tailocins, phage-tail-derived bacteriocins, are increasingly recognized as potent mediators of microbial antagonism, yet their ecological scope beyond kin-targeting remains poorly understood. Here, we investigated whether P2D1, a tailocin produced by the plant pathogen Dickeya dadantii 3937, can act against environmental bacteria phylogenetically distant from Dickeya spp. Screening 480 soil and rhizosphere isolates from three distinct plant-associated habitats in Poland, we identified nine Pseudomonas spp. strains susceptible to tailocin P2D1. Whole-genome sequencing and phenotype profiling revealed that these isolates spanned multiple clades, including taxa related to P. germanica, P. tensinigenes, and P. parakoreensis. The D. dadantii mutant lacking genes encoding tailocin sheath and tube proteins lost antagonistic activity against Pseudomonas isolates, confirming that tailocins alone…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
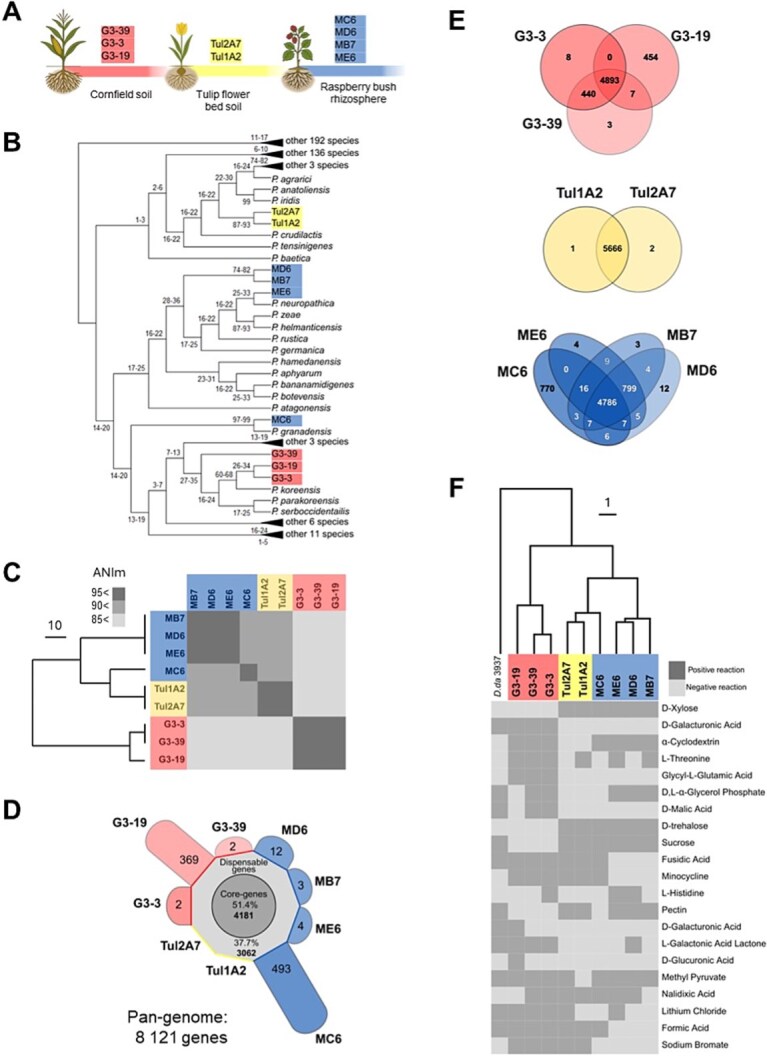 Figure 1
Figure 1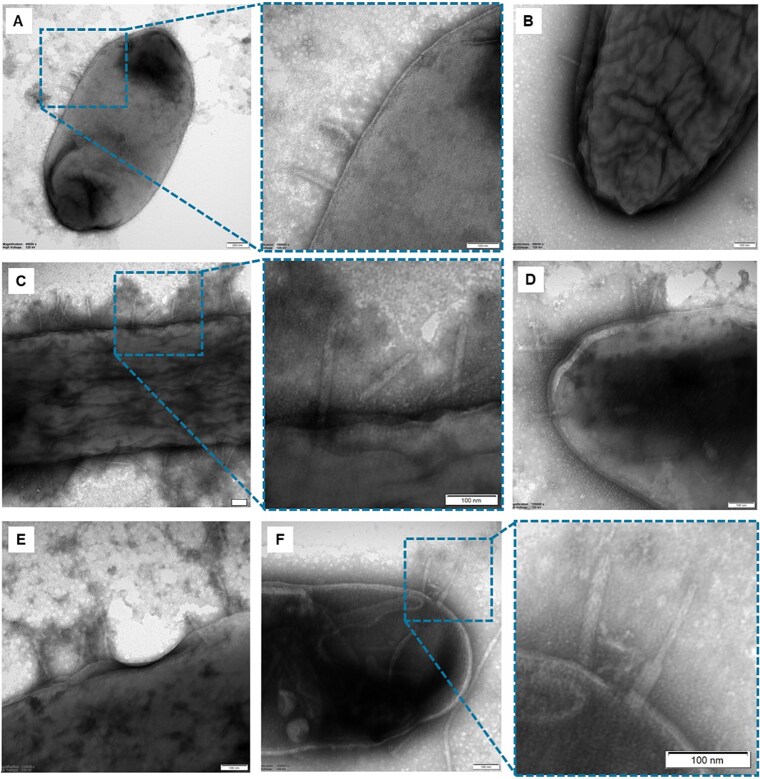 Figure 2
Figure 2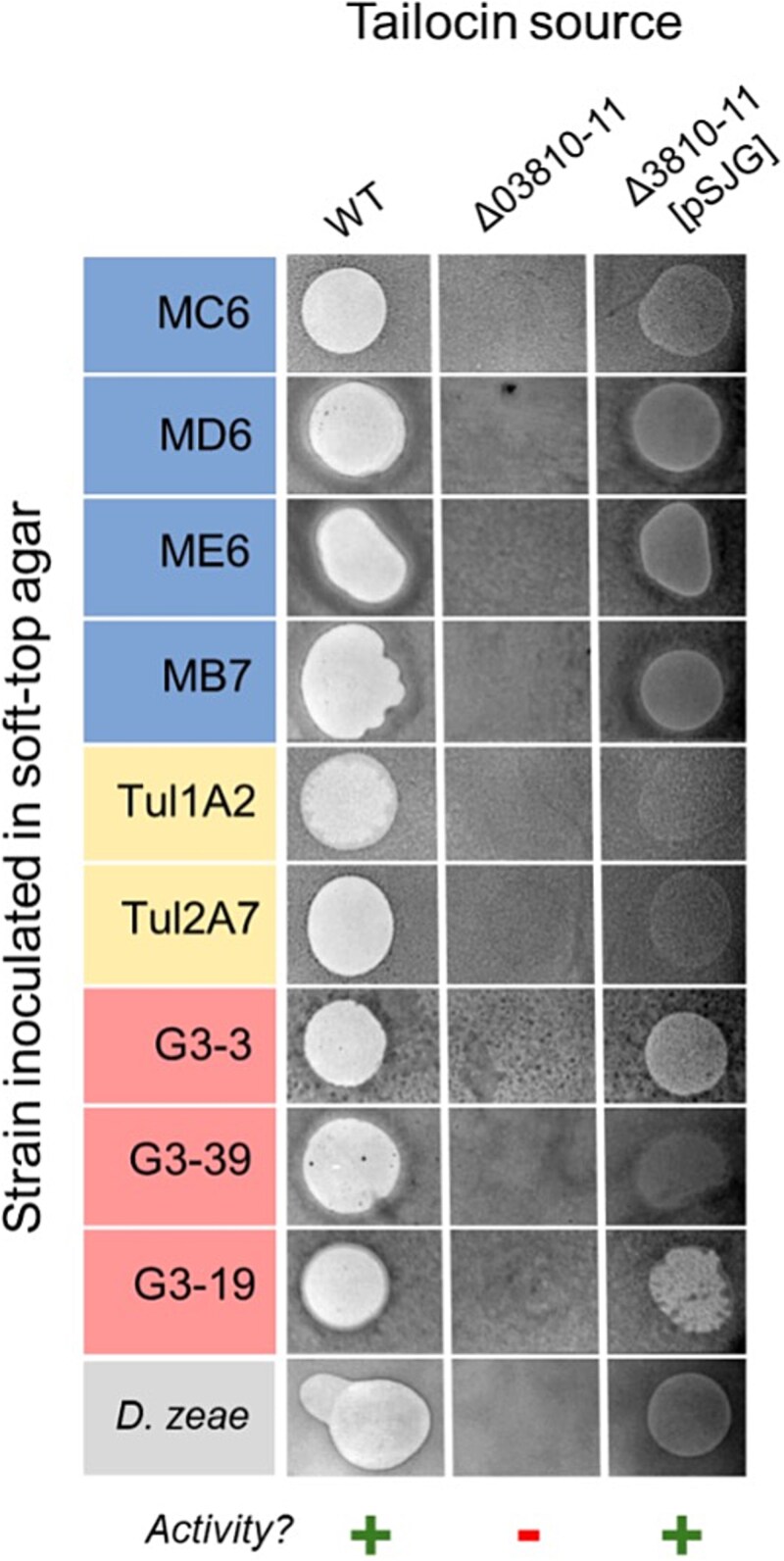 Figure 3
Figure 3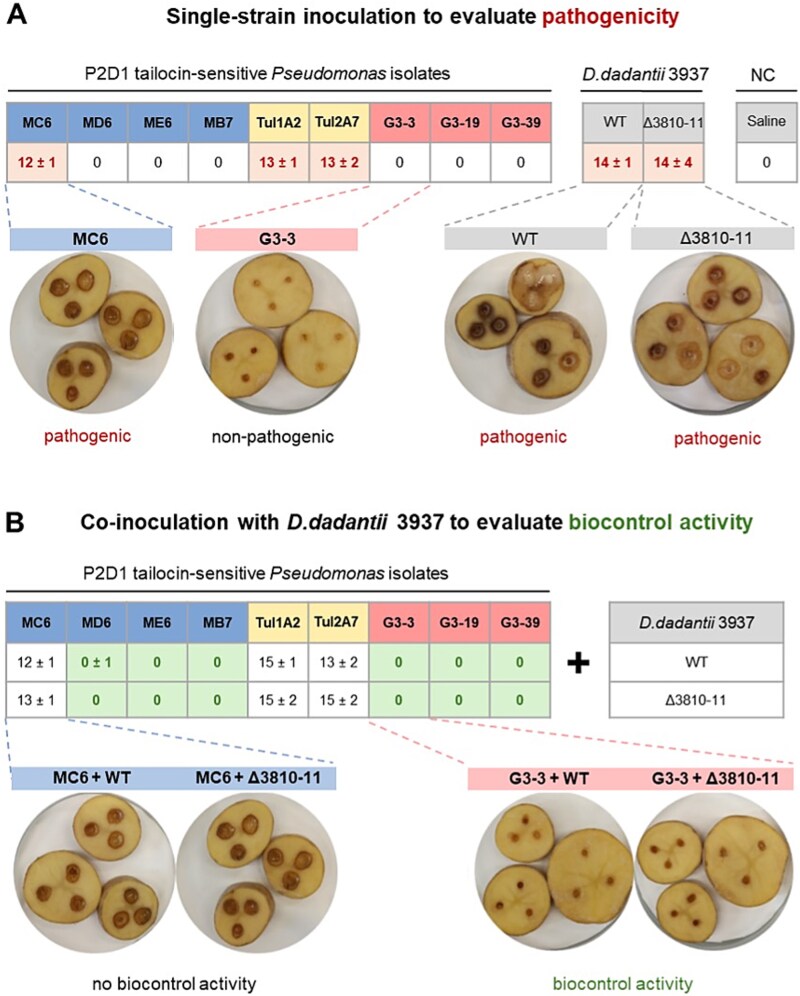 Figure 4
Figure 4| Bacterial strain | Description | Source |
|---|---|---|
|
| ||
| DH5α |
| Laboratory collection |
| DH5α λpir | λpir phage lysogen of DH5α | Laboratory collection |
| MFD |
| [ |
|
| ||
|
| Wild type, a representative member of Soft Rot | Laboratory collection |
| Δ3810 | ∆ | This study |
| Δ3811 | ∆ | This study |
| Δ3810-11 | ∆ | This study |
| Δ3810-11 [pSJG] | Double mutant Δ3810-11 carrying the complementation vector pSJG | This study |
|
| ||
| MC6 | A set of strains isolated from raspberry bush rhizosphere in Białogóra, Poland, 2024 (5 481 533°N, 1796580°E); susceptible to P2D1 tailocins from | This study |
| MD6 | ||
| ME6 | ||
| MB7 | ||
| Tul1A2 | A set of strains isolated from tulip flower bed rhizosphere in Gdynia Poland, 2024, (5 450 774° N, 1854757° E); susceptible to P2D1 tailocins produced by | This study |
| Tul2A7 | ||
| G3-3 | A set of strains isolated from cornfield soil in Częstochowa, Poland, 2024 (5 095 110° N, 1922670° E); susceptible to P2D1 tailocins produced by | This study |
| G3-19 | ||
| G3-39 | ||
|
| ||
|
| Member of the Soft Rot | Laboratory collection |
| Isolate | Source | Genome size (Mbp) | CDS | GenBank accession no. | Closest related type | ANIb (aligned sequence), (%) | ANIm (aligned sequence), (%) |
|---|---|---|---|---|---|---|---|
| MC6 | Rabsberry bush rhizosphere | 6.53 | 5761 |
|
| 91.86 (87.22) | 93.04 (86.76) |
| MD6 | 6.49 | 5759 |
|
|
|
| |
| ME6 | 6.49 | 5755 |
