Natural Alkaloids as Antiviral Agents Against RNA Viruses: A Comprehensive and Mechanistic Review
Kristi Leka, Lúcia Mamede, Elyn Vandeberg, Mutien-Marie Garigliany, Allison Ledoux

TL;DR
This paper reviews how natural alkaloids can act as antiviral agents against RNA viruses like influenza and coronaviruses by targeting various stages of the viral life cycle.
Contribution
The paper provides a comprehensive and mechanistic review of alkaloids with antiviral activity against RNA viruses, distinguishing in vitro, in vivo, and in silico evidence.
Findings
Alkaloids interfere with multiple stages of RNA virus life cycles, including entry, replication, and immune modulation.
Quaternary alkaloids have unique interactions due to their ionic charge, affecting membranes and host pathways.
Clinical translation of alkaloids is limited by incomplete mechanistic validation and poor bioavailability.
Abstract
RNA viruses pose a persistent global threat due to their high mutation rates, zoonotic potential, and rapid adaptability. Emergence events have risen steadily, as demonstrated by major outbreaks caused by Influenza A, Ebola, Zika, and Chikungunya viruses, followed by the coronavirus epidemics of Severe Acute Respiratory Syndrome coronavirus (SARS-CoV-1) and Middle East Respiratory Syndrome Coronavirus (MERS-CoV) and culminating in the COVID-19 pandemic. These characteristics frequently compromise the durability of existing vaccines and antiviral therapies, highlighting the urgent need for new antiviral agents. Alkaloids, a structurally diverse class of nitrogen-containing natural compounds, have gained attention for their ability to interfere with multiple stages of the viral life cycle, including entry, replication, protein synthesis, and host immune modulation. To our knowledge, this…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
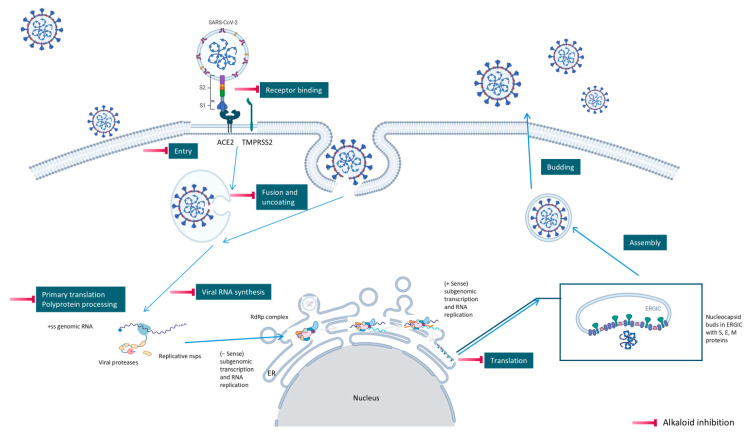 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12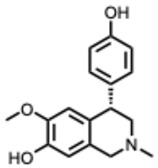 Figure 13
Figure 13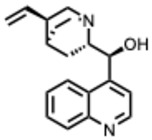 Figure 14
Figure 14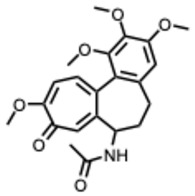 Figure 15
Figure 15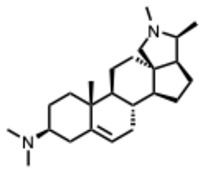 Figure 16
Figure 16 Figure 17
Figure 17 Figure 18
Figure 18 Figure 19
Figure 19 Figure 20
Figure 20 Figure 21
Figure 21 Figure 22
Figure 22 Figure 23
Figure 23 Figure 24
Figure 24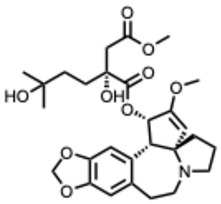 Figure 25
Figure 25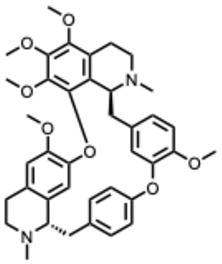 Figure 26
Figure 26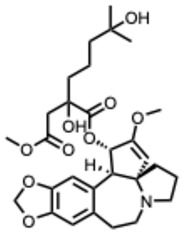 Figure 27
Figure 27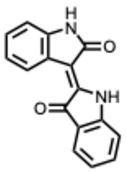 Figure 28
Figure 28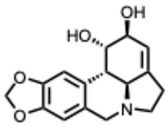 Figure 29
Figure 29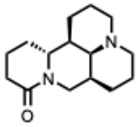 Figure 30
Figure 30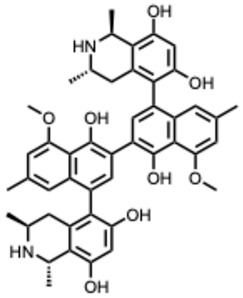 Figure 31
Figure 31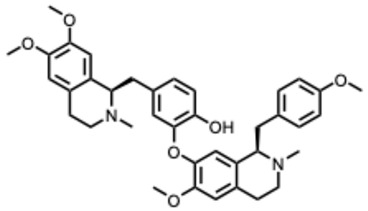 Figure 32
Figure 32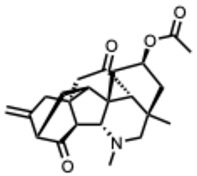 Figure 33
Figure 33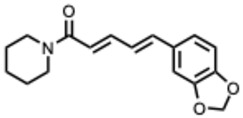 Figure 34
Figure 34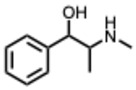 Figure 35
Figure 35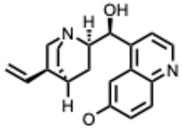 Figure 36
Figure 36 Figure 37
Figure 37 Figure 38
Figure 38 Figure 39
Figure 39 Figure 40
Figure 40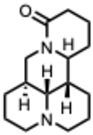 Figure 41
Figure 41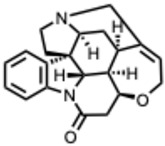 Figure 42
