Therapeutic potential of hookworm proteins in promoting regulatory immune responses to modulate Trypanosoma cruzi induced liver inflammation and oxidative stress
Maria Jose Villar, Cristina Poveda, Bin Zhan, Maya Gonsoulin, Kathryn M Jones, Villar Maria Jose, Kathryn M. Jones

TL;DR
This study explores how proteins from hookworms may help reduce liver inflammation caused by a parasitic infection, without worsening the infection itself.
Contribution
The study demonstrates the novel use of hookworm-derived proteins to modulate immune responses in T. cruzi-induced liver inflammation.
Findings
AIP-1 and AIP-2 increased IFN-γ and IL-10 while reducing Nfκ-B and Stat-1 in liver tissues.
AIP-1 uniquely upregulated Mmp9 and Btg2 gene expression.
Treatment increased specific immune cell populations but did not affect parasite load or fibrosis.
Abstract
Chronic Trypanosoma cruzi infection causes significant liver pathology, and current antiparasitic treatments often worsen hepatic damage. Hookworm-derived proteins have shown immunomodulatory effects in inflammatory diseases, including T. cruzi-induced myocarditis. This study evaluates recombinant hookworm proteins AIP-1 and AIP-2 for treating liver inflammation in a murine model of chronic Chagas disease (CD). Female BALB/c mice infected with T. cruzi were treated with AIP-1 or AIP-2 (1 mg/kg) for seven days. Controls were untreated or received aspirin (25 mg/kg) for 14 days. Liver tissues were analyzed for parasite burden (quantitative polymerase chain reaction - qPCR), histopathology (H&E, Picrosirius Red), and cytokines (multiplex assay). Splenocytes were assessed by flow cytometry, and serum was tested for liver enzyme levels. AIP-1 and AIP-2 increased hepatic interferon gamma…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
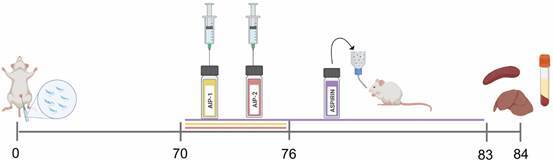 Figure 1
Figure 1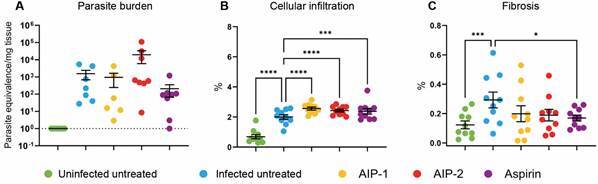 Figure 2
Figure 2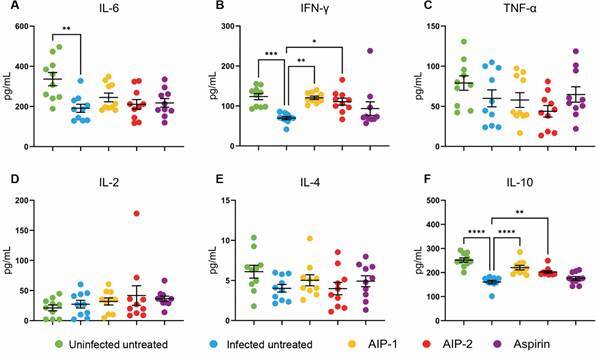 Figure 3
Figure 3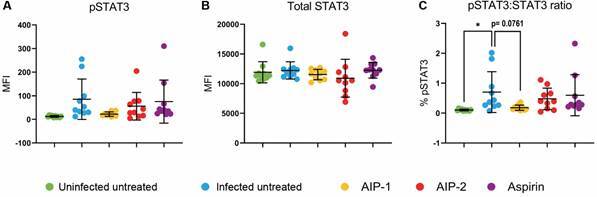 Figure 4
Figure 4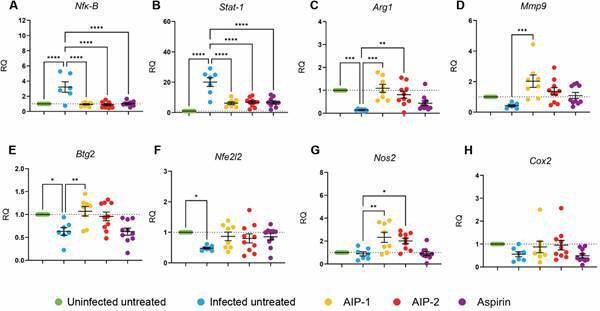 Figure 5
Figure 5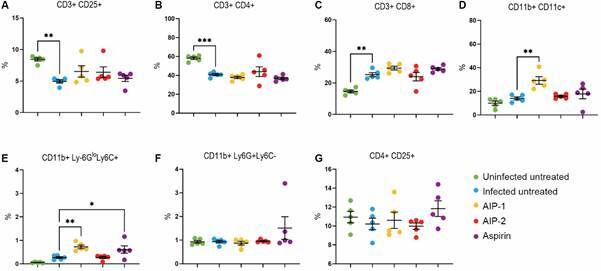 Figure 6
Figure 6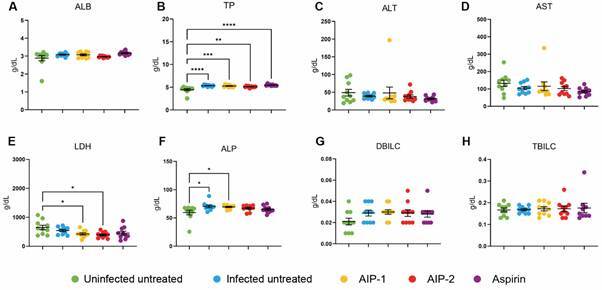 Figure 7
Figure 7- —NIH
- —the Comprehensive Cancer Center
- —CPRIT
- —NIH
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTrypanosoma species research and implications · Invertebrate Immune Response Mechanisms · Parasites and Host Interactions
Chagas disease (CD), caused by the protozoan parasite Trypanosoma cruzi, typically spreads through contact with feces from infected triatomine vectors, commonly referred as ‘kissing’ bugs. This neglected tropical disease is primarily prevalent in rural areas of the Southern United States, Mexico, Central, and South America, affecting approximately 6-7 million people globally.1 Rapid urbanization and migration from rural areas contribute to its spread beyond Latin America, resulting in cases emerging in regions such as Europe, Australia, and Japan.2 Recognizing the signs and symptoms of this infection is crucial, particularly since the acute stage often goes untreated, potentially leading to complications in the heart, digestive tract, brain, and other organs.3
Acute CD typically presents with asymptomatic or mild, non-specific symptoms after an incubation period of one to two weeks.4 As a result, it often progresses to the chronic stage, leading to severe complications due to tissue inflammation and degradation.5 T. cruzi infections evade innate immunity, in part by resisting phagocytosis. This process modulates nitric oxide (NO) levels and activates macrophages and cytokines.6 Such macrophage activation triggers a cascade of cytokines, including interleukin-2 (IL-2) and interferon-γ (IFN-γ), while regulatory cytokines such as IL-10 and IL-4 help mitigate potential harmful effects of an overactive immune response.6 In experimentally infected animals, myeloid derived suppressor cells (MDSC) accumulate in the heart and spleen, regulating inflammatory T cell proliferation and promoting regulatory T cells. This mechanism helps control tissue damage while facilitating parasite persistence.7 ^,^ 8