|
|
|
| |
| MB7 | 6.49 | 5752 |
|
|
|
| |
| Tul1A2 | Tulip flower bed soil | 6.63 | 5788 |
|
|
|
|
| Tul2A7 | 6.63 | 5787 |
|
|
|
| |
| G3-3 | Cornfield soil | 6.12 | 5451 |
|
| 92.42 (84.64) |
|
| G3-19 | 6.19 | 5491 |
| 92.38 (83.89) | 93.36 (85.78) | ||
| G3-39 | 6.12 | 5448 |
|
| 92.45 (84.51) | 93.45 (85.83) |
- —University of Gdansk
- —National Science Center, Poland (Narodowe Centrum Nauki, Polska)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogenic Bacteria Studies · Bacterial biofilms and quorum sensing · Plant-Microbe Interactions and Immunity
Introduction
Microbial communities in natural environments are shaped by intense competition for resources, with bacteria deploying a diverse array of strategies to outcompete their neighbors [1, 2]. Among these competitive factors, particularly intriguing yet poorly understood players are tailocins [3]. These phage tail-like bacteriocins are contractile nanomachines that, due to their structural similarities, are believed to be evolutionarily associated with bacteriophage tails [4]. Tailocins employ a single-hit killing mechanism facilitated by high-affinity recognition and membrane penetration in target bacterial cells, resulting in the rapid depolymerization of the susceptible cell and its ultimate death [5]. While tailocins production occurs across diverse bacterial species, their ecological role remains to be fully elucidated.
Tailocins are currently regarded as mediators of intraspecific competition, promoting kin killing through their specificity toward closely related strains, a context in which their activity has predominantly been characterized to date [6]. However, several studies have reported tailocins capable of acting across taxonomic boundaries [7–11], suggesting that their ecological role may be broader than previously assumed. This shift in perspective is particularly relevant in densely populated, taxonomically diverse microbial environments, where microorganisms continuously compete for limited space and resources [12, 13].
Soft rot Pectobacteriaceae (SRP), including Dickeya spp., are well-characterized plant-associated bacteria known to produce tailocins [14]. We recently described a novel tailocin, dickeyocin P2D1, produced by D. dadantii strain 3937. Genetic clusters, such as those that encode the P2D1 tailocin, have been found to be widely distributed across Dickeya spp. [14, 15]. This suggests that P2D1 may play a significant ecological role in these bacteria, potentially conferring a competitive advantage within infected plant tissues and in other niches from which these bacteria are commonly isolated, such as soil, the rhizosphere, and aquatic environments [16, 17]. To date, SRP tailocins, including P2D1, have primarily been examined in the context of intraspecific competition, while their capacity to target nonkin bacterial taxa has not yet been systematically explored.