Figure 42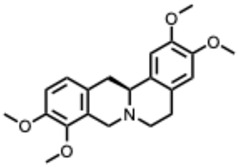 Figure 43
Figure 43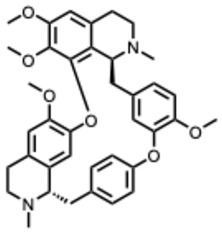 Figure 44
Figure 44 Figure 45
Figure 45 Figure 46
Figure 46 Figure 47
Figure 47 Figure 48
Figure 48 Figure 49
Figure 49 Figure 50
Figure 50| Compound & Origin | References | Study Type | Virus | Mechanisms | Antiviral Information Reported in the Original Studies (IC50, EC50, SI, Docking Scores, Qualitative Observations) * |
|---|---|---|---|---|---|
| Wu et al. 2011 [ | In vitro and in vivo | IAV | Berberine exerted strong inhibition on the inflammatory | IC50 = 0.025 g/L | |
| Wang et al. 2018 [ | In vitro | EV71 | Might inhibit MEK/ERK, | IC50: 7.12–14.8 | |
| Shao et al. 2020 [ | In vitro and in silico | HIV-1, clade B | It binds in the pocket of NHR and CHR of gp41. | IC50: 5.5–10.25 | |
| Ratanakomol et al. 2021 [ | In vitro | DENV, | Potential AMPK activation, | IC50: | |
| Botwina et al. 2020 [ | In vitro | IAV (H3N2) | Inhibits MAPK/ERK | IC50: | |
| Enkhtaivan et al. 2017 [ | In vitro and in silico | IAV | Competitive | Berberine derivatives IC50 | |
| Varghese et al. 2016 [ | In vitro | CHIKV | Might be affecting one or several host factors important for CHIKV replication | IC50: 1.9 ± 0.9 μM | |
| Nguyen, C. Q. et al. (2021) [ | In vitro and in silico | ZIKV | Potential candidate to inhibit NS2B-NS3 | Compound | |
| Españo, E. et al. (2022) [ | In vitro, in silico | ZIKV | Potential entry/attachment | EC50 = 692.4 nM | |
| Loe, M. et al. 2023 [ | In vitro, in vivo | ZIKV | Potent inhibitor of ZIKV infection that targets the ZIKV NS4B protein | Chelerythrine chloride | |
| Guo, W. et al. 2020 [ | In vitro ( | Tobacco Mosaic | Inactivation/proliferation | Chelerythrine at | |
| Orhan, I. et al. 2007 [ | In vitro | Parainfluenza-3 | N/A | CPE inhibitory concentration 16 µg/mL (≈ 40–44 µM ***) | |
| Dai, J. et al. 2018 [ | In vitro and in vivo | IAV multiple strains | Could significantly decrease the promoter activity of TLR signaling pathways | EC50: 5.91–23.67 | |
| Zhi et al. 2024 [ | In vitro | H9N2 Avian | TLR signaling pathways TLR3, NF-κB, IRF-3 | Dose-dependent | |
| Chen, N. et al. (2016) [ | In vitro | HCV | Proliferation inhibition | Inhibition of cell | |
| Majnooni, M. et al. (2021) [ |
| SARS-CoV-2 | Nucleotide biosynthesis inhibitor | EC50 = 0.31 μM, | |
| W. Zhang, 2024 [ | In vitro | Infectious Bronchitis virus | Inactivating the virus, inhibiting its replication, modulating NF-κB/IRF7/JAK-STAT signaling pathways, and regulating apoptosis | IC50 = 7.76 µM | |
| Fan Jia et al. 2010 [ | In vitro | WNV | Palmatine could significantly inhibit the activity of | WNV: | |
| Yi-Jung Ho et al. 2019 [ | In vitro | ZIKV | Inhibits Zika virus infection by disrupting virus binding, entry, and stability | Inhibited ZIKV binding by | |
| Qiyun Ke et al. 2023 [ | In vitro and in silico | PRRSV | Targeting internalization, replication, and release stages of the viral life cycle | Sanguinarine inhibits the propagation of PRRSV in a dose-dependent manner |
- —National Fund for Scientific Research (FNRS)
- —Foundation Léon Frédéricq
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlkaloids: synthesis and pharmacology · Chemical synthesis and alkaloids · Berberine and alkaloids research
