Ecotypic Variation in Photosynthesis, Stomatal Conductance, and Water Use Efficiency of Illicium lanceolatum in Response to Light Intensity Under Drought and Recovery
Yonghui Cao, Benzhi Zhou

TL;DR
This study examines how different ecotypes of Illicium lanceolatum respond to drought and light intensity, identifying more drought-tolerant varieties for conservation and cultivation.
Contribution
The study reveals ecotypic variation in drought tolerance and water use efficiency under different light intensities in Illicium lanceolatum.
Findings
Short-term drought increased photosynthetic efficiency in I. lanceolatum ecotypes.
The WN and KH ecotypes showed improved water use efficiency under drought and high light.
Stomatal conductance and water use efficiency varied significantly among ecotypes during rewatering.
Abstract
Increasingly frequent extreme droughts threaten forest vegetation and highlight the need to identify drought-tolerant germplasm. To support conservation and cultivation of Illicium lanceolatum, we investigated ecotypic differences in photosynthetic responses to short-term drought and rewatering under varying light intensity. One-year-old seedlings from four I. lanceolatum ecotypes originating from the Zhejiang (Lin’an, LA; Kaihua, KH), Jiangxi (Wu’ning, WN), and Fujian (Nan’ping, NP) provinces in China were subjected to drought stress by withholding irrigation and subsequent rewatering. Photosynthesis–light