Here, we hypothesize that P2D1 tailocins can target phylogenetically distant environmental bacteria outside SRP, extending their ecological function beyond intraspecies competition. To demonstrate this, we independently screened for P2D1 susceptibility in a pool of environmental isolates and constructed deletion mutants of D. dadantii 3937 that lacked genes encoding core structural components of the P2D1 tailocin to test their interaction with these isolates. With this approach, we assessed for the first time the potential widespread ecological role of tailocins, specifically P2D1 produced by Dickeya spp. strains.
Materials and Methods
Bacterial strains and culture conditions
All strains used in this study are listed in Table 1. Unless otherwise specified, routine cultivation of strains was carried out at 28°C in Trypticase Soy Broth (TSB; Oxoid) with agitation at 120 rpm, or on Trypticase Soy Agar plates (TSA; Oxoid). Escherichia coli strains were cultured on Luria-Bertani (LB) agar supplemented with diaminopimelic acid (DAP) for the MFDpir strain. Chloramphenicol or ampicillin was added as required to maintain plasmids (Table S1).
Isolation of environmental bacterial strains
To obtain environmental isolates for subsequent testing for tailocin sensitivity, soil and rhizosphere samples were collected from three locations in Poland: tulip flower bed soil in Białogóra (54.81533° N, 17.96580° E), raspberry bush rhizosphere in Gdynia (54.50774° N, 18.54757° E), and cornfield soil in Częstochowa (50.95110° N, 19.22670° E). Samples used for microbial isolation were collected from environments with a potential presence of SRP bacteria [17]. Two grams of each sample were suspended in 4 ml of one-fourth-strength Ringer’s buffer (BioMaxima), shaken at 140 rpm for 30 min at room temperature, diluted 100× in the same buffer, and plated on 10% TSB agar. After incubation for 48 h at 28°C, morphologically distinct colonies were collected, and the resulting 480 bacterial isolates were subcultured onto TSB agar plates under the same conditions to obtain pure cultures [19].
Purification of P2D1 tailocins
P2D1 tailocin particles from D. dadantii 3937 were purified from mitomycin C–treated cultures using a previously described protocol [15]. The purified preparations were stored at 4°C until use.