1. Introduction
The objective of this review is to comprehensively compile naturally occurring alkaloids reported to exhibit antiviral activity against RNA viruses, a group characterized by frequent emergence, high mutation rates, and substantial pandemic potential. Beyond cataloging these compounds, the review critically examines their proposed mechanisms of action in relation to the experimental approaches used to characterize antiviral activity. Because the strength and translational relevance of reported findings depend strongly on study design, it is essential to distinguish between predictive computational analyses, cell-based assays, and animal models. While in silico screening and early in vitro studies provide valuable insights for compound prioritization, mechanistic validation and in vivo evaluation are required to substantiate therapeutic potential. Accordingly, this review explicitly differentiates alkaloids supported by in silico, in vitro, and in vivo evidence and discusses computationally identified candidates in a dedicated section, with emphasis on their limitations and translational relevance.
To support this analysis, a comprehensive literature search was conducted between August 2025 and December 2025 to identify peer-reviewed studies reporting antiviral activity of natural alkaloids against RNA viruses. The search strategy combined broad academic literature databases and targeted keyword-based queries to ensure extensive coverage across virus families and experimental contexts. Search terms included combinations of “alkaloids”, “quaternary alkaloids”, “antiviral”, and “RNA viruses”, together with the names of individual RNA viruses searched separately, including SARS-CoV-1, SARS-CoV-2, MERS-CoV, influenza A virus (IAV), dengue virus (DENV), Zika virus (ZIKV), Ebola virus (EBOV), West Nile virus (WNV), Japanese encephalitis virus (JEV), chikungunya virus (CHIKV), respiratory syncytial virus (RSV), hepatitis C virus (HCV), foot-and-mouth disease virus (FMDV), and porcine epidemic diarrhea virus (PEDV). Searches were performed iteratively to capture studies spanning in silico, in vitro, and in vivo experimental approaches.
Only peer-reviewed articles published in English were considered. Studies were included if they reported antiviral activity, target engagement, or mechanistic effects of natural alkaloids against RNA viruses based on primary computational, cellular, or animal data. Reviews, editorials, patents, and studies lacking primary experimental or computational evidence were excluded, except when cited to provide contextual background. Data extraction focused on alkaloid identity, viral target or virus family, experimental model, and reported antiviral outcomes, with emphasis on quantitative parameters when available.
In contrast to previous reviews focusing primarily on compound listings or single virus families, the present review emphasizes mechanistic patterns across alkaloid classes, distinguishes classical from quaternary alkaloids, and critically evaluates antiviral evidence according to experimental level (in silico, in vitro, in vivo). Particular attention is given to translational bottlenecks, including toxicity, pharmacokinetics, and formulation challenges.