response curves were measured before drought; 2, 4, and 7 days after the last watering; and following rewatering. Short-term drought significantly affected photosynthetic traits in an ecotype-dependent manner. Maximum net photosynthetic rate, light saturation point, light…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
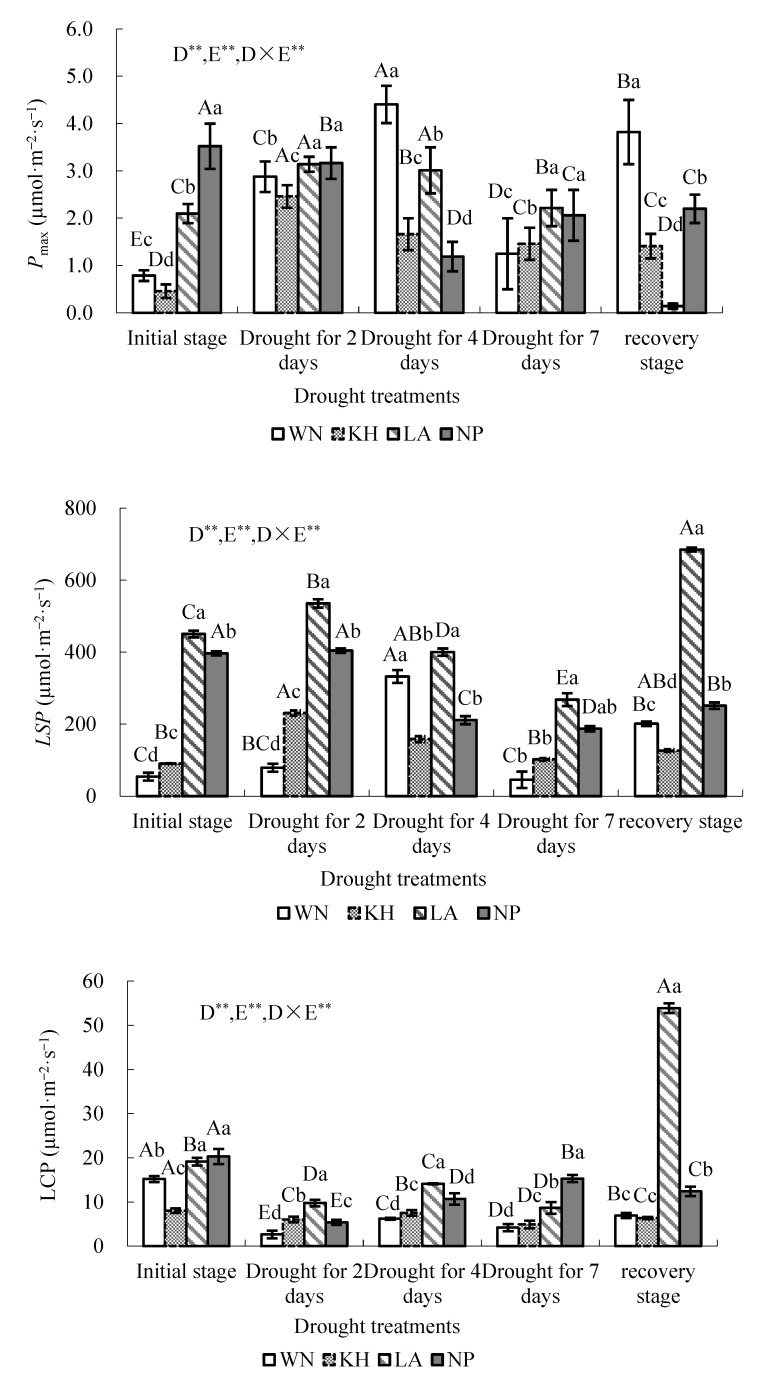 Figure 1
Figure 1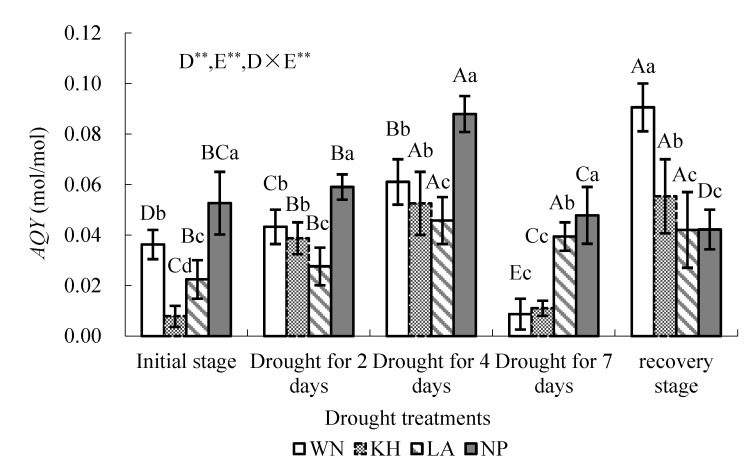 Figure 2
Figure 2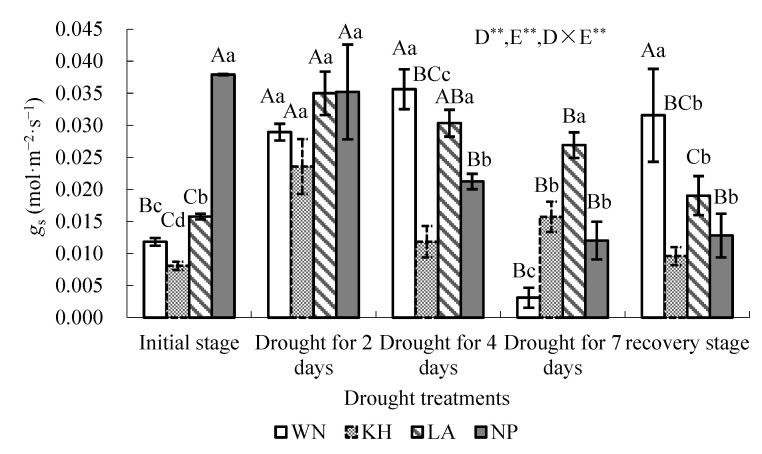 Figure 3
Figure 3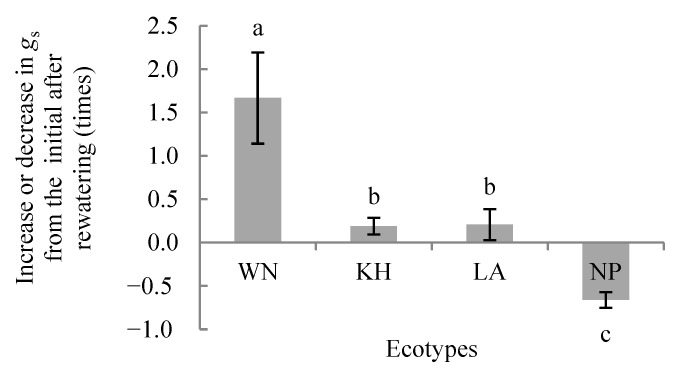 Figure 4
Figure 4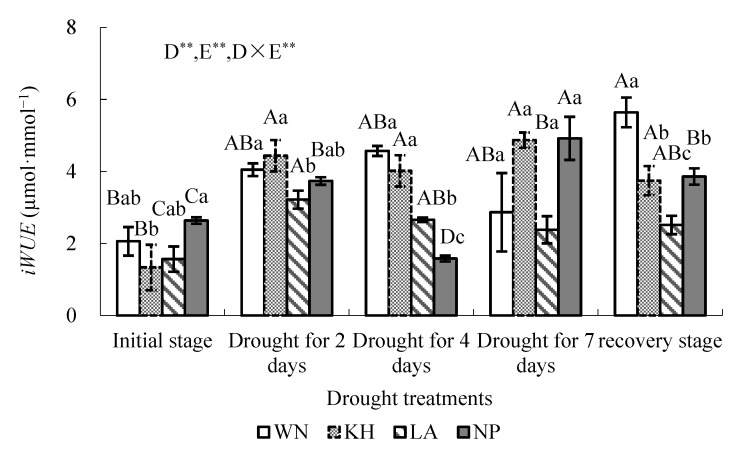 Figure 5
Figure 5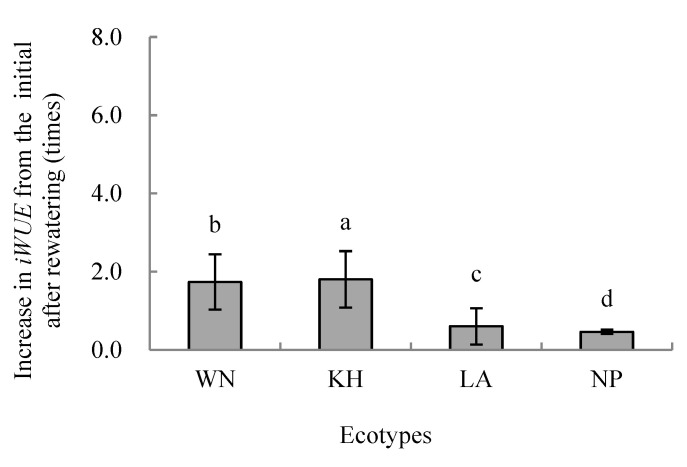 Figure 6
Figure 6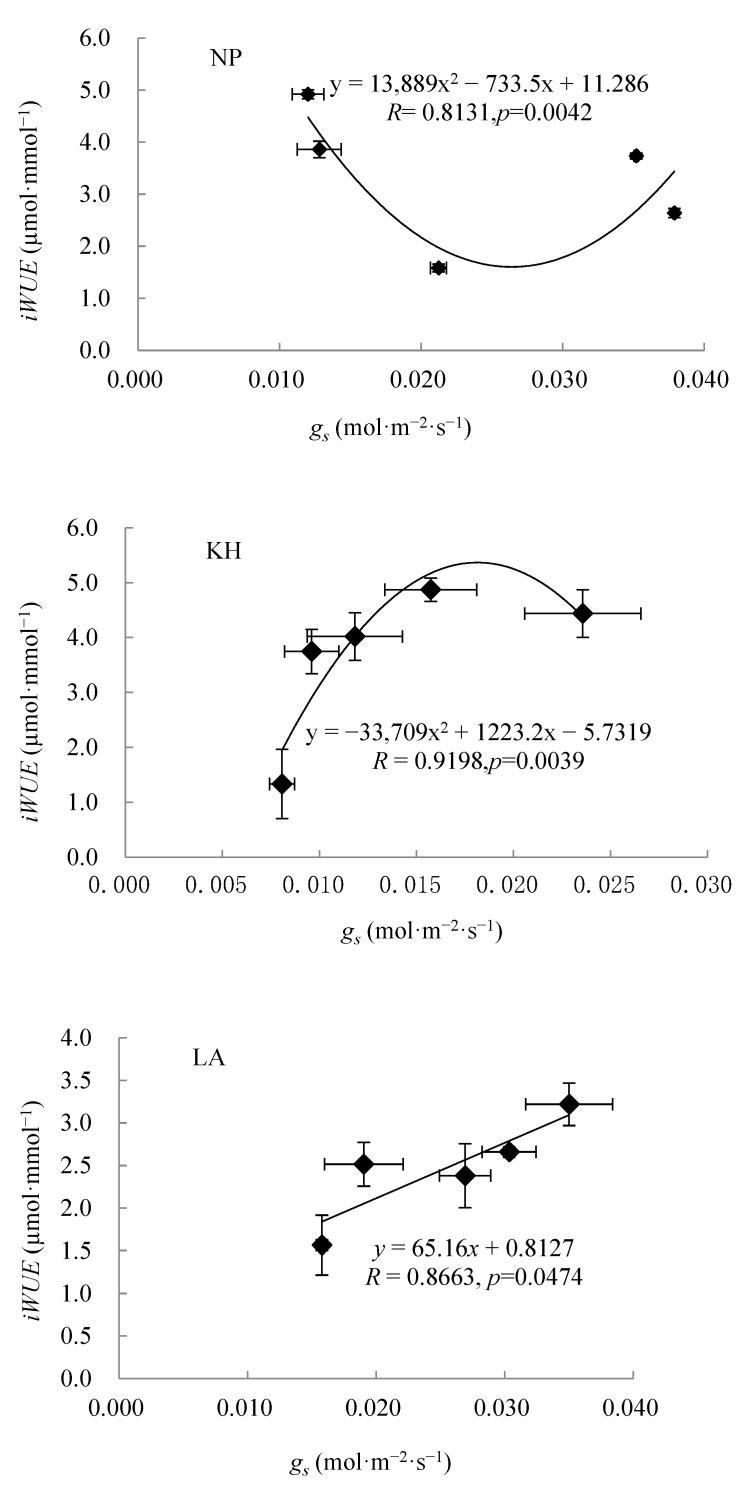 Figure 7
Figure 7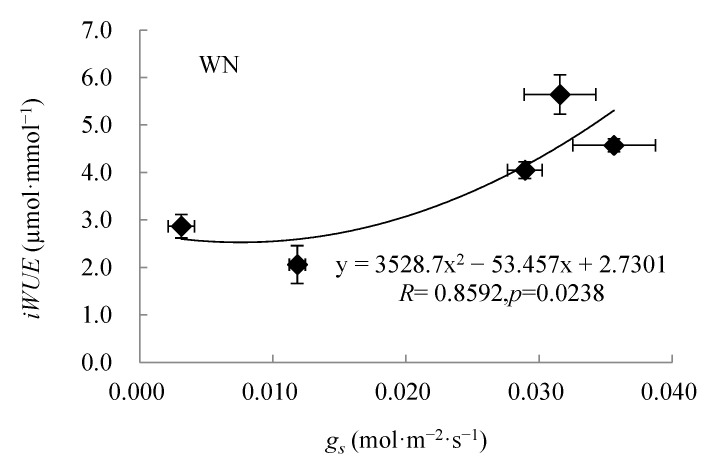 Figure 8
Figure 8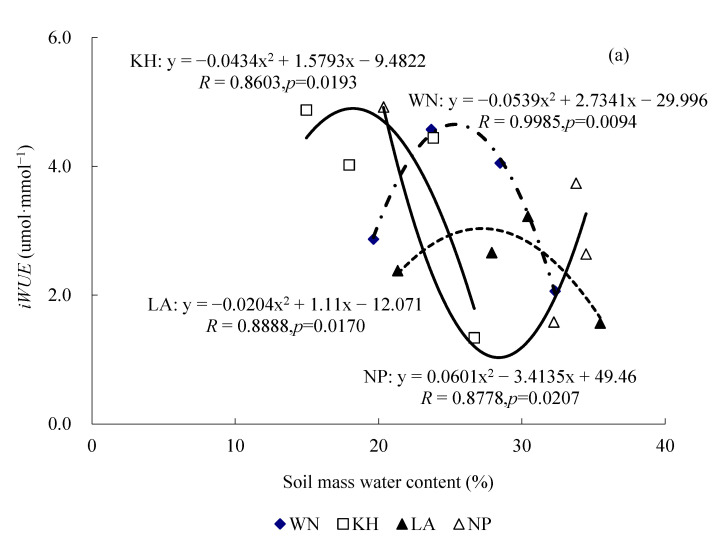 Figure 9
Figure 9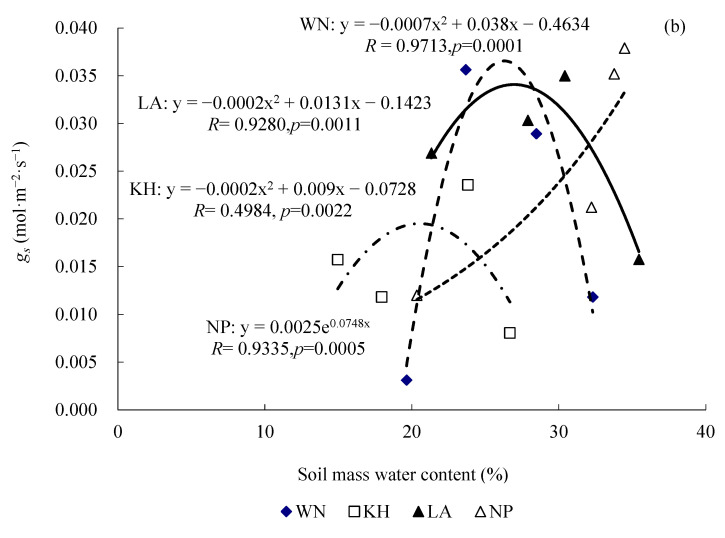 Figure 10
Figure 10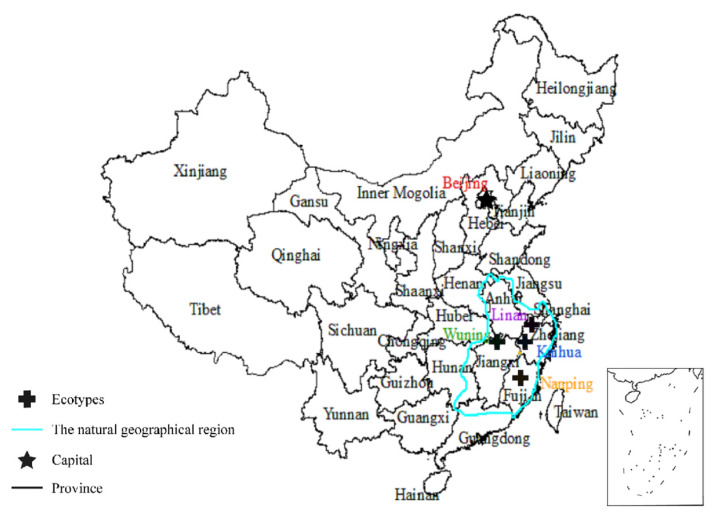 Figure 11
Figure 11- —General program of the Natural Science Foundation of China
- —Special Fund for Basic Scientific Research from Central-level Non-profit Scientific Research Institutes of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMagnolia and Illicium research · Plant Water Relations and Carbon Dynamics · Irrigation Practices and Water Management
1. Introduction
The increasing frequency and intensity of drought events driven by global climate change pose serious threats to plant diversity, ecosystem productivity, and species distribution worldwide [1,2,3,4]. Adjusting their physiological traits to cope with water scarcity is, therefore, essential for elucidating mechanisms of drought adaptation and resilience [5,6,7]. Among these traits, stomatal regulation plays a central role in balancing carbon gain and water loss. As the gatekeepers of leaf gas exchange, stomata strongly influence plant performance under conditions of elevated temperature and drought stress [8,9]. In general, stomata respond to declining soil water availability and increasing atmospheric vapor pressure deficit by closing [10,11], which reduces stomatal conductance (gₛ) and limits carbon assimilation [9,12,13]. Under drought conditions associated with approximately 30% reduction in annual precipitation, tree net CO_2_ assimilation decreased by 49%, whereas stomatal conductance, transpiration rate, and water use efficiency remained largely unchanged [14]. This indicates that plant species exhibit distinct and dynamic physiological responses to drought. While many studies in wheat report reductions in g_s_ under heat or drought stress [15,16], other studies show that g_s_ responses can vary markedly depending on environmental conditions, growth stage, and genotype [9]. These contrasting responses suggest that plant water use strategies are highly dynamic and context-dependent [17].