Modified spot test for high-throughput P2D1 tailocin susceptibility screening
A fast and robust screening method involving 48-well plates (Greiner) was designed to test the new bacterial isolates for sensitivity to P2D1 tailocins. Each well was first filled with 20 μl of TSB medium. Next, a single bacterial colony of each tested strain was picked from a solid medium using a wooden toothpick and suspended evenly in the TSB droplet within the well. After inoculating all wells, 500 μl of molten soft-top agar [containing 30 g TSB and 7 g bacteriological agar (Oxoid) per 1 l], cooled to ~ 45°C, was added to each well. To facilitate immediate mixing of the inoculum with the freshly added medium, the multi-well plate was continuously agitated on a mini orbital shaker (Mini-Shaker, Biosan) (150 rpm) throughout the addition of the soft top agar. Once the agar had solidified, 2 μl of the purified P2D1 tailocins was spotted at the center of each well. Following 24 h of incubation at 28°C, the wells were inspected for zones of growth inhibition (clearance) indicative of tailocin activity. Wells inoculated with a P2D1-susceptible strain IFB 0117 [14] served as the positive control, while the negative control comprised the resistant P2D1 producer strain D. dadantii 3937. Isolates identified as sensitive during the screening were further tested using a classical spot assay [7].
Genomic sequencing
Whole-genome sequencing of P2D1-susceptible isolates was performed on a MinION platform (Oxford Nanopore Technologies) using the wf-bacterial-genomes Nextflow workflow (v1.4.1). Genomic DNA was extracted with the Wizard Genomic DNA Purification Kit (Promega). Sequencing libraries were prepared according to the Oxford Nanopore ligation sequencing protocol and loaded onto R9.4.1 flow cells. Basecalling was performed with Guppy (v6.x, Oxford Nanopore). Reads were assembled de novo using Flye (v2.9.5) and polished with Medaka (v2.0.0). Assemblies were annotated with the NCBI Prokaryotic Genome Annotation Pipeline (PGAP, February 2025 release).
Bioinformatic analyses
Taxonomic identification
Genome-based taxonomic identification and phylogenetic placement of the P2D1-sensitive isolates were performed using the JSpeciesWS web server [20] with FASTA genome sequences as input. First, the most closely related type strains in the GenomeDB database were identified for each isolate based on the highest Z-score correlation coefficient in pairwise tetra-nucleotide correlation (Tetra) analysis. Next, five type strains showing the highest Z-scores in Tetra-nucleotide analysis were subjected to pairwise Average Nucleotide Identity (ANI) calculations against the genomes of the strains under investigation. Both BLAST-based (ANIb) and MUMmer-based (ANIm) algorithms were applied. Additionally, the phylogenetic placement of the strains within the target genus was assessed based on 16S ribosomal RNA (rRNA) gene sequences (details are provided in Supplementary Dataset S1).
Dendrograms
To investigate the phylogenetic relationships between bacterial isolates, dendrograms were generated based on two independent datasets: genomic similarity and phenotypic profiles. Pairwise ANIm values (expressed as percent identity) were used to quantify genomic relatedness, and hierarchical clustering (agglomerative approach) was applied to group the strains accordingly. The same clustering method was used to analyze phenotypic data obtained from BIOLOG plate-based assays (described below), where positive and negative reactions were encoded as binary values (1 or 0). All clustering and dendrogram visualizations were performed using scikit-learn, SciPy, pandas, matplotlib, and numpy libraries [21, 22]. Python scripts supporting the analysis were developed with assistance from ChatGPT-4o (OpenAI) (Supplementary Script S1).
Comparative genomic analysis
To assess the genetic similarity among the nine P2D1-susceptible strains, we compared their genomes with respect to shared, accessory, and unique genes. The analysis was performed using the Pan-genome Explorer platform (https://panexplorer.southgreen.fr/cgi-bin/home.cgi) [23]. To assess the distribution of core and accessory genes, the PanACoTA pipeline was employed. Gene clusters predicted to be present or absent in individual environmental genomes were extracted and visualized using Venn diagrams generated in PAleontological STatistics (PAST) software [24].
Strain profiling with BIOLOG phenotypic microarrays
The ability of bacterial strains to utilize different carbon sources and their tolerance to various chemical stressors were assessed using the GEN III MicroPlate™ (94 phenotypic traits, including carbon utilization, chemical sensitivity, and physiological properties) and EcoPlate™ (31 carbon-source utilization traits) (Biolog) [25]. Plates were inoculated according to the manufacturer’s protocol and incubated at 28°C. At 24 and 48 h postinoculation, absorbance in each well was measured at 595 nm in the Epoch 2 microplate reader (BioTek). Results were normalized to the negative control and averaged across three biological replicates. A positive result was defined as an absorbance value at least twice that of the negative control.
Assessment of phenotypic traits in P2D1-sensitive bacterial isolates
Bacterial isolates susceptible to P2D1 were investigated for selected traits on agar-solidified media plates: colony morphology was assessed on TSA and M9 0.4% glucose (MP Biomedicals), pectinolytic activity was evaluated on Crystal Violet Pectate (CVP) [26], siderophore production was assessed using Chrome Azurol S (CAS) agar [27, 28], and pyoverdine production was evaluated in King’s B [29].
Microscopic imaging
The morphology of tailocin particles purified from the cultures of the wild-type D. dadantii 3937 and its mutants was investigated by transmission electron microscopy (TEM). TEM imaging was performed as described earlier [15] using the Tecnai Spirit BioTWIN microscope (FEI).