Overview of RNA Viruses and Their Global Health Impact
Viruses are microscopic infectious agents incapable of independent replication, as they lack the cellular machinery required for reproduction [1]. To propagate, they must invade a host cell and exploit its biosynthetic systems for their own replication. In 1971, David Baltimore introduced a classification system for viruses based on the type and replication strategy of their genetic material [2]. Within this framework, RNA viruses are distinguished by their ribonucleic acid (RNA) genomes, which confer unique structural and functional characteristics distinct from those of desoxyribonucleic acid (DNA) viruses.
A defining feature of RNA viruses is their exceptionally high mutation rate, primarily attributed to the low fidelity of RNA-dependent RNA polymerases, the enzymes responsible for genome replication [3]. This intrinsic genetic instability enables RNA viruses to adapt rapidly to new hosts and environmental pressures, often resulting in immune evasion and the reduced efficacy of antiviral drugs.
Many human RNA viruses are zoonotic in origin, arising in animal reservoirs before crossing the barrier into human populations. The likelihood of such “spillover” events is increasing due to anthropogenic factors such as deforestation, urbanization, climate change, and intensified human–wildlife interactions [4]. Notable zoonotic RNA viruses include the Human Immunodeficiency Virus (HIV), Influenza A virus subtypes H1N1 and H5N1, Severe Acute Respiratory Syndrome coronavirus (SARS-CoV), Middle East Respiratory Syndrome coronavirus (MERS-CoV), and Severe Acute Respiratory Syndrome coronavirus 2 (SARS-CoV-2).
The COVID-19 pandemic, caused by SARS-CoV-2 virus, underscored not only the profound global health threat posed by RNA viruses but also the degree to which the world remains unprepared for such challenges. The pandemic demonstrated how rapidly RNA viruses can evolve and disseminate across populations by producing numerous variants within a remarkably short period. Each of the variants was characterized by distinct levels of transmissibility, virulence, and clinical presentation [5,6,7,8,9]. The emergence and global spread of these variants placed extraordinary long-term strain on healthcare systems and triggered extensive social, economic and public health disruptions.
When a novel RNA virus emerges in the human population, pre-existing immunity is typically absent, rendering communities highly susceptible to infection. The continuous co-evolution of RNA viruses and their hosts further complicates outbreak prediction and as such it delays the development of effective vaccines and therapeutic solutions [10]. In summary the high mutability, zoonotic potential, and rapid adaptability of RNA viruses underscore their significance as persistent and evolving threats to global health.
2. Virus Classification in the Genomic Era
The classification of viruses is a dynamic and continuously evolving process, shaped largely by advances in genomic sequencing technologies. Historically, viruses were categorized by phenotypic traits, host range, and associated diseases [11]. Today, the International Committee on Taxonomy of Viruses (ICTV) serves as the authoritative body for viral taxonomy [12]. Modern systems rely heavily on phylogenetics and genomic data, frequently structured within the Baltimore classification framework [11].
Metagenomic expansion has produced large datasets that challenge existing taxonomic frameworks, especially given recombination and reassortment [12]. Integrative approaches combining sequence identity analyses with phylogenetic methods, for example in Betaflexiviridae as described by Silva et al. (2022), have been proposed [13]. These methods aim to refine viral classification to better account for the diversity revealed by high-throughput sequencing [13].
Despite advances, the Baltimore system remains a pillar in organizing viruses by genome type and replication mode although minor revisions have reflected newly identified behaviors and genome structures [12].
Baltimore divides the viruses into the following groups:
- I: dsDNA (e.g., Herpes simplex virus)
- II: ssDNA → dsDNA before transcription (e.g., Parvovirus)
- III: dsRNA; mRNA transcribed from RNA genome (e.g., Rotavirus)
- IV: (+)ssRNA; genome functions as mRNA (e.g., Picornavirus, Coronavirus, Flavivirus)
- V: (−)ssRNA; mRNA transcribed from RNA genome (e.g., Rabies virus, Influenza virus, Ebola virus)
- VI: ssRNA with reverse transcriptase (e.g., HIV)
- VII: dsDNA with reverse transcriptase (e.g., Hepatitis B virus)
This review aims its focus on Groups III, IV, V, and VI concerning only the RNA viruses responsible for major outbreaks in recent decades, including Influenza A virus, MERS-CoV, SARS-CoV, HIV, Ebola virus, Zika virus, Japanese Encephalitis virus and SARS-CoV-2.