Water use efficiency (WUE) is a key indicator of plant drought adaptation [11]. Instantaneous water use efficiency (iWUE), defined as the ratio of carbon assimilation to transpiration, reflects how effectively plants assimilate carbon under water-limited conditions and is particularly informative during drought [6,13,18,19,20,21]. Improving iWUE is, therefore, critical for enhancing plant performance under future climate scenarios [22,23]. Although drought often reduces g_s_ and iWUE [21], moderate drought can increase iWUE by constraining water loss more strongly than carbon gain [24], whereas severe drought generally leads to declines in both traits [25,26].
Drought stress directly affects key determinants of plant productivity, including canopy structure, photosynthesis, and assimilate allocation. Consequently, identifying drought-tolerant ecotypes is increasingly important under ongoing climatic variability [27]. While intraspecific variation in drought tolerance is recognized as a major component of plant adaptation to climate change [4,28], such variation remains poorly characterized in many woody species. Widely distributed species often exhibit pronounced ecotypic differentiation in functional traits due to local environmental conditions [29,30], yet comparative studies examining ecotypic variation in iWUE and stomatal regulation under drought are still limited.
Illicium lanceolatum A.C. Smith, a medicinal plant commonly known as poisonous eight-angle [31], is an endemic evergreen tree species restricted to southern China. It is an important medicinal and aromatic plant, valued for its anti-inflammatory and analgesic properties and as a major natural source of shikimic acid, a key precursor of the antiviral drug oseltamivir (Tamiflu) [32,33]. Due to overharvesting and habitat degradation, I. lanceolatum has become an endangered species in China, highlighting the urgent need for conservation, germplasm protection, and large-scale artificial cultivation.
Adaptation to macro- and micro-climatic variation may depend on genetic differentiation among ecotypes [34]. Increasing water scarcity poses a major challenge for cultivated tree species such as Illicium lanceolatum. Consequently, identifying genotypes with high leaf iWUE [13,35]) is crucial for improving the sustainability of future artificial forests. In order to improve iWUE in cultivated plants, Gilbert et al. (2011) proposed either reducing g_s_ under well-watered conditions or enhancing stomatal responsiveness in the early stages of drought stress [36]. However, how the coupled effects between the drought and the light intensity influence g_s_ and iWUE across different I. lanceolatum ecotypes remains largely unexplored.
In its natural habitats, I. lanceolatum occurs discontinuously in valley broad-leaved forests at elevations of 600 to 1000 m, often growing in shaded, moist, and fertile soils alongside species such as Phoebe sheareri Phoebe sheareri, Michelia platypetala, and Manglietia chingii [37]. Although the species is shade-tolerant and morphologically robust, wild populations have declined sharply due to intensive exploitation. Despite its ecological and economic importance, experimental studies examining the physiological mechanisms underlying its responses to water and light availability—particularly under common garden conditions—remain scarce.
Stomatal regulation is a key mechanism by which plants mitigate drought stress [37,38]. Drought-tolerant genotypes often exhibit lower g_s_ under well-watered conditions or enhanced stomatal sensitivity during early drought, thereby conserving soil water and maintaining higher iWUE [39]. However, how g_s_ and iWUE jointly respond to the combined effects of drought and light intensity across different I. lanceolatum ecotypes remains unclear. In this study, we compared the dynamic responses of photosynthetic traits, g_s_ and iWUE among four I. lanceolatum ecotypes under varying drought durations and subsequent rewatering. Our objectives were to (1) evaluate the sensitivity of photosynthesis, g_s_, and iWUE to soil water availability under different light conditions and (2) identify ecotypic differences in drought tolerance. Ultimately, this work aims to select drought-tolerant ecotypes suitable for future cultivation and afforestation under climate change, thereby supporting ex situ conservation and sustainable utilization of I. lanceolatum genetic resources.
2. Results
2.1. Effects of Drought Stress on Photosynthetic Properties in Four I. lanceolatum Ecotypes
Photosynthetic parameters showed clear ecotypic differences during the short-term drought stress (DS) and subsequent recovery (Figure 1). The maximum net photosynthetic rate (Pmax), which reflects the maximum photosynthetic capacity of leaves under optimal conditions [40,41], was significantly influenced by drought treatment, ecotype, and their interaction (two-way variance analysis, p < 0.01). Under the initial water stage, after 4 days since the last watering, and after rewatering, Pmax differed significantly among the four ecotypes (p < 0.05). After 2 days and 7 days without watering, Pmaxs of the LA and NP ecotypes were significantly higher than those of WN and KH (p < 0.05). The temporal dynamics of Pmax varied among ecotypes. In the LA and KH ecotypes, Pmax increased rapidly soon after 2 days of drought and then declined (Figure 1). In contrast, Pmax in the WN ecotype continued to increase and reached the highest level after 4 days of drought. Following rewatering, Pmax continued to decrease in the LA and KH ecotypes, whereas it increased in the WN ecotype.
During the DS treatment, Pmax of the four ecotypes was positively correlated with soil water content (Wm). LSD analysis showed that Pmax of the WN seedlings differed significantly between the DS treatment period and the initial watering stage (p < 0.05), with the NP ecotype showing the highest value, followed by LA ecotype. After 2 days of drought, Pmax increased significantly in all ecotypes except NP, and no significant difference was observed between LA and NP, which maintained the highest Pmax level. After 4 days of drought, Pmax, again, differed significantly among all ecotypes (p < 0.05). These results indicate that ecotypes differ markedly in the magnitude and temporal patterns of Pmax responses to drought, likely reflecting distinct stomatal and physiological adjustment strategies under progressive water limitation.
Light saturation points (LSPs) and light compensation points (LCPs) reflect a plant’s ability to utilize high and low light intensities, respectively [40]. Two-way variance analysis showed that drought treatments, ecotype, and their interaction significantly affected LSPs and LCPs across the four ecotypes (p < 0.01) (Figure 1).
Under initial watering, after 2 days of drought, and after rewatering, LSPs differed significantly among the four ecotypes (p < 0.05). After 4 days of drought, the LSPs of WN and LA ecotypes were significantly higher than those of KH and NP, while after 7 days, the LA ecotype had the highest LSP (p < 0.05). Wm decreased to 23.81%–33.77% after 2 days of drought (Table 1), during which LSPs increased in all ecotypes. After 4 days, WN seedlings reached the highest LSP (332.30 µmol·m^−2^·s^−1^), whereas the other ecotypes declined as drought progressed. Across all drought stages, the LSP of LA remained consistently higher than the other ecotypes. Rewatering restored LSPs, which exceeded initial values in WN, LA, and KH. LSD analysis indicated significant differences between drought and initial water conditions for specific ecotypes at different time points: LA at all stages, NP after 4 days, WN on day 4, and KH on day 2 (p < 0.05).