Construction of P2D1-deficient mutants
P2D1 production in D. dadantii 3937 was abolished by generating in-frame deletion mutants in the loci Dda3937_03810 (alternative locus designation DDA3937_RS12110) and Dda3937_03811 (DDA3937_RS12115). The two genes encode the tail sheath protein (ADM98779.1) and tail tube protein (ADM98780.1) of the tailocin, respectively [15]. Additionally, a double mutant deprived of both genes was constructed. The deletions were performed using the pRE112 suicide plasmid (Cm^R^) carrying the sacB gene to enable counter-selection [30] (Supplementary Table S1). Procedures were analogous to those described earlier for D. solani [30]. Two polymerase chain reaction (PCR) fragments corresponding to the upstream and downstream 0.5 kbp DNA of the gene(s) to be deleted in D. dadantii 3937 were amplified using the Primestar master mix (Takara) and cloned into SacI/KpnI digested pRE112 using the T5 exonuclease DNA assembly (TEDA) method [31] (list of oligonucleotides available in Supplementary Table S2). Chemical ultracompetent DH5α λpir cells were prepared with the Mix & Go! E. coli Transformation Kit using standard procedures (Zymo Research). Transformants were selected onto an LB plate supplemented with chloramphenicol (20 μg ml^−1^) and screened for the presence of the target construct by colony PCR with primers L762/L763. Constructs were extracted, confirmed by restriction map and Sanger sequencing, and then transferred into the competent E. coli strain MFDpir [18] prepared with the TSS method [31]. Escherichia coli MFDpir produces the RP4 conjugation machinery, which allows the transfer of the suicide plasmid into D. dadantii 3937 by conjugation. For conjugation, colonies of D. dadantii 3937 and MFDpir were mixed in the same proportion in 500 μl LB and centrifuged for 2 min at 8000 rpm. The pellet was resuspended in 90 μl LB with 5 μl diaminopimelic acid at 57 mg ml^−1^ and deposited onto an LB agar plate. After an overnight incubation at 30°C, the bacteria were resuspended in 1 ml LB, diluted in a 10-fold series from 10^−1^ to 10^−4^, and spread onto LB agar supplemented with chloramphenicol at 4 μg ml^−1^ to select the first event of recombination. Transconjugants re-isolated on this medium were then spread onto LB agar without NaCl, supplemented with 5% sucrose, and incubated at 20°C for 2–3 days to allow for the second recombination event. Sucrose-resistant colonies were then patched on LB-Cm plates to check for plasmid loss and streaked onto LB agar plates. The successful in-frame deletions were verified by colony PCR on purified colonies.
Engineering a complementation construct for the P2D1 mutation
Previous RNAseq experiments [32] indicate that transcription of the Dda3937_03810 gene initiates at position 2 751 779, which is 16 bases upstream of its start codon. Furthermore, a strong transcription termination signal is located at position 2 753 545—39 bases downstream of the Dda3937_03811 gene. These findings suggest that Dda3937_03810 and Dda3937_03811 function together as an operon. Based on these data, we constructed a complementation plasmid designed to co-express both Dda3937_03810 and Dda3937_03811. We cloned these genes, along with the 200 bp region upstream of Dda3937_03810, into the low-copy mobilizable plasmid pEGL332. This arrangement ensures that the native promoter for Dda3937_03810 is aligned in the same orientation as the plac promoter within pEGL332. As a result, in the complementation plasmid, Dda3937_03810 and Dda3937_03811 are transcribed under the control of both their native promoter and the plac promoter, which increases the likelihood of transcription of both genes. The PCR fragment containing Dda3937_03810 and Dda3937_03811 were amplified with oligonucleotide pairs L1802/L1803 and then cloned into HindIII-linearized pEGL332 by TEDA [31]. The hybrid plasmid, designated as pSJG, was verified by restriction mapping and DNA sequencing. Then, it was transferred to D. dadantii 3937 strains by biparental mating using MFDpir cell as the donor strain.
Phenotypic comparison between wild-type D. dadantii 3937 and mutant strains
Phenotypes of the D. dadantii 3937 tailocin-deficient mutants, with and without complementation (Table 1), were compared to the wild-type strain. The comparisons included tailocin production, pathogenicity on potato tubers, and metabolic profiles assessed using GEN III MicroPlates and EcoPlates (Biolog), as described above. In addition, growth rates of the strains were evaluated at 28°C in TSB and in M9 medium supplemented with 0.4% glucose.
Antibiosis assay
The inhibitory activity of D. dadantii 3937 against environmental Pseudomonas spp. strains and vice versa was evaluated in an in vitro antibiosis assay on TSA plates, following the protocol [33]. After overnight incubation at 28°C, cultures were examined for the presence of inhibition zones. The assay was performed twice, with three technical replicates each time.
Virulence assays on potato tubers
Whole-tuber injection assay
To prepare the bacterial inoculum, cells from an overnight culture in TSB were harvested by centrifugation (4200 RCF, 5 min) and resuspended in PBS buffer. The turbidity of the suspension was adjusted to 0.06 McF (ca. 2 × 10^6^ CFU ml^−1^). To prepare plant material, tubers cv. Gala were surface sterilized by immersion for 20 min in 5% commercial bleach (ACE, Procter and Gamble), followed by a double rinse in distilled water and air drying under laminar flow. Each tuber was inoculated by inserting a pipette tip containing 50 μl of the test suspension into the tuber (up to the level of liquid within the tip) [34]. Tubers inoculated with PBS buffer alone were used as a negative control. Ten tubers per treatment were used to assess the potential virulence of P2D1-sensitive environmental isolates and to compare the D. dadantii 3937 wild-type strain with its mutants. Inoculated tubers were placed in humid boxes (85%–90% relative humidity). Samples were incubated at 28°C to enable the development of soft rot symptoms. After 72 h, the tubers were cut at the inoculation site, and the tissue macerated by bacteria, if present, was spooned out and weighed. The experiment was conducted twice under the same conditions.
Potato tuber slices assay
Virulence and biocontrol assay on potato tuber slices was performed according to a modified protocol from Krzyzanowska et al. [35]. Surface-sterilized potato tubers cv. Gala were sliced, and three wells (5 mm in diameter) were created in each slice. The slices were then placed in large glass Petri dishes (18 cm in diameter) lined with Whatman 3 filter paper disks, cut to fit, and moistened with 5 ml of sterile distilled water. To prepare bacterial inocula, cells were collected from overnight TSB cultures (4200 RCF for 5 min) and resuspended in 0.85% NaCl. Biocontrol activity was assessed by mixing equal volumes of pathogenic strain suspensions (adjusted to 0.03 McFarland, ≈10^6^ CFU ml^−1^) and candidate biocontrol strains (3 McFarland, ≈10^8^ CFU ml^−1^). For pathogenicity assessment and single-strain controls, treatments consisted of a single strain mixed 1:1 with sterile 0.85% NaCl. Each well was filled with 30 μl of the corresponding mixture, with nine wells inoculated per treatment using three slices from different tubers. The slices were incubated in a humid chamber at 28°C for 48 h, after which the diameter of tissue maceration around each well was measured. The experiment was performed twice.
Results
P2D1 tailocin-sensitive isolates were detected across all sampled locations
Out of 480 environmental isolates tested, originating from three locations, 9 were susceptible to P2D1: 4 from raspberry bush rhizosphere soil (MC6, MD6, ME6, MB7), 2 from tulip flower bed soil (Tul1A2, Tul2A7), and 3 from cornfield soil (G3-3, G3-19, G3-39) (Fig. 1A).