The only approved drugs for CD are benznidazole (BNZ) and nifurtimox, both of which have demonstrated a trypanocidal effect on the parasite. However, these antiparasitic treatments only reduce parasite load and may lead to toxic side effects, especially in chronic cases, when administered over an extended period.9 BNZ is extensively metabolized in the liver by the cytochrome P450 enzyme complex, often resulting in Grade 1 or Grade 2 toxicity during treatment. It is also associated with elevated serum enzymes, such as AST and ALT.10 ^,^ 11 Infection with T. cruzi can lead to severe cardiological and gastrointestinal complications and, in the presence of underlying conditions, infection can aggravate liver inflammation and accelerate hepatic injury.12 Importantly, both T. cruzi infection and BNZ have been shown to stimulate oxidative stress in the liver, leading to pathologic remodeling.13
The liver plays a pivotal role in inflammatory responses, notably through the acute-phase response and oxidative stress mechanisms. When inflammation occurs, the liver produces acute-phase proteins (APPs), which indicate systemic inflammation. This process is mainly controlled by IL-6, a cytokine released by hepatic macrophages and other cells. IL-6 prompts hepatocytes to generate APPs such as C-reactive protein and fibrinogen.14 The balance between positive APPs, which increase during inflammation, and negative APPs, which decrease, is essential for evaluating the body’s inflammatory response. Due to its role in metabolizing various substances, the liver is particularly vulnerable to reactive oxygen species (ROS), increasing the risk of oxidative stress.
The persisting ROS release from a chronic T. cruzi infection leads to mitochondrial dysfunction, inflammation, and heart tissue damage.15 Additionally, T. cruzi infection induced oxidative stress in the livers of infected mice.16 Therefore, this stress may contribute to liver damage and further inflammation.14 ^,^ 17 Drug-induced liver injury (DILI) arises when disruptions in drug metabolism trigger liver damage, often due to oxidative stress, inflammation, apoptosis, necrosis, or mitochondrial membrane breakdown, ultimately compromising organ function.18 As an alternative approach to reduce inflammation caused by infectious pathogens or autoimmune diseases, helminth-derived proteins have demonstrated significant anti-inflammatory property. Previous studies have shown that canine hookworm Ancylostoma caninum-secreted AIP-1 and AIP-2 conferred therapeutic effect on colitis 19 and cardiac inflammation caused by chronic infection of T. cruzi.20 In this study, we propose a novel treatment strategy utilizing these two hookworm-derived recombinant proteins (AIP-1 and AIP-2) provided immunomodulatory and antioxidant properties to treat T. cruzi infection caused liver inflammation and damage.20
MATERIALS AND METHODS
Ethics statement - Animal experiments were performed in full compliance with the Public Health Service Policy and the National Institutes of Health Guide for the Care and Use of Laboratory Animals, 8th edition, under a protocol approved by Baylor College of Medicine’s Institutional Animal Care and Use Committee, assurance number D16-00475.21
Expression and purification of recombinant protein - The recombinant proteins, AIP-1 and AIP-2, were expressed and purified as previously described.20 Briefly, DNDs encoding for Ac-TMP-1 (AIP-1) and Ac-TMP-2 (AIP-2) were cloned into Pichia pastoris expression vector pPICZαA. The recombinant plasmid DNAs including AIP-1 and AIP-2 were transformed into P. pastoris X-33, the recombinant proteins (AIP-1 and AIP-2) were expressed under induction of 1% methanol and purified with immobilized metal affinity chromatography for AIP-1 (with His-tag at C-terminus) and ion exchange (HiTrap QXL) for AIP-2.
