Plant Molecular Phylogenetics and Evolutionary Genomics III
Aleksey V. Troitsky

Abstract
Click any figure to enlarge with its caption.
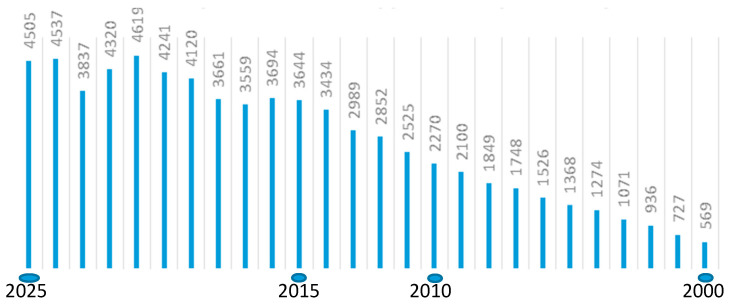 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Chromosomal and Genetic Variations · Genetic diversity and population structure
Comparative analyses of the structure of genes and genomes are an important tool in elucidating the phylogenetic relationships of organisms, the evolutionary pathways of genetic material, and its structural and functional basis. The evolutionary paradigm serves as a blueprint for investigations in almost all biological disciplines. Over time, the number of publications on this topic has only increased (Figure 1).
A PubMed search (https://pubmed.ncbi.nlm.nih.gov, accessed on 11 December 2025) identified more than 9000 articles on Plant Molecular Phylogenetics published in 2024–2025. The publications presented in this Special Issue represent a fairly representative cross-section of research in the topic.
The objects of analysis in about half of the articles were the genomes of chloroplasts (CP) from representatives of five families of angiosperms, as well as Sphenopsida. By comparing the CP genomes of 48 accessions of varieties and species of Zea, Montenegro et al. [1] established the polyphyletic origin of Peruvian purple maize in South America and the complex circumstances of its domestication with recurrent gene flow from wild relatives.
Lei et al. [2] reconstructed the evolutionary relationships of Kitagawia (Apiaceae) species, which are poorly understood, using CP genomes and nuclear rDNA sequences. The phylogenetic trees constructed from CDS of 27 CP genomes and nuclear ITS + ETS from the rDNA of Kitagawia are not completely congruent, as is commonly found in other studies, but indicate that all six examined Kitagawia species were divided into Subclade I and Subclade II within the tribe Selineae, and they were all distant from the representative members of Peucedanum sensu stricto. This study supported the separation of Kitagawia from Peucedanum sensu lato, confirming that Kitagawia belongs to Selineae, and that two species (K. praeruptora and K. formosana) within Subclade II should be placed in a new genus.
Goryunova et al. [3] sequenced the complete chloroplast genomes of Solanum tuberosum accessions for the first time, with five out of the six major cytoplasmic genome types. Based on the phylogenetic analysis of the complete plastome sequences, five main evolutionary branches of CP genomes (clades A–E) can be distinguished within the Petota section. Clade D comprises accessions with the M-, A, and P cytoplasm types. Samples with A- and P- cytoplasm formed isolated distant groups within a large and polymorphic group of samples with M-type cytoplasm, suggesting that the A and P genomes arose independently. Moreover, given the independent origin of the A- and P-type cytoplasmic groups, it seems unlikely that Andean cultivated tetraploids (S. tuberosum group Andigena) evolved directly from early landrace diploids (S. tuberosum groups Stenotomum and Phureja) through autopolyploidy. Accessions with the sterilizing cytoplasm types W, D, and T are part of the E clade. The divergence time between the D and E clades can be estimated as 4.71 MYA (3.78–5.70). The M type of cytoplasm appears to be the most ancient. The findings suggest that the diversity of the T-genome in S. tuberosum Group Tuberosum could be initially low due to the bottleneck already occurring at the origin of the Chilean clade. Revealed variations in the rbcL gene sequence may be one of the factors causing differences in the appearance of economically important traits between species with A and T-type cytoplasm.
Comparative analyses of 45 Plantago species (Plantaginaceae) based on fully sequenced plastomes identified the most variable mutational hotspots that were not suitable for the development of species-specific molecular markers; however, species-specific polymorphisms could discriminate P. lanceolata from its closest relatives [4]. Molecular evolutionary analyses indicated that eleven protein-coding genes involved in different functions in Plantago plastomes underwent positive selection, suggesting they might have contributed to enhancing species’ adaptation during the evolutionary history of Plantago. It is possible that the complex climatic oscillations that took place in the late Miocene and the beginning of the Pliocene could have at least in part contributed to determining the selective patterns observed in rpoB and rpoC2 genes of the studied Plantago.
Samigullin et al. [5] sequenced plastomes of 16 species from the tribe Loteae (Fabaceae) and reconstructed the phylogeny of this tribe basing on 23 complete CP genomes. Comparative analysis revealed several regions (petN-trnC and rps16-accD spacers from the LSC region of CP DNA, and in the ycf 1 gene within the SSC) as the most variable and potentially useful for phylogenetic purposes.
For horsetails (Sphenopsida), Satjara et al. sequenced the complete CP genome of Equisetum xylochaetum [6] but used only five markers (atpB, matK, rpoB, rps4, and trnL-F) to reconstruct the phylogeny of four species. In addition to this, the TCS haplotype networks of atpB, matK, rpoB, rps4, and trnL-F were calculated for 105 Equisetum accessions. Some Equisetum species were categorized as more than a single haplotype.