Genomic and phenotypic characterization of P2D1 tailocin-susceptible Pseudomonas isolates. (A) Summary of isolation sources with the number of isolates obtained from each environment and the corresponding colour coding: red—cornfield soil; yellow—tulip flowed bed soil; blue—raspberry rhizosphere. (B) Phylogenetic tree of Pseudomonas-type strains based on 16S rRNA gene sequences. The tree was inferred using the maximum likelihood method with adaptive bootstrap support. For clarity, some branches are collapsed, while those containing the strains of interest are shown in full detail. The complete tree is available in Supplementary Dataset S1. (C) Heatmap of pairwise ANIm values with hierarchical clustering. Higher values (darker shading) indicate greater genomic similarity, and the dendrogram illustrates the relationships among isolates. (D) Distribution of core, accessory, and strain-specific (unique) genes in the pangenome. (E) Venn diagrams showing: the shared gene clusters between isolates from distinct environmental origins; gene cluster overlap between strains from the same environment; and a comparison of gene content between a rhizosphere isolate from raspberry and isolates from tulip. (F) Phenotype-based dendrogram derived from BIOLOG metabolic profiles (125 individual assays), illustrating phenotypic diversity among susceptible strains. For clarity, only traits showing variation across Pseudomonas spp. strains are displayed in the heatmap. D. dadantii 3937 was used as an outgroup. Complete results are provided in Supplementary Dataset S3.
P2D1-sensitive isolates are members of Pseudomonas spp.
Analysis of 16S rRNA gene sequences classified all nine P2D1-sensitive isolates within the genus Pseudomonas spp. (Fig. 1B; Supplementary Dataset S1). Whole-genome sequencing enabled higher-resolution taxonomic assignment based on ANI, the accepted standard for species delineation (species-level cutoff: ~95% to 96%).
Three of the four raspberry rhizosphere isolates (MD6, ME6, MB7) were assigned to P. germanica, while the two tulip flower bed isolates (Tul1A2, Tul2A7) aligned with P. tensinigenes (ANI >95%) (Table 2). Strain MC6 also clustered closest to the P. tensinigenes–type strain, but its ANIm (93%) and ANIb (92%) values fell below the species threshold. Cornfield soil isolates (G3-3, G3-19, G3-39) were most closely related to P. parakoreensis yet similarly failed species assignment (ANIb 92%, ANIm 93%). Thus, while several isolates could be confidently assigned to known Pseudomonas species, others (MC6, G3-3, G3-19, G3-39) likely represent previously undescribed taxa. Several isolates: G3-3 and G3-39 from cornfield soil; MD6, ME6, and MB7 from raspberry; and Tul1A2 and Tul2A7 from tulip—exhibited ANIb values exceeding 99% with >99% alignment (Supplementary Dataset S2). This suggests that, although independently recovered from environmental samples, isolates within each source are in most cases clonal or near-clonal, with the notable exceptions of strains MC6 and G3-19.
Hierarchical clustering of ANIm values revealed three robust clades (Fig. 1C). Generally, isolates from a given sampling location clustered together, demonstrating phylogenetic consistency within habitats. An exception was strain MC6, which, although isolated from raspberry, clustered with the two tulip flower bed isolates. Despite this close clustering, ANIm values below 90% indicated that MC6 was conspecific with neither its environmental group nor the tulip isolates.
Within each environmental clade, the remaining strains exhibited ANI values >95%, consistent with species-level similarity (Fig. 1C; Supplementary Dataset S2).
P2D1-susceptible Pseudomonas spp. isolates form distinct groups based on genomic and phenotypic traits
Genome sizes of the analyzed Pseudomonas isolates ranged from 6.12 to 6.63 Mbp (Table 2). The collective gene pool of the nine strains comprised 8121 genes, including 4181 (51.4%) core genes shared by all strains and 3062 (37.7%) accessory genes present in at least two genomes (Fig. 1D). Strain-specific genes (10.9% of the total) were largely contributed by MC6 (493 genes) and G3-19 (369). In contrast, the tulip flower bed isolates carried zero, and the remaining five isolates harbored only 2–12 each (Fig. 1D). The gene count for G3-19 may be less reliable than for the other strains, as its genome is represented by contigs rather than a closed assembly.
Genomic diversity varied across clades: the tulip flower bed isolates were nearly identical, with only three unique genes in total, whereas the raspberry rhizosphere and cornfield soil clades displayed greater heterogeneity, each comprising two or more distinct P2D1-susceptible Pseudomonas strains (Fig. 1E). Out of 125 traits assayed across both BIOLOG plates, 21 showed variation among the isolates. Overall, strain-relatedness inferred from biochemical profiles was consistent with that inferred from genomic sequence analysis (Fig. 1F).
In plate-based growth assays, all nine isolates grew on TSA and on M9 with glucose as the sole carbon source. Colonies appeared glossy and mucoid, with abundant exopolysaccharide production most pronounced on TSA—a trait not universally observed among Pseudomonas spp. (Fig. S1A and B). All strains fluoresced on King’s B medium (Fig. S1C), consistent with pyoverdine production typical of fluorescent Pseudomonas. Only the three cornfield soil isolates (G3-3, G3-19, G3-39) grew and produced siderophores on CAS medium after 96 h, whereas the remaining six isolates showed no growth (Fig. S1D). No pectinolytic activity was detected on CVP medium at 24 h. Still, cavities appeared after 96 h in the tulip isolates Tul1A2 and Tul2A7 and in the raspberry isolate MC6—the three strains that also cluster together in both genomic and biochemical analyses (Fig. S1E and F). These findings indicate genomic and phenotypic heterogeneity among the P2D1-susceptible Pseudomonas spp. isolates.
D. dadantii 3937 tailocins attach to Pseudomonas cells, leading to cell puncture
Using transmission electron microscopy (TEM), we observed that P2D1 tailocins from D. dadantii 3937 attach to the surfaces of susceptible Pseudomonas cells, similarly to their binding on the positive control strain D. zeae NCPPB 3532 (Fig. 2A–F). These results provide direct ultrastructural evidence that D. dadantii tailocins physically bind to and interact with environmental isolates not related to SRP.