Mice and parasites - Male ICR-SCID (ICRSC-M) and female BALB/c (BALB/cAnNTac), five-six weeks of age from Taconic (Taconic Biosciences, Inc) were allowed to acclimate for approximately one week prior to initiating studies. Mice were housed four per group in small microisolator cages and provided ad libitum food and water, under a 12 h light/ dark cycle. To expand blood form trypomastigotes (bft) for experimental infection, male ICR-SCID mice were infected with 5000 bft of the bioluminescent T. cruzi H1 strain (T. cruzi H1 K68) generated in our laboratory20 ^,^ 22 by intraperitoneal (IP) injection. Approximately 28 days after infection, ICR-SCID mice were humanely euthanized and bft were collected, washed with sterile medical grade saline, and used to infect BALB/c mice.
Experimental infection and treatments - Female BALB/c mice were infected with 5000 T. cruzi H1 K-68 bft by IP injection. At 70 days post-infection (dpi), animals were randomly assigned to treatment groups as outlined in Table.
TABLEStudy groupsGroupInfectionTreatmentDurationRoute of administration#1NoneNoneN/AN/A#2 Trypanosoma cruzi H1 K68NoneN/AN/A#3 Trypanosoma cruzi H1 K681 mg/kg AIP-1Once daily for seven daysIntraperitoneal injection#4 Trypanosoma cruzi H1 K681 mg/kg AIP-2Once daily for seven daysIntraperitoneal injection#5 Trypanosoma cruzi H1 K6825 mg/kg Aspirin14 days Ad libitum drinking waterN/A: not applicable.
As illustrated in Fig. 1, mice received daily IP injections of either AIP-1 or AIP-2 protein (1 mg/kg) for seven consecutive days. Control groups included uninfected mice, infected untreated mice, and infected mice administered aspirin (25 mg/kg) via drinking water ad libitum for 14 consecutive days. Aspirin served as a positive control, administered according to a published study,23 to provide a benchmark for anti-inflammatory activity in T. cruzi-infected mice. At 84 dpi, liver, spleen, and serum samples were collected for comprehensive immunological analysis. These assessments included quantification of parasite burden, evaluation of liver histopathology, cytokine profiling via Luminex multiplex assay, relative gene expression analysis, splenic immune cell profiling by flow cytometry, and examination of serum for clinical abnormalities.
Fig. 1:study timeline. At 84 days post-infection (dpi), liver, spleen, and serum samples were collected for analysis, including parasite quantification, histopathology, cytokine profiling, gene expression, flow cytometry, and serum enzyme levels. Image created with BioRender.com.
Liver collection - The liver was harvested using aseptic techniques in cold 1X phosphate-buffered saline (PBS) within a biosafety cabinet. The largest liver lobe was then divided into four sections for DNA, RNA, and protein analyses, snap-frozen, and stored at -80ºC until further use. The fourth liver section was designated for histopathological analysis, wherein it was stained with Hematoxylin and Eosin (H&E) and Picrosirius Red to assess inflammation and fibrosis, respectively.
DNA extraction and parasite burden - DNA was extracted from a frozen liver section using PDQeX Nucleic Acid Extractor and PreGem Universal kits (MicroGem International) according to the manufacturer’s protocol. Levels of T. cruzi in the liver were quantified using Taqman primers and probes for the T. cruzi satellite DNA and mouse GAPDH as previously described24 [Supplementary data (Table I)]. Samples were amplified using a QuantStudio 7 Pro Real Time thermocycler. The observed Ct mean for each sample was normalized to calculate the parasite burden per mg tissue using the equation derived from the standard curve.
Histopathology - Formalin-fixed, paraffin-embedded liver tissues were sectioned at 5 µm and serial sections spaced 10-30 µm apart were analyzed to represent approximately one-quarter of the organ as described previously.25 Sections were stained with H&E for inflammation and Picrosirius Red for fibrosis. For each section, four representative fields were captured at 10× magnification using an AmScope 40X-2000X microscope. Inflammation was quantified by counting lymphocyte nuclei in Fiji ImageJ, applying a particle size filter to exclude hepatocyte nuclei and debris, with counts from the four selected fields averaged per section. Fibrotic areas were measured as pink-stained regions, and all values were normalized to tissue area to enable accurate comparisons across sections and animals.
Preparation of liver lysate - A snap-frozen liver section was maintained on ice and transferred to a GentleMACS M-tube containing RIPA Lysis and Extraction Buffer (Thermo Fisher) to ensure protein integrity and minimize degradation during subsequent processing. Subsequently, the liver was homogenized using the gentleMACS™ Dissociator’s Protein saved program. The resulting lysate was centrifuged at 500 x g for 5 min, and the supernatant was carefully transferred to a low-temperature resistant Eppendorf tube. Total protein concentration was determined using Pierce™ BCA Protein Assay Kits (Thermo Fisher). Stock lysate was stored at -80ºC until use.
Luminex multiplex bead panel - The liver lysate was diluted to 5 µg/µL for the subsequent assessment of cytokines and proteins using Luminex assays. Two kits were used for these assays: the MILLIPLEX^®^ Phospho/Total STAT3 Magnetic Bead 2-Plex Kit (Millipore Sigma) and the Mouse Th17 Panel MAGNETIC with IL-6, IFN-γ, TNF-α, IL-2, IL-4, and IL-10 (Millipore Sigma). Protein and cytokine levels were then determined and normalized to the standards provided with the respective kits.