3. Alkaloid Biosynthetic Pathways and Structural Diversity
Plant alkaloids are structurally diverse nitrogen-containing compounds with broad biological activity and long medicinal use. Their complexity arises from enzyme-mediated modifications, glycosylation, acylation, reduction, oxidation, and methylation, which diversify structures and modulate bioactivity [14]. Cytochrome P450–mediated oxidations are particularly pivotal, enabling ring formation, cleavage, and expansion that greatly increase molecular diversity [15].
Advances in molecular biology and biotechnology have illuminated genes involved in alkaloid biosynthesis. For example, Kishimoto et al. 2016 used synthetic biology and microbial systems (Escherichia coli, Saccharomyces cerevisiae) to verify gene function, optimize production, and generate structural analogs, facilitating scalable pharmaceutical applications [15,16].
Obtaining useful amounts of alkaloids from plants often requires large quantities of raw material, considerable time, and specialized equipment to extract and identify each compound. Because of these demands, traditional isolation can be costly and environmentally taxing. Biosynthetic approaches, on the other hand, offer a more sustainable and practical alternative, allowing alkaloids to be produced without relying heavily on plant harvesting.
3.1. Classification of Alkaloids
Alkaloids are commonly classified by biosynthetic origin, emphasizing the presence and source of the nitrogen atom. Protoalkaloids are defined by the nitrogen outside the heterocyclic ring. On the contrary, for the true alkaloids the nitrogen is incorporated within a heterocyclic ring. Whereas for the pseudoalkaloids, the nitrogen is derived from non–amino acid precursors [17].
A structural classification based on the ring systems is also available into the following groups: quinoline, isoquinoline, pyrrole, pyrrolidine, pyridine, piperidine, pyrrolizidine, and indole-containing alkaloids [18].
3.2. Host Hijacking and Viral Translation
RNA viruses have evolved strategies to hijack host machinery for replication and protein synthesis. By manipulating ribosomes, they redirect translation toward viral proteins, often at the expense of host antiviral factors [19]. Some cleave key host factors or modulate PRR (pattern recognition receptor) activation, evading immune detection and suppression [20,21].
Because many RNA viruses lack a canonical 5′ cap and differ in other structural features, they employ non-canonical translation mechanisms to initiate protein synthesis, including internal ribosome entry, leaky scanning, and ribosomal frameshifting. Viral RNAs can also form circular structures that, similar to cellular mRNAs, recruit host initiation factors to drive protein production [22].
A major challenge for RNA viruses is coordinating translation and replication, which compete for shared host factors. To ensure efficient infection, viruses must precisely regulate these processes, maintaining a balance that allows genome replication to proceed while still producing the proteins required for virion assembly.
3.3. Alkaloid Antiviral Modes of Action
Alkaloids exhibit antiviral activity against RNA viruses and, in some cases, DNA viruses, demonstrating effects at multiple stages of the viral life cycle [23,24]. Their antiviral mechanisms include inhibition of viral entry, either by interfering with receptor binding or by inducing membrane perturbation. They may also suppress viral replication through direct inhibition of essential enzymes such as RNA-dependent RNA polymerase (RdRp) or viral proteases. Additionally, several alkaloids disrupt viral protein synthesis, further limiting viral proliferation.
Beyond direct antiviral effects, alkaloids can exert indirect activity by modulating immune responses, including the regulation of cytokines, reactive oxygen species (ROS), and interferons [25].
Examples that illustrate these mechanisms include 10-hydroxyusambarensine and cryptospirolepine, which show strong binding affinity in silico for multiple viral targets [26,27]. Isoquinoline alkaloids modulate key inflammatory pathways such as NF-κB and MAPK/ERK [28]. Meanwhile, compounds such as tetrandrine, oxymatrine, and berberine demonstrate potent immunomodulatory activity by suppressing excessive inflammation [29].
During the course of this review, clear distinctions emerged between the antiviral activities of general alkaloids and those of their specialized subclass, the quaternary alkaloids. These differences were not limited to their molecular structures, specifically the permanently charged quaternary ammonium group, but were also evident at the pharmacological level.