P2D1 tailocins of D. dadantii 3937 attached to the cells of susceptible environmental Pseudomonas spp. panels: (A) Tul1A2; (B) G3-39; (C) ME6; (D, E) MD6; (F) D. zeae NCPPB3532 (positive control). Scale bar: 100 nm.
Tailocin-deficient D. dadantii mutants fail to kill environmental Pseudomonas spp.
To confirm that the inhibition of environmental strains by D. dadantii 3937 was mediated solely by tailocins, we constructed single (∆3810, ∆3811) and double (∆3810-11) mutants of D. dadantii, lacking genes encoding the sheath, the tube, or both genes, respectively (Table 1). None of these mutants produced complete (functional) tailocin particles (Fig. S2) or inhibited growth of the tailocin-susceptible SRP strain in soft-top agar overlay assays (Fig. 3). Aside from the absence of tailocin production, the mutants were indistinguishable from the wild type in growth rate (Fig. S3), virulence on potato tubers (Fig. S4, Fig. 4A), and biochemical profiles (Fig. S5). The double mutant (∆3810-11) was selected for subsequent experiments.
Activity of P2D1 tailocins isolated from D. dadantii 3937 WT, its P2D1-deficient mutant Δ3810-11, and a mutant with complementation plasmid pSJG. Tailocins were isolated from mitomycin C–induced cultures of the three tested strains and evaluated for activity against susceptible Pseudomonas isolates, as well as a control susceptible strain, D. zeae NCPPB 3532. The presence of a clearance zone indicates the presence of active tailocins in the tested preparation.
Effect of Pseudomonas isolates on potato tuber tissue in a slice assay. (A) shows the ability of pseudomonas isolates, D. dadantii 3937, and the 3937 tailocin mutant (Δ3810-11) to cause disease symptoms on tuber tissue when inoculated alone (pathogenicity). (B) shows the potential of the pseudomonas isolates to protect potato tissue from maceration when co-inoculated with the known pectinolytic pathogen D. dadantii 3937 (biocontrol activity). Median diameter of rotten tissue (mm; ± half interquartile range) is provided in the tables for all tested combinations. Alongside the tabulated results, the figure shows representative images depicting the characteristic appearance of healthy and diseased samples.
Tailocin preparations from induced ∆3810-11 cultures failed to inhibit any of the nine Pseudomonas isolates, in contrast to the wild type; however, complementation with plasmid pSJG (Table S2), which carries both disrupted genes under the operon’s native promoter, restored tailocin production and activity (Fig. 3, Fig. S2). Transformation with the complementation plasmid pSJG, most likely due to metabolic cost, resulted in a some unsignificant reduced growth rate of the transformed strains in M9 medium supplemented with 0.4% glucose, both in the mutant and in the wild-type host (Fig. S3).
Additionally, direct antibiosis was assessed using a plate assay to evaluate the production of antimicrobial metabolites. For both the wild type and ∆3810-11, only contact inhibition of Pseudomonas spp. isolates at colony borders was observed (Fig. S6A). These results indicate that, under the tested conditions, diffusible secondary metabolites of Dickeya do not contribute to antagonism against environmental Pseudomonas, supporting the conclusion that tailocins are the primary inhibitory factor. In a reciprocal setup, no inhibition of Dickeya strains by the tested Pseudomonas spp. was observed (Fig. S6B).
Pseudomonas strains sensitive to tailocin P2D1 either cause potato decay or suppress D. dadantii in biocontrol assays
In the potato slice assay, three Pseudomonas spp. strains—Tul1A2, Tul2A7, and MC6—caused tuber tissue maceration (Fig. 4A). These included two isolates originating from tulip flower bed soil and one raspberry isolate (MC6), all most closely related to P. tensinigenes. In the potato injection assay, which provides more microaerophilic conditions, only MC6 displayed mild pathogenicity within the experimental timeframe (72 h postinoculation). In contrast, six isolates (MD6, ME6, MB7, G3-3, G3-19, and G3-39) not only lacked pathogenicity when inoculated alone (Fig. 4A) but also exhibited a biocontrol effect, significantly reducing soft rot symptoms caused by co-inoculated D. dadantii 3937 (Fig. 4B; with Pseudomonas isolates applied in excess, as typical for biocontrol assays). Together, these findings highlight the contrasting outcomes among P2D1-susceptible Pseudomonas isolates, ranging from independent pathogenicity to biocontrol of soft rot.
Discussion
This study is the first to demonstrate that P2D1 tailocins produced by the Dickeya spp. can kill soil-associated Pseudomonas spp., which are phylogenetically distant from bacteria belonging to the Soft Rot Pectobacteriaceae (SRP) family. So far, tailocins have generally been described as possessing narrow host ranges, restricting their ecological role to competition with closely related strains [3, 36]. Only a few studies have demonstrated broader killing spectra, including Pseudomonas fluorescens tailocins suppressing Xanthomonas vesicatoria [8], a Burkholderia cenocepacia tailocin killing P. aeruginosa [7], and P. syringae targeting Erwinia amylovora, X. perforans, and the human pathogen Salmonella enterica [10]. Our present findings, derived from a study explicitly designed to search for such “off-target” effects, suggest that previously reported cases may not be rare exceptions but instead part of a broader and underexplored phenomenon with potentially significant ecological implications.
Among 480 environmental isolates tested, only nine (1.9%) were sensitive to P2D1 tailocins, yet these isolates were found across distinct environments and geographical locations. This so-called rare-but-widespread sensitivity pattern is consistent with results obtained in other studies, where tailocin-susceptible bacterial populations were typically low in frequency but widely distributed. For example, Yao and coworkers demonstrated that BceTMilo was active against 76 Burkholderia isolates, including both clinical and environmental samples, indicating its efficacy across multiple geographic and environmental contexts [7]. In other studies, maltocin P28 was active against 38 clinical and environmental Stenotrophomonas maltophilia strains [37], and tailocin from P. syringae USA011R targeted strains from distinct genera sourced from plant, clinical, and laboratory environments [10]. Likewise, R-type pyocins of P. aeruginosa targeted only a small fraction of clinical and environmental isolates; however, sensitive strains are found in different cystic fibrosis patients [38]. Furthermore, from an ecological perspective, finding susceptible isolates in geographically distant soils supports the idea that tailocins may act as selective forces in diverse plant microbiomes. Rhizosphere studies demonstrate that pyocins can significantly influence strain competition and community assembly, particularly under conditions of nutrient limitation [3].