RNA extraction and reverse transcription polymerase chain reaction (RT-PCR) - RNA was extracted from a frozen liver section using the Qiagen RNeasy Plus Mini Kit, following the manufacturer’s protocol. The RNA concentration was determined using the Thermo Fisher NanoDrop One/OneC UV-Vis Spectrophotometer, and the final product was diluted to 10 ng/µL with RNase-free water for RT-PCR. An RT-PCR mix was prepared using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems™), which was then added to each diluted sample before using the Biorad T100 Thermal Cycler.
For RT-PCR, the thermal cycler was programmed to maintain 25ºC for 10 min, followed by 37ºC for 2 h, then 85ºC for 5 min, and finally held at 4ºC until the tubes were retrieved. cDNA was stored in -20ºC until use. The cDNA was diluted 1:10 to evaluate gene expression using quantitative PCR (QuantStudio 7 Pro). Gene expression analysis was performed using TaqMan primers targeting Nuclear factor kappa-light-chain-enhancer of activated B cells (Nfk-B), Signal transducer and activator of transcription 1 (Stat-1), Nitric oxide synthase 2 (Nos2), Cyclooxygenase-2 (Cox2), Arginase 1 (Arg1), Matrix metalloproteinase-9 (Mmp9), B-cell translocation gene (Btg2), and Nuclear factor erythroid 2-related factor 2 (Nfe2I2) [Supplementary data (Table II)]. Each sample was analyzed in duplicate. The relative expression level (RQ) of each target gene was determined using the 2-ΔΔCt method, where ΔCt is the difference between the Ct values of the target gene and Gapdh [Ct(target) - Ct(Gapdh)], and ΔΔCt is the difference between the ΔCt of the sample and the average ΔCt of the uninfected untreated control mice. The RQ was then calculated using the formula RQ = 2(-ΔΔCt).20
Single cell suspension of splenocytes - Spleens were first rinsed with sterile 1X PBS and transferred to a gentleMACS C Tube containing 3 mL of sterile PBS for homogenization using a gentleMACS Dissociator (Miltenyi Biotech). To lyse red blood cells, ACK lysis buffer (Lonza) was added to the spleen homogenates, and the solution was diluted 5-fold with RPMI medium supplemented with 10% fetal bovine serum (FBS), 1X penicillin-streptomycin (Pen-Strep), and L-glutamine (cRPMI medium). Splenocytes were then pelleted by centrifugation at 300×g for 5 min. The resulting pellet was resuspended in 5 mL of cRPMI medium and passed through a 40 μm filter (BD Biosciences) to remove debris. Cell viability was assessed using acridine orange-propidium iodide (AOPI) live/dead dye and a Cellometer Auto 2000 (Nexcelom Bioscience) automated cell counter. For each sample, 1 × 10^6^ viable splenocytes were plated in a 96-well non-tissue culture plate and cells were stained for immunophenotyping analysis.
Immunophenotyping panel - Immunophenotyping was performed by first staining 1 × 10⁶ splenocytes with ViaDye Red fixable viability dye for 30 min on ice. After viability assessment, cells were incubated with a cocktail of fluorochrome-conjugated antibodies targeting surface markers - CD3, CD4, CD8, CD25, CD11b, CD11c+, Ly6G and Ly6C - [Supplementary data (Table III)] for 30 min on ice in the dark. Then, it was fixed using BD Cytofix for 20 min. After fixation, cells were resuspended and analyzed using an Aurora spectral flow cytometer. Data analysis was performed using FlowJo software. The representative gating strategy is shown in Supplementary data (Figure).
Liver enzymes - Whole blood was collected in serum separator tubes and left to clot at room temperature (RT) for at least 30 min. The tubes were then centrifuged at 10,000xg for 5 min at RT to separate the serum. The serum was transferred to a cryovial and frozen at -80ºC. A 100 µL serum sample was sent to the Clinical Pathology Laboratory at Baylor College of Medicine for analysis. Liver enzymes, including albumin (ALB), total protein (TP), alanine transaminase (ALT), aspartate transaminase (AST), lactate dehydrogenase (LDH), alkaline phosphatase (ALP), direct bilirubin (DBILC), and total bilirubin (TBILC), were measured.
Statistical analysis - All data were analyzed and visualized using GraphPad Prism (Version 10.2.3). Statistical significance was determined by two-way analysis of variance (ANOVA) with multiple comparisons relative to the Infected Untreated control group (p < 0.05). In addition, liver enzyme levels from serum samples were also compared to the Uninfected Untreated control using the same analysis. Error bars represent the mean ± standard deviation (SD). Significance is indicated as follows: *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001.
RESULTS
Parasite load, hepatic cellular infiltration, and fibrotic changes - Our results confirm successful infection in the mice, as demonstrated by an increased parasite burden across all infected groups compared to the Uninfected Untreated group (Fig. 2A). Although no statistically significant differences in liver parasite burdens were observed, notable variations in cellular infiltrate and fibrosis were detected. As expected, infection led to a significant increase in hepatic cellular infiltrate compared to uninfected mice. Similar to our previous cardiac study,20 parasitemia levels and cardiac parasite burdens did not differ with treatment, indicating that the observed effects are independent of parasite load. Treatment with AIP-1, AIP-2, and aspirin each resulted in a further significant increase in cellular infiltrate compared to the Infected Untreated group (Fig. 2B).
