High-quality genome sequence of five Antarctic actinomycetes: Micromonospora ureilytica DSM 120150, Nocardiopsis akebiae DSM 120151, Streptomyces fildesensis DSM 41987T, Streptomyces hypolithicus DSM 41950T and Streptomyces albidoflavus DSM 120149
Ulrike Tarazona Janampa, Meike Döppner, Jolantha Swiderski, Imen Nouioui, Cathrin Spröer, Boyke Bunk, Yvonne Mast

TL;DR
This paper provides high-quality genome sequences for five Antarctic actinomycete bacteria, which could help in understanding their unique adaptations and potential applications.
Contribution
The study presents newly sequenced and high-quality genomes of five Antarctic actinomycete strains.
Findings
The genome sizes of the five strains range from 6.01 Mbp to 8.91 Mbp.
The genomes were sequenced to high quality, enabling detailed genetic analysis.
These sequences contribute to the understanding of Antarctic microbial diversity.
Abstract
Micromonospora ureilytica DSM 120150, Nocardiopsis akebiae DSM 120151, Streptomyces fildesensis DSM 41987T, Streptomyces hypolithicus DSM 41950T and Streptomyces albidoflavus DSM 120149 are five Antarctic strains. Here, we present the high-quality genome sequences of DSM 120150, DSM 120151, DSM 41987T, DSM 41950T and DSM 120149 with sizes of 7.51 Mbp, 6.90 Mbp, 8.91 Mbp, 6.01 Mbp and 6.85 Mbp, respectively.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
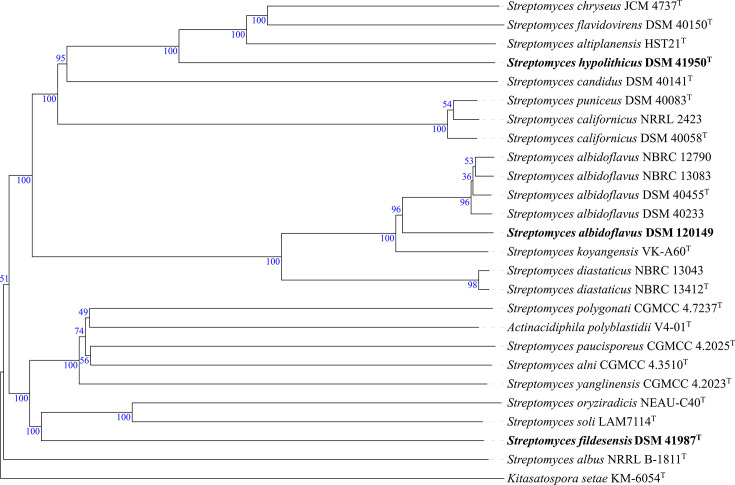 Fig. 1
Fig. 1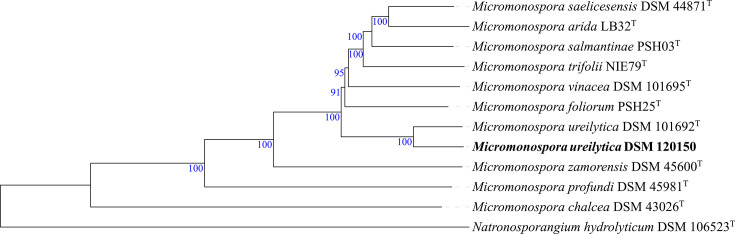 Fig. 2
Fig. 2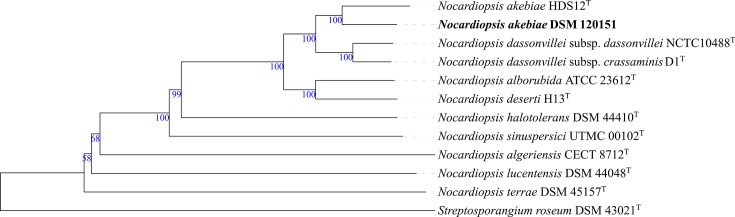 Fig. 3
Fig. 3 Fig. 4
Fig. 4| Attribute | Strain | ||||
|---|---|---|---|---|---|
| DSM 120150 | DSM 120151 | DSM 41987 | DSM 41950 | DSM 120149 | |
| Genome length (bp) | 7,508,249 | 6,900,836 | 8,913,051 | 6,014,151 | 6,852,435 |
| Contigs | 1 | 1 | 1 | 3 | 3 |
| Genome coverage | 76 × | 36 × | 101 × | 88 × | 121 × |
| Average G+C content (%) | 71 | 72.5 | 70.5 | 70 | 73.5 |
| CDS | 6,871 | 5,911 | 8,013 | 5,763 | 6,032 |
| tRNAs | 50 | 58 | 70 | 70 | 67 |
| rRNAs | 3, 3, 3 | 5, 5, 5 | 6, 6, 6 | 7, 7, 7 | 7, 7, 7 |
| RNA numbers | 62 | 76 | 91 | 94 | 91 |
| N50 (Mb) | 7.5 | 6.9 | 8.9 | 6 | 6.9 |
| CheckM completeness (%) | 97.86 | 99.01 | 100 | 99.28 | 99.43 |
| CheckM contamination (%) | 3.54 | 3.97 | 1.26 | 0 | 0.38 |
| Accession number | |||||
| BioProject number | PRJNA1293185 | PRJNA1293186 | PRJNA224116 | PRJNA1334162 | PRJNA1293190 |
- —http://dx.doi.org/10.13039/100021828 German Academic Exchange Service
- —http://dx.doi.org/10.13039/501100001664 Leibniz-Gemeinschaft
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Polar Research and Ecology · Microbial Natural Products and Biosynthesis
Data Summary
Genome sequence-related data availability is listed in Table 1 .