LCP, the light intensity at which photosynthesis balances respiration [42], decreased during drought and recovered after rewatering [42], with a marked increase observed only in LA. Initially, LCPs of LA and NP were higher than WN and KH (p < 0.05). As drought progressed, LCPs differed significantly among ecotypes (p < 0.05). After rewatering, LA maintained the highest LCP, while LA and NP were significantly higher than WN and KH.
The apparent photo quantum efficiency (AQY) reflects the efficiency of light utilization under low light [43]. Two-way variance analysis revealed that drought, ecotype, and their interaction significantly affected AQY (p < 0.01; Figure 1). AQY increased in all ecotypes until day 4 of drought. LSD analysis showed that AQY of NP was significantly lower than initial values (p < 0.05), whereas AQY of LA, WN, and KH increased significantly (p < 0.05). Initially, AQY differed significantly among ecotypes (p < 0.05). As drought progressed, differences between WN and KH became nonsignificant. After 2 and 4 days, WN and KH had higher AQY than LA but lower than NP (p < 0.05). By day 7, WN and KH had significantly lower AQY than LA and NP. After rewatering, AQY in WN and KH remained higher than in LA and NP. These results indicate that LSP, LCP, and AQY exhibit ecotype-specific dynamic responses to drought, reflecting differences in light use strategy and stomatal adaptation among I. lanceolatum ecotypes.
2.2. Effect of Drought Stress on Stomatal Conductance of I. lanceolatum Ecotypes
The variation of average gₛ under PAR > 1500 µmol·m^−2^·s^−1^ for the four ecotypes during drought stress is shown in Figure 2. Two-way variance analysis showed that drought treatments, ecotype, and their interaction significantly affected the gₛ across the experimental period (p < 0.01). LSD analysis revealed that gₛ differed significantly both among water treatments within the same ecotype and among ecotypes under the same water treatment.
At the initial watering stage, NP exhibited the highest gₛ, while KH had the lowest (Figure 2). During drought, gₛ trends varied among ecotypes. In NP, gₛ decreased continuously with drought and remained below initial levels after rewatering (p < 0.05), indicating strong limitation by soil moisture under high light. In contrast, WN gₛ increased rapidly during short-term drought (Figure 3), peaking at 4 days when soil water content dropped to 19.64 ± 0.34% (Table 1), then declined sharply by day 7. After rewatering, gₛ recovered significantly above initial levels (p < 0.05).
For the LA ecotype, gₛ increased rapidly during the first 2 days of drought, then declined as drought progressed, and partially recovered after rewatering, with significant differences compared to the initial water stage (p < 0.05). In the NP ecotype, gₛ decreased rapidly during drought and increased slightly after rewatering, but remained significantly lower than the initial water stage (p < 0.05), indicating that soil moisture strongly limits gₛ under high light (>1500 µmol·m^−^^2^·s^−^^1^). In KH, gₛ initially increased during early drought, then declined sharply, reaching a minimum at 7 days when soil water content dropped to 14.96 ± 0.09% (Table 1), and slightly exceeded initial levels after rewatering. For both LA and KH ecotypes, gₛ recovered and showed a slight increase after rewatering compared with the initial stage, although these changes were not statistically significant (p > 0.05). Overall, under high light (>1500 µmol·m^−^^2^·s^−^^1^), the ecotypes displayed distinct gₛ dynamics in response to soil water limitation. WN and LA showed rapid short-term increases in gₛ during early drought, while NP was consistently constrained by low soil moisture, and KH exhibited an intermediate response.
2.3. Effect of Drought Stress on Water Use Efficiency of I. lanceolatum Ecotypes
Two-way variance analysis showed that drought treatments, ecotype and their interaction significantly affected the iWUE of each ecotype (p < 0.01) (Figure 4). LSD analysis revealed that at the initial watering stage, NP had the highest iWUE, while KH and LA were lower, with a significant difference only between KH and NP (p < 0.05) (Figure 4). During short-term drought, iWUE increased in all ecotypes except NP on day 4, which was slightly lower than its initial value. By day 7, iWUE of KH, LA, and NP had increased significantly compared with initial conditions (p < 0.05), whereas WN showed no significant change. After rewatering, iWUE of all ecotypes increased significantly above initial values (p < 0.05; Figure 4), with WN and KH exhibiting the highest iWUE (Figure 5).
Temporal patterns varied among ecotypes. KH showed a prolonged rise in iWUE after drought, followed by WN, whose iWUE decreased by day 7 but rebounded after rewatering to exceed initial levels (Figure 4). LA consistently displayed lower iWUE than WN and KH during short-term drought. These results indicate that under short-term soil drought, higher light conditions promote iWUE in all ecotypes. Moreover, the relative increase in iWUE from the initial stage after rewatering differed significantly among ecotypes (Figure 5), highlighting ecotype-specific adaptation strategies in carbon assimilation and water use under drought and recovery.
2.4. Relationship Between Stomatal Conductance and Water Use Efficiency
Under PAR exceeding 1500 µmol·m^−2^·s^−1^, a significant polynomial relationship was observed between mean stomatal conductance gₛ and iWUE in seedlings of the NP, KH, and WN ecotypes of I. lanceolatum across different water treatments. The correlation coefficients (R) ranged from 0.8131 to 0.9198, indicating a strong association between these two physiological parameters (Figure 6). For the WN and NP ecotypes, iWUEs initially decreased and subsequently increased with increasing gₛ, although the magnitude of variation differed between the two ecotypes. In contrast, iWUE in the KH ecotype increased rapidly at the beginning and then gradually as gₛ increased further. By comparison, iWUE in the LA ecotype exhibited a continuous increase with increasing gₛ.
2.5. Responses of iWUE and gₛ to Change of Water Stress Under Saturating Light
The results indicate pronounced ecotypic differences in the responses of iWUE to the light changes in water status under short-term drought stress and subsequent rewatering. Under saturating light conditions (>1500 umol·m^−2^·s^−1^), significant polynomial relationships were observed between Wm and iWUE in seedlings of all four Illicium lanceolatum ecotypes (Figure 7a). Specifically, iWUE in the KH, LA, and WN ecotypes initially increased and then decreased, along with the increase in Wm, whereas the NP ecotype exhibited the opposite trend, showing an initial decrease followed by an increase (Figure 7a). The polynomial relationships were significantly significant (R was 0.8603, 0.8888, 0.9985, and 0.8778 for KH, LA, WN, and NP, respectively), suggesting that iWUE is highly sensitive to changes in soil water status under high light conditions. Overall, these results suggest that an optimal water supply under saturating light enhances iWUE in the KH, LA, and WN ecotypes, while excessive water availability reduces water use efficiency.