Fig. 2:parasite burden and tissue pathology. Parasite burden in the liver was measured at the end of the study by quantitative polymerase chain reaction (qPCR). Hepatic cellular infiltration was evaluated using Hematosin & Eosin (H&E) staining, while fibrosis was assessed through Picrosirius Red staining. The resulting images were analyzed with ImageJ software. (A) Parasite burden; (B) Cellular infiltration; (C) Fibrosis. Data from individual mice are shown, n = 10. Error bars are defined by mean with standard deviation (SD). *p ≤ 0.05; ***p ≤ 0.001; ****p ≤ 0.0001.
Additionally, infection induced a significant increase in fibrosis relative to Uninfected Untreated mice; however, only aspirin treatment significantly reduced fibrosis when compared to Infected Untreated mice (Fig. 2C). These data suggest that the immunomodulatory effects of AIP-1 and AIP-2 in the liver are not mediated by reductions in parasite burden.
Interestingly, while fibrosis levels remained unchanged in the AIP-1- and AIP-2-treated groups compared to the Infected Untreated group, cellular infiltration followed a distinct pattern. These findings highlight the complex relationship between immune response and liver pathology in T. cruzi infection, underscoring the need for further investigation into how AIP-1 and AIP-2 influence these processes.
Treatments with AIP-1 and AIP-2 promote a balanced hepatic immune response - To assess the impact of treatments on inflammatory cytokines in the liver, Luminex assays were performed on liver protein extracts. Pro-inflammatory cytokines - including IL-6, IFN-γ, and TNF-α - were measured alongside anti-inflammatory markers such as IL-2, IL-4, and IL-10.
Our results show that, compared to Uninfected Untreated mice, infection alone led to a significant decrease in IL-6 (Fig. 3A), IFN-γ (Fig. 3B), and IL-10 (Fig. 3F), with no significant differences observed in TNF-α (Fig. 3C), IL-2 (Fig. 3D), or IL-4 (Fig. 3E). Interestingly, treatment with AIP-1 and AIP-2 resulted in a significant increase in IFN-γ (Fig. 3B) and IL-10 (Fig. 3F) compared to the Infected Untreated group. No significant changes were observed in IL-6, TNF-α, IL-2 or IL-4 levels following AIP-1 or AIP-2 treatment.
Fig. 3:cytokines from liver lysate. Pro- and anti-inflammatory cytokine levels were quantified using protein extracts from liver samples. (A) Interleukin (IL)-6 ; (B) Interferon (IFN)-γ; (C) Tumor necrosis factor (TNF)-α; (D) IL-2; (E) IL-4; (F) IL-10. Data from individual mice are shown, n = 10. Error bars are defined by mean with standard deviation (SD). *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001.
Together, these data suggest that AIP-1 and AIP-2 may promote a more balanced T_H_1/T_H_2 hepatic immune environment, characterized by elevated levels of both pro- and anti-inflammatory cytokines. In our previous study,20 we reported that serum cytokine levels differed from cardiac tissue cytokine levels, and here we similarly show that hepatic cytokine levels follow a distinct pattern from serum. This underscores the importance of examining local tissue immune responses to fully assess the impact of treatment.
Treatment with AIP-1 reduced levels of pSTAT3 - Our findings show no significant differences in pSTAT3 or total STAT3 levels among the treatment groups compared to the Infected Untreated group (Fig. 4A-B). However, T. cruzi infection alone significantly increases the pSTAT3:STAT3 ratio relative to Uninfected Untreated mice, while AIP-1 treatment trends toward reducing this ratio, with a p-value of 0.0761 (Fig. 4C).
Fig. 4:signal transducer and activator of transcription 3 (STAT3) levels from liver lysate. Total STAT3 and phosphorylated (p)-STAT3 levels were quantified by Luminex assays. (A) pSTAT3 levels; (B) Total STAT3 levels; (C) pSTAT3 ratio. Data from individual mice are shown, n = 10. Error bars are defined by mean with standard deviation (SD). *p ≤ 0.05.
Treatment with AIP-1 and AIP-2 modulate expression of inflammatory and oxidative stress genes - T. cruzi infection induces inflammation and oxidative stress in the liver.16 ^,^ 26 In our model, infected untreated mice showed increased expression of Nfκ-B (Fig. 5A) and Stat-1 (Fig. 5B) compared to Uninfected Untreated mice, while the expression of Arg1 (Fig. 5C), Mmp9 (Fig. 5D), Btg2 (Fig. 5E), and Nfe2l2 (Fig. 5F) was decreased. Treatment with AIP-1, AIP-2, and aspirin reduced the expression of Nfκ-B and Stat-1 relative to Infected Untreated mice, suggesting that these treatments downregulate inflammatory gene expression.
Fig. 5:relative quantification of gene expression levels in liver tissues. Gene expression was quantified using qPCR on cDNA synthesized from each liver sample. (A) Nuclear factor kappa-light-chain-enhancer of activated B cells (Nfk-B); (B) Signal transducer and activator of transcription 1 (Stat-1); (C) Arginase 1 (Arg1); (D) Matrix metalloproteinase-9 (Mmp9); (E) B-cell translocation gene 2 (Btg2); (F) Nuclear factor, erythroid 2-like 2 (Nfe2l2); (G) Nitric oxide synthase 2 (Nos2); (H) Cyclooxygenase-2 (Cox2). Data from individual mice are shown, n = 10. Error bars are defined by mean with standard deviation (SD). *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001.