Announcement
The Antarctic habitat is one of the most unique and extreme environments on Earth [1]. Its harsh conditions, characterized by geographical and oceanographic isolation, freezing temperatures, long periods of darkness and limited accessibility, have created a habitat largely untouched by human influence [24]. The geographical and environmental isolation of Antarctica has allowed the development of highly specialized ecosystems, particularly in the benthic zones of the Southern Ocean, where life has adapted to thrive under some of the most challenging conditions on Earth [5]. Actinomycetes are one of the major sources of novel natural compounds [6]. Recent efforts in bioprospecting have focused on unique or extreme environments to increase the probability of finding structurally novel secondary metabolites [710]. As part of a bioprospecting effort targeting polar environments, five actinomycete strains were isolated from Antarctic quartz rock (DSM 41950^T^), soil (DSM 41987^T^) and marine sediment samples (DSM 120149, DSM 120150 and DSM 120151), and their genomes were sequenced.
Despite increasing interest in Antarctic micro-organisms, genome sequences of taxonomically well-characterized Antarctic actinomycetes, particularly type strains, remain limited. The five strains analysed here represent phylogenetically diverse actinomycetes isolated from distinct Antarctic habitats, including terrestrial, hypolithic and marine environments. The generation of high-quality genome sequences for these strains provides a valuable genomic resource that strengthens genome-based taxonomy and enables comparative analyses of biosynthetic gene cluster diversity across different actinomycete lineages.
Here, we present the annotated whole-genome sequences of five Antarctic strains, Micromonospora ureilytica DSM 120150, Nocardiopsis akebiae DSM 120151, Streptomyces fildesensis DSM 41987^T^, Streptomyces hypolithicus DSM 41950^T^ and Streptomyces albidoflavus DSM 120149, and describe their biosynthetic gene cluster content based on genome mining.
Strain DSM 120150 was cultivated on NL410 agar, strains DSM 120151 and DSM 120149 in R5 and S media and strains DSM 41950^T^ and DSM 41987^T^ in S-medium [1112], with all cultures grown in 10 ml medium at 28 °C with shaking at 180 r.p.m. After 3 days, biomass was harvested for subsequent DNA extraction. Genomic DNA extraction was carried out using the MasterPure Gram-positive DNA purification kit (Lucigen, Middleton, WI, USA) according to the manufacturer’s instructions. DNA integrity was checked on a FemtoPulse system (Agilent, Santa Clara, CA, USA). The genomes of the strains were sequenced using the PacBio Sequel-IIe platform (Pacific Biosciences, Menlo Park, CA, USA). SMRTbell template libraries were prepared according to the instructions from Pacific Biosciences outlined in the Procedure and Checklist – Preparing Multiplexed Microbial Libraries Using SMRTbell Express Template Prep Kit v.2.0 [13]. Briefly, for preparation of 10 kb libraries, 1 µg genomic DNA was sheared using the Megaruptor v.3 (Diagenode, Denville, NJ, USA) according to the manufacturer’s instructions. The DNA was end-repaired and ligated to barcoded adapters, applying components from the SMRTbell Express Template Prep Kit 2.0 (Pacific Biosciences). Samples were pooled according to the calculations provided by the Microbial Multiplexing Calculator (Pacific Biosciences). Conditions for primer annealing and polymerase binding to purified SMRTbell templates were assessed with the Calculator in SMRT Link (Pacific Biosciences). Libraries were sequenced using one 15 h movie per SMRT cell. SAMtools v.1.12 was used to obtain PacBio reads in FASTQ format [14]. The quality of the PacBio reads was confirmed using LongQC v.1.2.0c [15], and no error correction was performed. PacBio reads were de novo assembled using Flye v.2.8.1 (https://github.com/fenderglass/Flye) with the command ‘flye--pacbio-raw’, which included circularization of the contigs. The presence of plasmids was analysed using RFPlasmid v0.0.18 [16], and assembly quality was evaluated using QUAST v4.4 [17]. The genome sequences of M. ureilytica DSM 120150, N. akebiae DSM 120151, S. fildesensis DSM 41987^T^, S. hypolithicus DSM 41950^T^ and S. albidoflavus DSM 120149 were deposited in GenBank under accession numbers CP196338 (DB10_83), CP196194 (M1B1), CP189853, CM128811 and CP196343 (DB7_152), respectively. Genome annotation was carried out with the NCBI Prokaryotic Genome Annotation Pipeline [18] (Table 1). Genome completeness and contamination were assessed with CheckM v1.0.18 [19]. All strains showed genome completeness above 97.86% and contamination below 3.97% (Table 1).