Under the saturating light conditions, significant polynomial function correlations were also detected between Wm and gₛ for the WN, LA, and KH ecotypes (Figure 7b); R ranged from 0.4984 to 0.9713, p < 0.01. In contrast, the correlation between Wm and gₛ for the NP ecotype belongs to a significant exponential function (R = 0.9335, p = 0.0005). These findings suggest that gₛ in all four ecotypes is more sensitive to Wm under the saturating light intensity (>1500 µmol·m^−2^·s^−1^).
3. Discussion
3.1. Responses of I. lanceolatum Photosynthetic Parameters to Drought Stress
Climate change is expected to reduce annual precipitation and prolong dry summers, thereby posing increasing threats to forest ecosystems [14]. Drought stress primarily constrains photosynthesis by inducing stomatal closure and, under severe conditions, by causing damage to palisade mesophyll cells [37,44,45,46,47,48]. Plants with a low LCP and a high LSP generally exhibit greater adaptability to fluctuating light environments; therefore, selecting genotypes capable of tolerating a wide range of light conditions is critical for plant survival under extreme climatic scenarios [45,49]. In the present study, the four Illicium lanceolatum ecotypes exhibited distinct strategies of photosynthetic adjustment in response to progressive drought stress. The LSP of the WN ecotype increased initially and then decreased with advancing drought, reaching a maximum value significantly higher than that under the initial water supply after 4 days of drought. In contrast, the LSP of the KH, LA, and NP ecotypes significantly increased after 2 days of drought and subsequently declined. These patterns indicate ecotype-specific drought adaptation strategies mediated through dynamic regulation of LSP. Notably, the WN ecotype maintained a relatively high LSP for a longer drought duration, suggesting an enhanced ability to utilize high light under water-limited conditions.
Previous studies have reported that drought can reduce photosynthesis by 29.5% to 57.7% for Tilia amurensis Rupr. seedlings [5]. In this study, the timing and magnitude of the decline in Pmax differed markedly among ecotypes. The NP ecotype experienced the earliest reduction in Pmax, occurring after only 2 days of drought, while the WN ecotype had the latest decrease, which occurred after 7 days of drought and remained higher than the initial well-watered level. These contrasting responses likely reflect differences in stomatal regulation and hydraulic behavior among ecotypes, although the underlying mechanisms of stomatal dynamics require further investigation.
Consistent with Li et al. (2019) [44], our study reveals a progressive sequence of leaf photophysiological adjustments in response to increasing drought severity (Figure 1, Figure 2 and Figure 4). Clear ecotypic differences were observed in photosynthetic traits throughout the drought treatment. For the WN ecotype, both Pmax and the LSP peaked when Wm dropped to 23.67%, after which they declined (Figure 1, Table 1). In contrast, the threshold Wm values corresponding to declines in Pmax and LSP were 23.81% and 30.41% for the KH and LA ecotypes, respectively (Figure 1, Table 1). These findings indicate that the photosynthetic activities in I. lanceolatum seedlings is suppressed when soil moisture drops below an ecotype-specific threshold. Such thresholds provide valuable reference points for optimizing irrigation regimes in artificial cultivation and for defining drought-tolerance limits.
Following rewatering, Pmax increased markedly in the WN ecotype, whereas it continued to decline or recovered slowly in the LA and KH ecotypes. These results suggest that the LA ecotype is more sensitive to drought and high light stress and exhibits limited recovery capacity, whereas the WN ecotype demonstrates both reduced photosynthetic inhibition during drought and strong resilience upon rehydration. Previous metabolomic studies have shown that drought induces the accumulation of amino acids and sugars, implicating these pathways in stress tolerance [50]. The superior post-drought recovery of the WN ecotype observed here may, therefore, be associated with specific metabolic adjustments, a hypothesis that warrants further investigation.
Climatic data from the native habitats of the four ecotypes (Table 2) reveal that the WN ecotype originates from regions with significantly lower annual precipitation than those of the other ecotypes (p < 0.05). This long-term exposure to water-limited environments likely underlies its enhanced drought tolerance and recovery capacity. In contrast, although the annual mean temperatures of the WN, KH, and LA habitats are comparable, the NP ecotype originates from a significantly warmer region, which may explain its relatively slower recovery following drought stress.
Rewatering restored the LSP and LCP in all ecotypes, with LSP values in the WN, KH, and LA ecotypes exceeding those under initial water supply conditions. Moreover, AQY increased significantly after drought stress in all ecotypes except NP (p < 0.05), indicating that short-term drought enhanced low-light utilization efficiency. These results provide important guidance for optimizing light conditions in I. lanceolatum cultivation.
The ecotypic differences observed in this study are consistent with previous reports in other woody species, such as olive (Olea europaea L.) [45] and European beech (Fagus sylvatica L.) [51], which have demonstrated contrasting drought sensitivity and recovery among genotypes. Collectively, our findings suggest that genetic variation plays a critical role in determining drought adaptive potential, even among ecotypes originating from adjacent subtropical regions [51,52].
3.2. Responses of Stomatal Conductance and Water Use Efficiency Response to Light Under Drought Stress
Stomata regulate gas exchange between plants and the atmosphere and are central to plant water and carbon balance [8]. Under drought conditions, reductions in stomatal conductance are often linked to declining hydraulic capacity [8], making stomatal regulation a key determinant of water use efficiency (WUE) and drought survival [20,22]. Understanding these responses is crucial for identifying the type of stomata that optimizes water use efficiency and photosynthetic performance under different environmental conditions [53]. However, stomatal responses to drought vary widely among species, functional types, and genotypes [54,55,56], and remain poorly understood in I. lanceolatum.
Previous studies have shown that stomatal conductance declines rapidly with drought in some species while remaining relatively stable in others [54]. Herbaceous plants generally exhibit steeper declines in g_s_ than woody species, with shrubs and lianas showing intermediate responses [54]. In woody plants, drought strategies range from stomatal closure to maintain hydraulic safety to sustained gas exchange under moderate water stress [22,55,56,57]. Integrating an empirical response into the g_s_ model based on experimental measurements was a very simple way to simulate the g_s_ response to drought [58,59].
In the present study, g_s_ responses to drought differed markedly among I. lanceolatum ecotypes. Stomatal conductance decreased under drought in the NP ecotype, consistent with conservative water use strategies reported in other species. In contrast, g_s_ increased in the WN, KH, and LA ecotypes during short-term drought and remained higher after rewatering than at the onset of the experiment. These contrasting patterns highlight pronounced ecotypic variation in stomatal regulation.
After rewatering, g_s_ in the WN and KH ecotypes exceeded values observed during drought, whereas g_s_ in the LA and NP ecotypes remained significantly lower than initial levels. These results suggest that drought-tolerant ecotypes maintain greater stomatal plasticity and recovery capacity. Interestingly, the initial low g_s_ values observed under well-watered conditions may have resulted from excessive irrigation, potentially causing transient root hypoxia. This finding underscores the importance of avoiding overwatering in cultivation practices for I. lanceolatum.