Additionally, AIP-1 and AIP-2 significantly increased the expression of Arg1 (Fig. 5C) and Nos2 (Fig. 5G) compared to Infected Untreated mice. Co-expression of Arg1 and Nos2 is a hallmark of myeloid-derived suppressor cells (MDSCs), which play a role in nitric oxide-mediated suppression of immune cell proliferation and promote survival in experimental infection models.26 These findings suggest that AIP-1 and AIP-2 may enhance recruitment of MDSCs to the liver, thereby reducing inflammation.
Interestingly, only AIP-1 treatment increased Mmp9 expression compared to Infected Untreated mice (Fig. 5D). Mmp9 is involved in tissue remodeling and fibrosis during CD,27 but it has also been shown to release TNF-α from cell surfaces, indicating a dual role in promoting inflammation.28 Our observation of reduced Mmp9 expression in Infected Untreated mice compared to the Uninfected control suggests that T. cruzi may suppress Mmp9 as an immune evasion strategy, which is potentially reversed by AIP-1 treatment.
Finally, AIP-1 significantly increased Btg2 expression relative to Infected Untreated mice (Fig. 5E). Btg2 is known to trigger antioxidant responses, including activation of Nfe2l2, and is typically downregulated in oxidative stress and cancer models.29 The Nfe2l2 pathway is a key antioxidant defense mechanism in the liver.17 In this study, we show that T. cruzi infection significantly reduces the expression of Btg2 and Nfe2l2, while treatment with AIP-1 and AIP-2 trends toward restoring their expression (Fig. 5E-F), suggesting a potential role in enhancing protective antioxidant responses during infection.30 ^,^ 31
Alterations in splenic immune cell populations following T. cruzi infection and treatment with AIP-1 and AIP-2 - Splenocyte single-cell suspensions were stained with CD3, CD11b, CD11c, Ly6G, Ly6C, CD4, CD8, and CD25, and analyzed by flow cytometry. All infected mice exhibited a decrease in the CD3⁺CD25⁺ population compared to Uninfected Untreated control, with a significant reduction observed in the Infected Untreated group (Fig. 6A). While the CD4⁺ population was reduced across all infected groups, the CD8⁺ population increased in response to infection. Notably, mice treated with AIP-1, AIP-2, or aspirin exhibited a trend toward reduced frequencies of CD4⁺ T cells accompanied by a corresponding increase in CD8⁺ T cells (Fig. 6B-C), suggesting a potential shift in T cell dynamics.
Fig. 6:immunophenotyping of immune cell populations. Splenocytes were stained and acquired by flow cytometry. (A) Cluster of differentiation (CD)3+CD25+; (B) CD3+CD4+; (C) CD3+CD8+; (D) CD11b+CD11c+; (E) CD11b+Ly6G^lo^Ly6C+; (F) CD11b+Ly6G+Ly6C-; (G) CD4+CD25+. Data from individual mice are shown, n = 10. Error bars are defined by mean with standard deviation (SD). *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001.
We also analyzed monocyte populations and observed that treatment with AIP-1 and AIP-2 led to significant alterations in splenic immune cell composition. AIP-1 notably increased the frequency of CD11b⁺CD11c⁺ dendritic cells (DCs) and CD11b+Ly6G^lo^Ly6C+ MDSCs. Specifically, infected mice treated with AIP-1 displayed a significant rise in both DCs (Fig. 6D) and MDSCs (Fig. 6E) compared to Infected Untreated mice, suggesting that AIP-1 promotes the expansion of these regulatory immune cell populations.
In contrast, no significant differences were observed in the frequencies of CD11b⁺Ly6G⁺Ly6C⁻ neutrophils (Fig. 6F) or CD4⁺CD25⁺ regulatory T cells (Fig. 6G) across treatment groups relative to the Infected Untreated control.
Liver enzymes suggest a favorable response from the treatment groups - Plasma samples were collected and analyzed to assess liver function and identify potential abnormalities resulting from infection or treatment.
ALB and TP levels serve as indicators of liver synthetic function, with reduced ALB often linked to chronic liver disease or inflammation.32 ^,^ 33 Treatment with AIP-2 resulted in lower ALB levels compared to the Infected Untreated group; however, no significant differences were observed between the treatment groups and the Uninfected Untreated control, except for a notable increase in ALB levels in the Aspirin group (Fig. 7A). Similarly, all infected groups showed elevated TP levels relative to the Uninfected Untreated control, suggesting that T. cruzi infection may impair liver synthetic function (Fig. 7B).
Fig. 7:liver enzymes. Whole blood plasma was used to quantify liver function enzymes to evaluate for clinical implications of abnormalities in the liver. (A) albumin (ALB); (B) total protein (TP); (C) alanine transaminase (ALT); (D) aspartate aminotransferase (AST); (E) lactate dehydrogenase (LDH); (F) alkaline phosphatase (ALP); (G) direct bilirubin (DBILC); (H) total bilirubin (TBILC). Data from individual mice are shown, n = 10. Error bars are defined by mean with standard deviation (SD). *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, ****p ≤ 0.0001.
ALT and AST are key markers of hepatocellular damage and are commonly used to monitor treatment response, as elevations in these enzymes post-treatment can indicate hepatotoxicity in Chagasic patients.10 ^,^ 33 In this study, treatment with either AIP-1 or AIP-2 did not significantly affect ALT or AST levels when compared to either the Uninfected Untreated or Infected Untreated groups (Fig. 7C-D).