Because quaternary alkaloids carry a permanent positive charge, their limited membrane permeability often restricts their antiviral activity to early steps of the viral life cycle, such as viral attachment or entry. They often act through mechanisms such as membrane disruption, interference with viral entry, or modulation of lysosomal and host signaling pathways. In contrast, many non-quaternary alkaloids display activity across a broader range of viral processes, including replication, protein synthesis, and immune modulation.
Differences were also observed in potency, cytotoxicity, selectivity index, bioavailability, and overall potential for clinical translation. Given these mechanistic and pharmacological distinctions, we determined that it was more appropriate to present alkaloids and quaternary alkaloids in two separate tables (Table 1 and Table 2) accordingly to ensure clarity and scientific accuracy.
In this review, antiviral evidence is discussed according to the experimental level at which it was generated, namely in silico, in vitro, and in vivo. While computational studies provide valuable hypotheses regarding potential targets and binding modes, they do not constitute proof of antiviral efficacy and must be interpreted with caution. In vitro and in vivo studies are therefore discussed separately where possible to reflect their respective levels of biological validation.
Figure 1 presents SARS-CoV-2, the most recent coronavirus responsible for a global pandemic, as an example to illustrate the viral replication cycle within the host and the key steps at which a known alkaloid can exert inhibitory activity.
4. Quaternary Alkaloids: Distinct Mechanisms and Pharmacology
Quaternary alkaloids constitute a distinct subclass of alkaloids defined by a permanently charged quaternary ammonium group. Because of this ionic structure, they exhibit physiochemical and pharmacological properties distinguishing them from other alkaloid types.
Quaternary alkaloids can influence several stages of the viral replication cycle, including entry, replication, protein synthesis, and modulation of host responses, similar to alkaloids in general. However, because of their permanent positive charge, these compounds have low membrane permeability and limited intracellular diffusion. As a result, their antiviral activity is often strongest during the early steps of infection, particularly at the level of viral attachment or entry. For example, tetrandrine, a known quaternary alkaloid, blocks viral entry by targeting NPC1, causing lysosomal cholesterol accumulation and triggering interferon responses via NPC1–STING [110].
Unfortunately, quaternary alkaloids display poor bioavailability, rapid clearance, and short half-lives [111]. However, emerging delivery technologies such as liposomes, nanoparticles, transdermal systems can improve absorption, extend circulation, and enhance pharmacological performance [111,112,113]. Another important issue with the quaternary alkaloids is that the activity often occurs near the cytotoxic thresholds [114]. The following (Table 2) summarizes the key quaternary alkaloids studied to date and their antiviral activities across RNA viruses.
To improve clarity and methodological transparency, we compiled an exhaustive transversal overview summarizing the level of experimental evidence supporting the antiviral activity of each alkaloid discussed in this review. Alkaloids are traditionally presented alphabetically to facilitate compound identification; however, this organization may obscure differences in experimental validation levels. The present table (Table 3) addresses this limitation by distinguishing in silico, in vitro, and in vivo evidence without duplicating compound-specific antiviral data already detailed in Table 1 and Table 2. Missing information is indicated by “–”, reflecting the absence of reported data rather than negative results. This overview highlights the current imbalance between extensive computational or cell-based screening and the limited number of compounds validated in vivo, underscoring key translational gaps in alkaloid-based antiviral research.
5. Discussion
Across both datasets, a coherent picture emerges, alkaloids represent a structurally diverse and mechanistically rich source of antiviral agents, with activity documented against a broad range of RNA viruses. Throughout this review, antiviral evidence is discussed according to the experimental level at which it was generated, namely in silico, in vitro, and in vivo, to reflect the degree of biological validation associated with each approach. In the first dataset, 42 compounds supported by 77 experimental entries show that approximately 45 percent of studies were performed exclusively in vitro, around 30 percent combined in vitro and in vivo approaches, and 25 percent relied mainly on in silico predictions. While computational studies provide valuable hypotheses and help prioritize candidate scaffolds, they do not constitute proof of antiviral efficacy and therefore require careful interpretation in the absence of experimental validation. This imbalance highlights extensive early screening but a shortage of deeper, mechanistic, and translational work. Another major bottleneck often occurs at the “in vitro–in vivo” transition, where a majority of compounds showing antiviral activity in vitro ultimately fail to demonstrate efficacy in vivo [23].