The phylogenetic relationship of the strains was analysed with the EzBioCloud server (https://www.ezbiocloud.net) [20], based on 16S rRNA gene sequences extracted from the genomes. The authenticity of the strains was confirmed by comparing genome-derived 16S rRNA gene sequences with those obtained by PCR. 16S phylogenetic analysis revealed that strains DSM 120150, DSM 120151, DSM 41987^T^, DSM 41950^T^ and DSM 120149 were most similar to Micromonospora arida LB32^T^ (MG725912) with 99.8% similarity [21], Nocardiopsis dassonvillei subsp. crassaminis D1^T^ (LR606207) with 99.91% [22], S. fildesensis GW25-5^T^ (DQ408297) with 99.72% [23], S. hypolithicus HSM10^T^ (EU196762) with 99.87% [24] and Streptomyces pyxinicus LP11^T^ (OL765279) with 98.89% similarity [25], respectively.
Phylogenomic analysis based on whole-genome sequences and digital DNA–DNA hybridization (dDDH) (formula d_4_) was carried out with the Type Strain Genome Server [2627]. The phylogenomic tree constructed from the Streptomyces genome sequences positioned strain S. hypolithicus DSM 41950^T^ in close association with Streptomyces altiplanensis HST21^T^, Streptomyces flavidovirens DSM 40150^T^ and Streptomyces chryseus JCM 4737^T^ [2830], forming a well-supported subcluster, with dDDH values ranging from 29.9 to 31.1%. This study reports, for the first time, the genome sequence of S. hypolithicus DSM 41950^T^, designated as the type strain of S. hypolithicus in the original taxonomic description by Le Roes-Hill et al. [24] [24]. The strain S. albidoflavus DSM 120149 formed a distinct branch, closely related to a subcluster that included the type strain S. albidoflavus DSM 40455ᵀ [31], with a dDDH value of 64.7%. This subcluster also encompassed Streptomyces limosus NBRC 12790^T^, Streptomyces sampsonii NBRC 13083^T^ and Streptomyces coelicolor DSM 40233^T^ [3133], with dDDH values ranging from 64.3 to 64.6%. All these strains are heterotypic synonyms of S. albidoflavus. To further assess the taxonomic position of DSM 120149, the average nucleotide identity (ANI) was analysed using the EzBioCloud tool ANI Calculator [34]. Strain DSM 120149 shared an ANI value of 95.97% with DSM 40455^T^, which is at the threshold for prokaryotic species delineation (95–96%) [35]. Strain S. fildesensis DSM 41987^T^ formed a distinct branch, closely related to a well-supported subclade comprising Streptomyces soli LAM7114^T^ and Streptomyces oryziradicis NEAU-C40^T^ [3637], with dDDH values of 23.4% and 23.0%, respectively (Fig. 1). The strain DSM 41987^T^ was originally designated as the type strain of S. fildesensis by Li et al. [23] [23]. All dDDH values reported in this study were below the 70% species demarcation threshold, consistent with their assignment as distinct species. The Antarctic Micromonospora strain DSM 120150 formed a distinct branch closely related to M. ureilytica DSM 101692^T^ [38] (Fig. 2), with a dDDH value of 77.3%. In the Nocardiopsis phylogenomic tree, strain N. akebiae DSM 120151 formed a well-supported subbranch closely related to N. akebiae HDS12^T^ [39] (Fig. 3), with a dDDH value of 69.6% and an ANI value of 96.44%. The dDDH and ANI values between strains DSM 120150 and DSM 120151 and their closest relatives support their assignment to M. ureilytica and N. akebiae, respectively, although the borderline dDDH value observed for strain DSM 120151 suggests that additional genomic and phenotypic analyses would be required to further resolve its taxonomic status.