LDH, although a nonspecific marker, is often associated with hepatocellular damage.34 Interestingly, LDH levels were significantly reduced in the AIP-1 and AIP-2 treatment groups compared to the Uninfected Untreated control (Fig. 7E). Lastly, ALP and DBILC are typically associated with biliary obstruction. The observed increase in ALP levels in the AIP-1 group compared to the Uninfected Untreated control may reflect infection-related changes (Fig. 7F).
DISCUSSION
We previously demonstrated in our experimental mouse model of early chronic T. cruzi infection that the hookworm-derived proteins AIP-1 and AIP-2 significantly reduce cardiac cellular infiltrate and expression of inflammatory genes and cytokines.20 Building on these findings, this study explored the liver-specific immune response to AIP-1 and AIP-2 treatment, as the liver plays a critical role in the clearance of blood-stage T. cruzi parasites and the induction of protective immune responses during chronic infection.35 ^,^ 36 Given the significant inflammatory burden associated with T. cruzi-induced liver pathology, we aimed to determine whether AIP-1 and AIP-2 could mitigate inflammation while preserving hepatic function. Our findings suggest that these recombinant proteins function not merely as anti-inflammatory agents, but as immune modulators with potential antioxidant properties - capable of rebalancing immune responses, dampening pro-inflammatory signaling pathways, and promoting protective gene expression, all without causing hepatotoxicity or worsening fibrosis.
We observed increased cellularity in the liver (Fig. 2B), however levels of the proinflammatory cytokines IL-6 and TNF-α remained unchanged across infected treatment groups, suggesting that the increased immune cell presence was not inflammatory in nature. Instead, flow cytometry data revealed that AIP-1 treatment elevated the proportion of CD11b⁺CD11c⁺ dendritic cells and CD11b⁺Ly6G^lo^Ly6C⁺ MDSCs (Fig. 6D-E). Given the well-established roles of MDSCs in dampening immune overactivation and promoting tissue repair37 ^,^ 38 their expansion may explain the observed immune cell infiltration without an associated rise in proinflammatory cytokines. MDSCs are also known to inhibit T-cell proliferation and contribute to IL-10-mediated downregulation of hepatic stellate cell activation, potentially mitigating fibrosis.38 Although we observed increased MDSCs in the spleen following AIP-1 treatment, further investigation is needed to determine whether similar populations accumulate in the liver and contribute to the increased infiltrate observed histologically in Fig. 2B.
Consistent with previous studies, T. cruzi infection increased hepatic pSTAT3 levels, a key driver of inflammation and fibrosis.39 ^,^ 40 ^,^ 41 AIP-1 treatment was associated with a trending reduction in the pSTAT3 ratio (Fig. 4C), which may help explain the observed tendency toward reduced fibrosis in treated mice, despite the increase in immune cell infiltration. STAT3 inhibition, alongside modest increases in Mmp9 - a matrix metalloprotease involved in extracellular matrix remodeling - suggests a potential mechanism for preserving liver architecture without promoting fibrogenesis.42 ^,^ 43 Although STAT-3 is typically activated by IL-6, it can also be triggered by other cytokines, including IL-10.44 In our previous cardiac study,20 infection was associated with elevated serum IL-10, which may have contributed to the observed increase in hepatic pSTAT3, even though IL-10 levels within the liver were reduced by T. cruzi infection. Because both IL-6 and IL-10 can be produced by immune and non-immune cells in the liver, future studies will aim to characterize the liver immune cell composition to identify the primary cytokine-producing cells and determine how AIP-1 and AIP-2 affect both immune and non-immune cell populations.
Transcriptional profiling revealed further support for this immune-modulatory role. AIP-1 and AIP-2 treatments upregulated protective genes such as Btg2, Nos2, Nfe2l2, while downregulating proinflammatory mediators Nfκ-B and Stat-1 (Fig. 5). These changes highlight a shift toward a regulated immune state that may help maintain hepatic function in the face of persistent infection.
The roles of Btg2, Nos2, and Nfe2l2 in liver health are particularly relevant.15 ^,^ 45 ^,^ 46 Btg2 is associated with antioxidant defense and reduced cellular proliferation30 while Nfe2l2 orchestrates the hepatic antioxidant response and offers cytoprotective effects during oxidative stress.46 Nos2 typically associated with nitric oxide production and antimicrobial defense, has a complex role in CD. During acute infection, Nos2 promotes parasite clearance and Th1 immune responses. However, in the chronic phase, sustained or excessive Nos2 expression can contribute to tissue damage, immune dysregulation, and fibrosis.45 In our study, we observed elevated Nos2 expression in both AIP-1 and AIP-2 treatment groups (Fig. 5C), yet this was not accompanied by increased fibrosis or hepatocellular damage. These findings suggest that AIP treatments may modulate Nos2 activity in a way that maintains host defense - possibly through oxidative stress regulation - while mitigating tissue injury.
Importantly, despite transcriptional upregulation of Nos2 we did not observe a reduction in liver parasite burden. This finding suggests that AIP-1 and AIP-2 do not act as antiparasitic agents per se but instead modulate the immune environment-potentially supporting host tolerance rather than resistance. This distinction is critical; while parasite control remains a therapeutic goal, preventing tissue damage is important to preserve tissue function in chronic CD.47
Supporting this, neither AIP-1 nor AIP-2 increased liver enzymes (ALT, AST, LDH, or ALP), reinforcing their hepatoprotective profile (Fig. 7). These results contrast sharply with previous observations where curative doses of BNZ significantly elevated liver enzymes, indicating hepatotoxicity.18 ^,^ 25 Thus, AIP-based therapies may offer a safer alternative or complement to antiparasitic treatments by mitigating tissue damage without compromising liver function.