Of the nuclear genes, rDNA is the most frequently used in molecular phylogenetics. Internal transcribed spacers (ITS) of nuclear rDNA were used as barcodes for the identification of medicinal plants from the Tianshan wild fruit forest region [7] and to study the intragenomic polymorphism of cultivated and wild Avena species [8]. According to the data obtained, the diploid A. strigosa could have evolved independently of the polyploid-cultivated species, and the tetraploid A. abyssinica could be a cultivated derivative of A. vaviloviana and hexaploid-cultivated species, A. byzantina and A. sativa, which could have a different origin. A. sativa could be the cultivated form of A. fatua, whereas A. byzantina could originate independently.
Using RAD-seq technology, 2996 high-quality single-nucleotide polymorphisms (SNPs) were identified in Stewartia gemmata and S. acutisepala (Theaceae), the analysis of which was used to infer the evolutionary relationships of two species [9]. The results suggested strong monophyly of both species, and S. acutisepala was nested within S. gemmate.
Structure, evolution, and expression of several gene families were studied. The objects of the research were the lipoxygenase gene families (LOX) in angiosperms [10], the aquaporin (AQP) family in papaya Carica papaya (Brassicales) [11], the MYB gene family in Chrysanthemum X morifolium (Asteraceae) [12], and TCP Interactor containing EAR motif proteins genes (TIE) in Embryophyta [13]. Evolutionary trees were constructed for 247 LOX genes from 23 species of angiosperms and basal plants, 550 AQP genes from seven species, 1945 MYB genes in six plant species, and 102 TIE proteins from 33 embryophytes. Henning et al. [14], analyzing the newly sequenced and annotated draft genome of the Turnera subulata (Passifloraceae), identified in this plant 169 candidate genes of the three families BAHD, SPH, and YUC and reconstructed evolutionary trees for 1741 S-families members from 57 embryophytes.
Two articles are reviews. The first is devoted to the role of transposons in the evolution of polyploid plant genomes [15]. Plant transposons are the most variable part of the genome, causing extensive changes in the overall structure and gene function of the genome, affecting the genome regulatory network and evolution. Transposons and polyploidy are interconnected because polyploidy can induce transposon activity, and transposons enable polyploidy to produce new variants. The authors formulate key questions about the role of transposons in polyploid plant genome evolution and their regulatory mechanisms, which deserve further study. The second review [16] summarizes current knowledge on the biodiversity, taxonomy, and physiology of the genus Ulva and assesses the applications of these algae in food, feed, bioremediation, biofuels, pharmaceuticals, and biomaterials.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Montenegro J.D. Julca I. Chumbe-Nolasco L.D. Rodríguez-Pérez L.M. Sevilla Panizo R. Medina-Hoyos A. Gutiérrez-Reynoso D.L. Guerrero-Abad J.C. Amasifuen Guerra C.A. García-Serquén A.L. Phylogenomic Analysis of the Plastid Genome of the Peruvian Purple Maize Zea mays subsp. mays cv. ‘INIA 601’Plants 202211272710.3390/plants 1120272736297753 PMC 9612013 · doi ↗ · pubmed ↗
- 2Lei J.-Q. Liu C.-K. Cai J. Price M. Zhou S.-D. He X.-J. Evidence from Phylogenomics and Morphology Provide Insights into the Phylogeny, Plastome Evolution, and Taxonomy of Kitagawia Plants 202211327510.3390/plants 1123327536501315 PMC 9740501 · doi ↗ · pubmed ↗
- 3Goryunova S. Sivolapova A. Polivanova O. Sotnikova E. Meleshin A. Gaitova N. Egorova A. Semenov A. Gins E. Koroleva A. Comparative Study of Plastomes in Solanum tuberosum with Different Cytoplasm Types Plants 202312399510.3390/plants 1223399538068632 PMC 10708428 · doi ↗ · pubmed ↗
- 4Mehmood F. Li M. Bertolli A. Prosser F. Varotto C. Comparative Plastomics of Plantains (Plantago, Plantaginaceae) as a Tool for the Development of Species-Specific DNA Barcodes Plants 202413269110.3390/plants 1319269139409561 PMC 11478842 · doi ↗ · pubmed ↗
- 5Samigullin T.H. Kopylov-Guskov Y.O. Nikitina O.V. Krinitsina A.A. Polevova S.V. Kramina T.E. Sixteen New Complete Plastid Genomes in the Tribe Loteae (Leguminosae): Structure and Phylogenetic Analysis Plants 20251461810.3390/plants 1404061840006877 PMC 11859275 · doi ↗ · pubmed ↗
- 6Satjarak A. Graham L.E. Trest M.T. Arancibia-Avila P. Plastid Genome of Equisetum xylochaetum from the Atacama Desert, Chile and the Relationships of Equisetum Based on Frequently Used Plastid Genes and Network Analysis Plants 202211100110.3390/plants 1107100135406981 PMC 9002811 · doi ↗ · pubmed ↗
- 7Jiamahate A. Bozorov T.A. Wang J. Zhang J. Zhang H. Wang X. Yang H. Zhang D. Insights from DNA Barcodes-Based Phylogenetic Analysis of Medicinal Plants and Estimation of Their Conservation Status: A Case Study in the Tianshan Wild Forest, China Plants 2025149910.3390/plants 1401009939795359 PMC 11723300 · doi ↗ · pubmed ↗
- 8Gnutikov A.A. Nosov N.N. Loskutov I.G. Rodionov A.V. Shneyer V.S. Participation of Wild Species Genus Avena L. (Poaceae) of Different Ploidy in the Origin of Cultivated Species According to Data on Intragenomic Polymorphism of the ITS 1-5.8S r RNA Region Plants 202514155010.3390/plants 1410155040431114 PMC 12114916 · doi ↗ · pubmed ↗