Phylogenetic tree of S. fildesensis DSM 41987ᵀ, S. hypolithicus DSM 41950ᵀ, and S. albidoflavus DSM 120149 inferred with FastME 2.1.6.1 [41] from Genome BLAST Distance Phylogeny (GBDP) distances calculated from genome sequences. The branch lengths are scaled in terms of the GBDP distance formula d5. The tree was rooted at the midpoint [42] and visually edited using Interactive Tree of Life (iTOL) v.7.2.1 [43].
Phylogenetic tree of M. ureilytica DSM 120150 inferred with FastME 2.1.6.1 [41] from GBDP distances calculated from genome sequences. The branch lengths are scaled in terms of the GBDP distance formula d5. The tree was rooted at the midpoint [42] and visually edited using Interactive Tree of Life (iTOL) v.7.2.1 [43].
Phylogenetic tree of N. akebiae DSM 120151 inferred with FastME 2.1.6.1 [41] from GBDP distances calculated from genome sequences. The branch lengths are scaled in terms of the GBDP distance formula d5. The tree was rooted at the midpoint [42] and visually edited using Interactive Tree of Life (iTOL) v.7.2.1 [43].
To investigate the genetic potential for secondary metabolite production, the genome sequences of the five Antarctic strains were analysed with the bioinformatic web tool antiSMASH v.8.0 [40] for the abundance of biosynthetic gene clusters (BGCs). A total of 21, 20, 30, 30 and 24 BGCs were identified for strains DSM 120150, DSM 120151, DSM 41987^T^, DSM 41950^T^ and DSM 120149, respectively (Fig. 4a–e). The number and diversity of identified BGCs indicate a broad genetic repertoire associated with secondary metabolism.
AntiSMASH output from Antarctic actinomycetes strains (a) M. ureilytica DSM 120150, (b) N. akebiae DSM 120151, (c) S. fildesensis DSM 41987T, (d) S. hypolithicus DSM 41950T and (e) S. albidoflavus DSM 120149. Colour code according to antiSMASH v.8.0 [40].
This study provides high-quality genome sequence data for the five Antarctic actinomycete strains M. ureilytica DSM 120150, N. akebiae DSM 120151, S. fildesensis DSM 41987^T^, S. hypolithicus DSM 41950^T^ and S. albidoflavus DSM 120149 and their BGC cluster content associated with secondary metabolism.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Avila C Biological and chemical diversity in Antarctica: from new species to new natural products Biodiversity 20161751110.1080/14888386.2016.1176957 · doi ↗
- 2Sivalingam P Hong K Pote J Prabakar K Extreme environment Streptomyces: potential sources for new antibacterial and anticancer drug leads?Int J Microbiol 20192019528394810.1155/2019/528394831354829 PMC 6636559 · doi ↗ · pubmed ↗
- 3Liao L Su S Zhao B Fan C Zhang J et al Biosynthetic potential of a novel antarctic actinobacterium Marisediminicola antarctica ZS 314T revealed by genomic data mining and pigment characterization Mar Drugs 20191711510.3390/md 17070388 PMC 666964431266176 · doi ↗ · pubmed ↗
- 4Núñez-Pons L Avila C Romano G Verde C Giordano D UV-Protective Compounds in Marine Organisms from the Southern Ocean Mar Drugs 20181633610.3390/md 1609033630223486 PMC 6165330 · doi ↗ · pubmed ↗
- 5Li A-Z Han X-B Zhang M-X Zhou Y Chen M et al Culture-dependent and -independent analyses reveal the diversity, structure, and assembly mechanism of benthic bacterial community in the Ross Sea, Antarctica Front Microbiol 201910252310.3389/fmicb.2019.0252331787942 PMC 6856632 · doi ↗ · pubmed ↗
- 6Mast Y Stegmann E Actinomycetes: the antibiotics producers Antibiotics 2019810510.3390/antibiotics 803010531362405 PMC 6783997 · doi ↗ · pubmed ↗
- 7Subramani R Sipkema D Marine rare actinomycetes: a promising source of structurally diverse and unique novel natural products Mar Drugs 20191724910.3390/md 1705024931035452 PMC 6562664 · doi ↗ · pubmed ↗
- 8Nouioui I Boldt J Zimmermann A Makitrynskyy R Pötter G et al Biotechnological and pharmaceutical potential of twenty-eight novel type strains of Actinomycetes from different environments worldwide Curr Res Microb Sci 2024710029010.1016/j.crmicr.2024.10029039497933 PMC 11533595 · doi ↗ · pubmed ↗