While our findings suggest that AIP-1 and AIP-2 modulate immune responses without exacerbating liver pathology, several limitations remain. Further investigation is needed to determine whether the immune cells recruited from the liver are regulatory in nature. In particular, the role of MDSCs in the liver during T. cruzi infection requires clarification, as our current data are limited to splenic populations. STAT-1 protein and activation were not assessed, which will be important in future studies to fully understand IFN-γ-mediated signaling. Additional flow cytometry analyses are necessary to more precisely characterize the cellular composition of the hepatic infiltrate. Moreover, our study did not differentiate responses based on sex, and future comparisons between male and female mice will be important to assess potential sex-specific differences in immune modulation and treatment efficacy.
Trypanosoma cruzi infection is well known to trigger a robust pro-inflammatory response, often resulting in tissue damage and fibrosis, particularly within the liver.35 In our model, infection led to elevated parasite burden, increased cellular infiltration, and pronounced liver fibrosis - hallmarks of CD pathology - confirming that our model effectively replicates the characteristic disease features (Fig. 2). Interestingly, treatment with AIP-1 and AIP-2 further increased cellular infiltration without significantly reducing fibrosis (Fig. 2B-C), suggesting that these proteins may primarily influence immune cell recruitment rather than directly modulating fibrotic processes. In our previously published study evaluating the anti-inflammatory effects of AIP-1 and AIP-2 in the heart,20 we observed a significant reduction in cardiac inflammatory markers at 84 days post-infection, although only aspirin treatment significantly reduced cardiac fibrosis. Notably, we have also shown that cardiac fibrosis in this mouse model is progressive.48 Therefore, potential anti-fibrotic effects of AIP treatment may not yet be evident at this time point. Inclusion of additional time points - both immediately after treatment and at later intervals - would allow a more comprehensive understanding of the kinetics of fibrosis and immunomodulation. We plan to address this in future studies through time-course experiments designed to capture both the short- and long-term effects of AIP treatment.
In summary, our findings indicate that AIP-1 and AIP-2 act as immune modulators with potential antioxidant properties, helping to preserve liver structure and function during chronic T. cruzi infection. They promote immune cell infiltration - likely of regulatory and suppressor cell types - without exacerbating fibrosis, possibly through downregulation of pSTAT3, Stat1, and Nfκ-B, and upregulation of protective genes such as Btg2, Mmp9, Nos2, and Nfe2l2. Although these treatments do not reduce parasite burden, their capacity to fine-tune immune responses and prevent pathological inflammation highlights their potential as adjunctive therapies that could be combined with antiparasitic treatment to improve clinical outcomes in CD.
Conclusions and future directions - Overall, our findings suggest that AIP-1 and AIP-2 exert anti-inflammatory and antioxidant effects in chronic T. cruzi infection by reducing inflammatory signaling, promoting a balanced cytokine response, and preserving liver function. The data suggests that the immunomodulatory effects of may be mediated in part by regulatory DCs and MDSCs, which were increased in the spleen. Overall, this data provides promising insights into the therapeutic potential of these proteins. However, further studies are warranted to delineate their precise mechanisms of action and assess their long-term effects in chronic CD. Future research should also explore whether AIP-1 and AIP-2 can synergize with existing anti-parasitic therapies, such as BNZ, to enhance treatment efficacy and improve disease outcomes.
SUPPLEMENTARY MATERIALS
Supplementary material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO Chagas disease (also known as American trypanosomiasis)2025 https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis)
- 2Schmunis GA Yadon ZE Chagas disease a Latin American health problem becoming a world health problem Acta Trop 20101151-2142110.1016/j.actatropica.2009.11.00319932071 · doi ↗ · pubmed ↗
- 3Suárez C Nolder D García-Mingo A Moore DA Chiodini PL Diagnosis and clinical management of Chagas disease an increasing challenge in non-endemic areas Res Rep Trop Med 202213254010.2147/RRTM.S 34587935912165 PMC 9326036 · doi ↗ · pubmed ↗
- 4CDC Acute phase of Chagas disease 2010 https://www.cdc.gov/parasites/chagas/index.html
- 5Teixeira ARL Hecht MM Guimaro MC Sousa AO Nitz N Pathogenesis of Chagas disease parasite persistence and autoimmunity Clin Microbiol Rev 201124359263010.1128/CMR.00063-1021734249 PMC 3131057 · doi ↗ · pubmed ↗
- 6Basso B Modulation of immune response in experimental Chagas disease World J Exp Med 20133111010.5493/wjem.v 3.i 1.124520540 PMC 3905588 · doi ↗ · pubmed ↗
- 7Hoechst B Ormandy LA Ballmaier M Lehner F Krüger C Manns MP A new population of myeloid-derived suppressor cells in hepatocellular carcinoma patients induces CD 4+CD 25+Foxp 3+ T cells Gastroenterology 2008135123424310.1053/j.gastro.2008.03.02018485901 · doi ↗ · pubmed ↗
- 8Laloy-Borgna G Puyo L Nishino H Atlan M Catheline S Observation of natural flexural pulse waves in retinal and carotid arteries for wall elasticity estimation Sci Adv 202392510.1126/sciadv.adg 4287 PMC 1028454137343110 · doi ↗ · pubmed ↗