Microscopic imaging revealed that P2D1 tailocins attach directly to the cell surface of susceptible Pseudomonas spp. isolates, resembling the interaction observed in the known susceptible strain D. zeae NCPPB 3532 [15]. This observation was supported by experiments with a P2D1-defective D. dadantii 3937 mutant, which failed to kill susceptible Pseudomonas spp. cells, thereby demonstrating that P2D1 tailocins alone are sufficient to mediate the observed killing. However, the specific surface determinants underlying P2D1 susceptibility in both Dickeya and Pseudomonas remain unidentified. Studies of Pseudomonas strains susceptible to tailocins produced by other Pseudomonas (pyocins) have shown that their binding specificity and target sensitivity are primarily governed by the structure, composition, and proper presentation of the lipopolysaccharide (LPS) O-antigen on the bacterial surface [39]. In the same model, conserved LPS chemotypes promote cross-species binding and susceptibility to tailocins among Pseudomonas strains [39]. In Dickeya spp. and the related Pectobacterium spp., LPS has been implicated in sensitivity to certain, but not all, bacteriophages [40, 41]. By contrast, the determinants of tailocin susceptibility in these bacteria remain uncharacterized. Identifying the surface features, particularly those shared between Dickeya spp. and Pseudomonas spp. that enable P2D1 cross-reactivity, may represent an important direction for future research, as described in other bacterial systems [42, 43].
In plant assays, P2D1-susceptible Pseudomonas spp. showed contrasting effects on plant tissue, both when inoculated alone and when co-inoculated with D. dadantii 3937. Three environmental isolates behaved as opportunistic soft-rot pathogens, whereas six others exhibited attenuation of D. dadantii-induced soft rot in potato tubers. These dynamics mirror the known dual roles of Pseudomonas spp.: some species (e.g. P. marginalis, P. palleroniana) cause soft rot [44, 45], whereas others serve as biocontrol and plant-beneficial agents [46]. Importantly, Pseudomonas spp. are known to co-occur with Dickeya spp. and Pectobacterium spp. in rotting potato tissues as part of a polymicrobial “spoilage microbiota” [47]. Several pectinolytic P. fluorescens strains have been identified alongside Pectobacterium spp. in diseased potato tubers in Kenya, confirming that both strains can coexist in rotten potato tissue [48]. At the same time, other Pseudomonas spp. are known biocontrol organisms: P. chlororaphis suppresses Dickeya spp. virulence by quenching quorum-sensing signals [49], while P. fluorescens eliminates competing Pectobacterium through a Type VI-secreted amidase, thereby protecting potato tubers [50]. Consistently, co-inoculation of antagonistic P. fluorescens, P. putida, or P. donghuensis strains has been shown to significantly reduce potato blackleg/soft-rot severity caused by SRP pathogens [19, 33, 35, 51].
It is therefore evident that P2D1-susceptible Pseudomonas spp. isolates can compete with SRP, including D. dadantii, on different planes—either as alternative pathogens of potato tissue or as antagonists mitigating soft rot. Our results, therefore, indicate that by targeting Pseudomonas with divergent ecological functions, P2D1 tailocins could function as potent modulators of both microbial competition and plant health.
Taken together, our findings indicate that tailocins produced by plant-associated bacteria can act against ecologically co-occurring competitors across genera, rather than being restricted to genetically closely related strains. Clusters encoding P2D1-like tailocins are widespread across Dickeya spp., suggesting positive selection for this trait [14]. Such an antagonistic capacity may be particularly advantageous for soft-rot pathogens, which cycle between insect, aquatic, soil, and plant niches and repeatedly encounter diverse resident microbiota [16, 17, 52]. Likewise, tailocin production is inherently costly, as it requires cell lysis and can leave producing populations vulnerable to competition by insensitive community members in mixed habitats [53]. Consequently, the net benefit of tailocin deployment likely depends on spatial structure, timing, and the fraction of cells committing to production [54]. At the same time, the precise contribution of P2D1 to Dickeya success during niche invasion remains unresolved. Future in vivo studies will be crucial to determining the ecological relevance of tailocins and their role in shaping microbial community structure.
Supplementary Material
SupplementaryDataset1_ycag012
SupplementaryDataset2_ycag012
SupplementaryDataset3_ycag012
SupplementaryScript1_ycag012
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Virgo M, Mostowy S, Ho BT. Emerging models to study competitive interactions within bacterial communities. Trends Microbiol 2025;33:688–700. 10.1016/j.tim.2024.12.00939799088 · doi ↗ · pubmed ↗
- 2Foster KR, Bell T. Competition, not cooperation, dominates interactions among culturable microbial species. Curr Biol 2012;22:1845–50. 10.1016/j.cub.2012.08.00522959348 · doi ↗ · pubmed ↗
- 3Backman T, Burbano HA, Karasov TL. Tradeoffs and constraints on the evolution of tailocins. Trends Microbiol 2024;32:1084–95. 10.1016/j.tim.2024.04.00139504934 PMC 12090994 · doi ↗ · pubmed ↗
- 4Ghequire MGK, De Mot R. The tailocin tale: peeling off phage tails. Trends Microbiol 2015;23:587–90. 10.1016/j.tim.2015.07.011 · doi ↗
- 5Taylor NMI, van Raaij MJ, Leiman PG. Contractile injection systems of bacteriophages and related systems. Mol Microbiol 2018;108:6–15. 10.1111/mmi.1392129405518 · doi ↗ · pubmed ↗
- 6Patz S, Becker Y, Richert-Poggeler KR et al. Phage tail-like particles are versatile bacterial nanomachines - a mini-review. J Adv Res 2019;19:75–84. 10.1016/j.jare.2019.04.00331341672 PMC 6629978 · doi ↗ · pubmed ↗
- 7Yao GW, Duarte I, Le TT et al. A broad-host-range tailocin from Burkholderia cenocepacia. Appl Environ Microbiol 2017;83:83. 10.1128/AEM.03414-16 · doi ↗
- 8Principe A, Fernandez M, Torasso M et al. Effectiveness of tailocins produced by Pseudomonas fluorescens sf 4c in controlling the bacterial-spot disease in tomatoes caused by Xanthomonas vesicatoria. Microbiol Res 2018;212-213:94–102. 10.1016/j.micres.2018.05.01029853172 · doi ↗ · pubmed ↗