Under saturating light conditions (PAR > 1500 µmol·m^−^^2^·s^−^^1^), significant polynomial relationships were observed between soil water content and g_s_ in the WN, KH, and LA ecotypes, while the NP ecotype exhibited an exponential relationship. These nonlinear responses differ from the linear relationships reported in some previous studies [60], likely reflecting differences in genetic background, environmental context, and temporal scale of measurement.
Stomatal conductance plays a dual role in regulating photosynthesis and water use efficiency [61,62,63,64]. In this study, dynamic changes in g_s_ had ecotype-dependent effects on iWUE. Under short-term drought and high light conditions, increased iWUE was observed across ecotypes, particularly in the WN and KH ecotypes. These results indicate that these two ecotypes are better able to balance carbon gain and water loss under stressful conditions.
Overall, the WN ecotype exhibited superior performance across a wide range of light intensities and water conditions, reflecting broad ecological adaptability. The NP ecotype also showed relatively high tolerance, whereas the KH and LA ecotypes exhibited narrower soil moisture requirements. These findings have practical implications for urban forestry and conservation, where water limitation is increasingly common due to urban warming and climate change [65,66].
Based on our results, we recommend reducing light intensity through shading when soil water availability is limited, as this strategy can mitigate drought-induced stress and enhance iWUE in I. lanceolatum. This conclusion aligns with previous findings showing improved light use efficiency under shaded conditions in olive trees [45]. Ultimately, successful introduction and cultivation of I. lanceolatum across diverse environments should consider both genetic background and environmental interactions, with integrated water and light management strategies tailored to ecotype-specific responses.
4. Materials and Methods
4.1. Study Site
The experiment was carried out in Tian-mu Mountain Forest Nursery, Lin’an, Zhejiang province, China (118°51′–119°52′ E, 29°56′–30°23′ N). The elevation of the study site was 47 m asl. Central North Asia features a seasonal monsoon climate. The average annual temperature was 15.4 °C and the mean annual rainfall was 1600 mm in 2020. The experimental plot was set up on a flat land in front of a slope. The physical and chemical properties of the soil were determined synchronously in this study site before our experiment. Therefore, soils were acidic yellow loam with a pH value of 4.62. The chemical properties of the soils featured total organic matter content of 3.95 g·kg^−1^, total nitrogen of 0.44 g·kg^−1^, total phosphorus of 10.09 g·kg^−1^, total potassium of 6.31 g·kg^−1^, available nitrogen of 217.62 mg·kg^−1^, available phosphorus of 363.97 mg·kg^−1^, and available potassium of 40.51 mg·kg^−1^. Specific determination standards and test methods referred to forestry industry standards of the People’s Republic of China (LY/T 1228-2015, LY/T 1232-2015, LY/T 1234-2015, LY/T 1237-2015, and LY/T 1239-2015) [67,68,69,70,71], and all of the standards drafted by the Research Institute of Forestry, Chinese Academy of Forestry, and issued by the State Forestry and Grassland Administration. Among them, the determination of soil total nitrogen and available nitrogen is based on the standard of LY/T 1228-2015 [67], the determination of soil total phosphorus and available phosphorus is based on the standard of LY/T 1232-2015 [68], the determination of soil total potassium and available potassium is based on the standard of LY/T 1234-2015 [69], the determinations of soil featured total organic matter content is based on the standard of LY/T 1237-2015 [70], and the determination of pH value is based on the standard of LY/T 1239-2015 [71].
4.2. Method
4.2.1. Preparation of Experimental Plants
This study focused on four ecotypes from Lin’an Zhejiang (LA, 119°24′–119°28′ E, 30°18′–30°24′ N), Kai’hua Zhejiang (KH, 118°01′–118°37′ E, 28°54′–29°30′ N), Wu’ning Jiangxi (WN, 114°29′–114°36′ E, 28°53′–29°14′ N), and Nan’ping Fujian (NP, 117°10′–117°24′ E, 26°15′–27°19′ N), respectively. Seedlings were raised in an original location for each ecotype. Fruits were harvested from healthy wild plants in each location during September–October in 2020. Air-dried seeds were stored in moist sands at 4 °C. In mid-April 2021, seeds were sown in the field on a 10 m × 1.0 m seed bed with inter-row spacing of 10 cm and in-row spacing of 5 cm.
The growth medium was a mixture of garden soils and an organic fertilizer (2:1). During the seedling emergence period in early June 2021, the average planting density was 189 plants·m^−2^. This was the seeding emergence density. Later, as the seedlings grew, thinning was carried out to reduce the seedling density avoid shading. Following a routine practice, the field was mulched with straw immediately after seeding, which could provide 50% of shade during the hot summer season, which ranged from July to September in 2021. Irrigation was applied at the seed emergence stage, and seedlings were thinned to the same density at all four locations. One-year-old seedlings were then transferred to the location where this study was conducted. The natural geographical distribution regions and the four ecotypes in this experiment of I. lanceolatum are shown in Figure 8, and the climatic conditions of the four original distribution regions for the four ecotypes are provided in Table 2. There was a significant difference among the average annual rainfall for the four ecotypes in their natural growth environments. The annual average temperature of the natural growth environment of the NP ecotype was significantly different from those of the other three ecotypes (Table 2). The growth characters of 50 one-year-old seedlings for each I. lanceolatum ecotype are shown in Table 3.
4.2.2. Experimental Design
On 10 June 2022, 10 uniform plants of one-year-old seedlings were selected for each ecotype, and repotted into plastic pots 20 cm wide and 25 cm deep. To prevent the different root systems of the four ecotypes having varying effects on water absorption and utilization intensity, a single-plant-per-pot planting experimental method was adopted. Each pot was filled with 10 kg of a potting mix of sieved yellow soils (2 parts), sands soils (1 part), and humus (1 part). Soil bulk density (d) was measured, and then seedlings were transplanted into the pots. One-third of each pot was buried in the ground, and plants were watered regularly. In order to prevent the effect of rainfall on the water stress test, the experiment was conducted in the plastic awning, and the seedlings were given normal illumination in sunny days. All photosynthetic parameters were measured outside the shed.
The fast-growing season begins in mid-July. During the experiment, a soil water probe (ThetaProbe, ML2x, Delta-T Devices, Cambridge, UK) was used to monitor the soil volumetric water content (Wv). For each drought treatment, three replicates of measurements for Wv were taken, and the mean value was finally calculated and recorded. The soil mass water content (Wm) was estimated using the formula Wm (%) = Wv × 100%/d, where Wm and Wv are soil mass water content and soil volumetric water content, respectively, and d is the mean value of soil bulk density from all the pots. Wm at the four original seedling growing locations and during different periods of drought treatment is given in Table 1.
Pots were watered weekly. On 20 July 2022, after sufficient watering until it reached a constant weight, drought treatment was initiated, photosynthesis was measured on the same day, and data of pre-drought treatment was recorded. Subsequent measurements were taken at 2 d, 4 d, and 7 d after last well watering and during the recovery stage of rewatering. The measurement was performed on the second day after rewatering sufficiently until it reached a constant weight.
4.2.3. Photosynthesis and Changes in Light Response Parameters
Photosynthesis was measured using a portable photosynthesis system (Licor-6400, Li-Cor Inc., Lincoln, NE, USA) on the day of watering (control); 2 d, 4 d, 7 d after drought treatments; and during the recovery period after watering was resumed. Five plants from each ecotype were selected for fixed photosynthesis measurement, and two or three of the upper south facing canopy leaves were used randomly for the measurements with one reading of each leaf. During measurement of light responses of photosynthetic parameters, the red–blue light source was provided by the Licor-6400. Measurements were conducted at an air flow rate of 0.5 L·min^−1^, leaf temperature of 26 °C, relative humidity of 60%, and CO_2_ concentration of 400 µmol·mol^−1^. Each measurement followed an illumination gradient and started at 2000 µmol·m^−2^·s^−1^ of PAR, and gradually decreased to 1800, 1500, 1200, 1000, 800, 500, 200, 120, 80, 50, 20, and 0 µmol·m^−2^·s^−1^.
A light response curve was generated using data taken on sunny days from 9:00–11:00 a.m. A net photosynthetic rate (Pn)–photosynthetic active radiation (PAR) curve was fitted to a nonrectangle hyperbola model [72] to estimate photosynthetic parameter values [72]. The net photosynthetic rate (Pn), stomatal conductance (gₛ), and the transpiration rate (E) were determined using the Licor-6400 portable photosynthesis system (Licor-6400, Li-Cor Inc., Lincoln, NE, USA). At the leaf level, instantaneous water use efficiency (iWUE) was calculated by dividing the net photosynthetic rate by the transpiration rate (A/E) [36].
4.2.4. Data Analysis
Microsoft Excel 2003 (11.0, Microsoft Corporation, Redmond, WA, USA) and SPSS16.0 (SPSS Corporation, Chicago, IL, USA) were used to conduct the statistical analyses. Two-way analysis of variance (ANOVA) (p < 0.05) was used in the statistical software SPSS16.0 to compare the effect of the drought stress and duration time on the photosynthetic parameters according to different ecotypes. In this study, two-way analysis of variance showed that the differences among different drought treatments, among different ecotypes, and the interaction between drought and ecotype were all significant (p < 0.01). Subsequently, the least significant difference method (LSD) was used to perform post hoc comparisons to examine the significant differences of various photosynthetic physiological parameters among different drought treatments and among different provenances. The correlation between the gₛ and the iWUE was also analyzed for each ecotype according to different lengths in the drought period.
5. Conclusions
Under global climate change, drought has become an increasingly serious constraint on plant growth and productivity. This study demonstrates clear ecotypic differences in photosynthetic performance, gₛ, and iWUE in four ecotypes of Illicium lanceolatum in response to short-term drought stress.
Compared with well-watered conditions, photosynthetic capacity was moderately enhanced after 2–4 days without watering, accompanied by increased gₛ across ecotypes. This response likely reflects alleviation of root hypoxia caused by excessive watering prior to drought onset. Under saturating light intensity (>1500 μmol·m^−^^2^·s^−^^1^), significant polynomial relationships were observed between soil water content (W_m_) and gₛ in the WN, LA, and KH ecotypes, indicating strong stomatal sensitivity to soil moisture under high irradiance.
Following rewatering, g_s_ in the WN and KH ecotypes exceeded values observed during drought, whereas g_s_ in the LA and NP ecotypes remained significantly lower than those under initial well-watered conditions. Short-term soil drought combined with high light intensity promoted iWUE across ecotypes, with the WN ecotype exhibiting the widest ecological adaptability. This ecotype maintained strong stomatal regulation under severe drought, enabling relatively high photosynthetic potential and efficient use of light and water resources.
Overall, drought stress enhanced leaf sensitivity to high light, and both water limitation and light inhibition were the dominant factors driving photosynthetic decline among ecotypes. These findings highlight the importance of considering interactions between genetic background (ecotype) and environmental factors (water availability and light intensity) when introducing I. lanceolatum into new climatic regions. Optimized irrigation and shading strategies should, therefore, be integrated into management practices to achieve high-efficiency cultivation under increasingly water-limited conditions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Khan Z. Jan R. Asif S. Farooq M. Jang Y.H. Kim E.G. Kim N. Kim K.M. Exogenous melatonin induces salt and drought stress tolerance in rice by promoting plant growth and defense system Sci. Rep.202414121410.1038/s 41598-024-51369-038216610 PMC 10786868 · doi ↗ · pubmed ↗
- 2Ali S. Mir R.A. Haque M.A. Danishuddin Almalki M.A. Alfredan M. Khalifa A. Mahmoudi H. Shahid M. Tyagi A. Exploring physiological and molecular dynamics of drought stress responses in plants: Challenges and future directions Front. Plant Sci.202516156563510.3389/fpls.2025.156563540196426 PMC 11973087 · doi ↗ · pubmed ↗
- 3Guan X.Y. Jansen S. Huang L.X. Chen S.L. Zhu S.D. Evergreen species exhibit higher growth resistance under drought: Insights from carbon-water relations Tree Physiol.202545 tpaf 11510.1093/treephys/tpaf 11540981668 · doi ↗ · pubmed ↗
- 4Niemczyk M. Wrzesiński P. Wojda T. Mohytych V. High phenotypic variation in growth, stomatal regulation, and water-use efficiency in Quercus robur L. provenances may suffice for climate change adaptation For. Ecol. Manag.202559812324210.1016/j.foreco.2025.123242 · doi ↗
- 5Liu D. Wang Z.G. Han B.X. Mencuccini M. Camarero J.J. Xie Y.S. Zhao B.Q. Wang X.C. Plasticity of physiological, anatomical and structural traits defines seedling growth during sustained drought Tree Physiol.202545 tpaf 10310.1093/treephys/tpaf 10340844023 · doi ↗ · pubmed ↗
- 6Theroux Rancourt G. Ethier G. Pepin S. Greater efficiency of water use in poplar clones having a delayed response of mesophyll conductance to drought Tree Physiol.20153517218410.1093/treephys/tpv 00625721370 · doi ↗ · pubmed ↗
- 7Allen B.S. Stewart J.J. Polutchko S.K. Ocheltree T.W. Gleason S.M. Long-term in vivo observation of Maize leaf xylem embolism, transpiration and photosynthesis during drought and recovery Plant Cell Environ.2025484114412510.1111/pce.1541439901747 PMC 12050386 · doi ↗ · pubmed ↗
- 8Martin-St Paul N. Delzon S. Cochard H. Plant resistance to drought depends on timely stomatal closure Ecol. Lett.2017201437144710.1111/ele.1285128922708 · doi ↗ · pubmed ↗