A central theme is the convergence of many alkaloids on viral replication machinery, particularly the RdRp. Compounds such as tetrahydroisoquinoline derivatives, lycorine, and aloperine consistently interact with RdRp or polymerase-associated subunits, often at low micromolar or nanomolar levels. Their rigid, polycyclic scaffolds appear intrinsically suited to binding nucleic acid–proximal enzymatic pockets, which may underlie their broad-spectrum effectiveness across coronaviruses, orthoflaviviruses, alphaviruses, and orthomyxoviruses.
A second mechanistic cluster involves entry inhibition. Neferine, fangchinoline, rutaecarpine, berbamine and capsaicin interfere with viral attachment, membrane fusion, or endosomal trafficking. These activities correlate with amphiphilic or cationic structural elements capable of perturbing lipid or acidic intracellular environments. Because many of these effects are host-directed, resistance may emerge more slowly, though this benefit must be balanced against increased toxicity risks.
Host-directed immunomodulation represents a third recurring pattern. Indirubin, emetine, ephedrine, and canthin-6-one derivatives modulate key pathways including NFκB, JAK, STAT, TLR, MAPK, and ERK. This dual antiviral and anti-inflammatory activity may be especially valuable in infections characterized by immunopathology such as influenza and SARS-CoV-2. At the same time, manipulating host signaling requires careful dosing and safety evaluation.
The second dataset, focused on berberine, chelerythrine, dehydrocorydaline, oxymatrine, palmatine, and sanguinarine, reveals similar mechanistic tendencies but with a clearer emphasis on innate immunity, inflammatory regulation, and endolysosomal physiology. Study distribution again skews toward in vitro work (≈50 percent), though about 35 percent include in vivo components. Berberine, palmatine, and oxymatrine frequently modulate TLR3, TLR4, NFκB, IRF3, AMPK, and autophagy-related pathways, producing broad-spectrum antiviral effects across influenza viruses, enteroviruses, flaviviruses, chikungunya virus, hepatitis C virus, and SARS-CoV-2. Meanwhile, quaternary or quaternary-like alkaloids such as chelerythrine and sanguinarine are particularly effective at disrupting viral entry and endosomal maturation through their permanent or semi-permanent cationic charge.
Cepharanthine (CEP) is one of the few alkaloids near clinical translation. Although an enhanced oral formulation (PD 001) received FDA clearance for Phase I and II testing in mild to moderate COVID 19, no PD 001 trials have begun, and the only active study is a Phase II placebo-controlled trial in mild COVID 19 (NCT05398705). Because intravenous CEP is impractical and standard oral dosing fails to reach antiviral levels, animal pharmacokinetic data indicating over 64 percent bioavailability with pulmonary delivery highlight inhaled formulations as the most promising route for clinical development [45].
Despite these strengths, significant limitations remain. Many alkaloids show narrow therapeutic windows, with active concentrations approaching cytotoxic levels. Depending on their chemical structure and administered dose, alkaloids may induce neurotoxicity, hepatotoxicity, nephrotoxicity, or cardiotoxicity [134]. Moreover, species-specific differences in metabolism and organ sensitivity further complicate the translation of preclinical findings to humans. A well-documented example is the class of pyrrolizidine alkaloids, which are strongly associated with hepatotoxic and carcinogenic effects due to their metabolic conversion into reactive intermediates [135]. Toxicity assessment is often limited to single- dose MTT assay (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay) which measures cell metabolic activity or LDH assays (Lactate Dehydrogenase release assay) which measure cell membrane integrity in immortalized cell lines. However, these assays cannot predict organ-specific or long-term effects. Pharmacokinetic challenges such as poor oral bioavailability, limited solubility, extensive first-pass metabolism and short half-lives, further restrict their translational potential [136]. These issues highlight the need for optimized formulations, including nanoparticles, liposomes, prodrugs, and targeted delivery systems.
Beyond conventional molecular docking and virtual screening approaches, artificial intelligence (AI) and machine learning (ML)-based methods are increasingly being explored in antiviral drug discovery to prioritize natural compounds, including alkaloids. These approaches differ fundamentally from classical docking by integrating large multidimensional datasets to identify non-obvious structure–activity relationships through quantitative structure–activity relationship (QSAR) models, supervised learning algorithms, or deep neural networks. In principle, AI-driven screening may enable rapid prioritization of alkaloids with multi-target potential, optimization of scaffold–activity relationships, and early prediction of pharmacokinetic and ADMET properties. However, despite their growing visibility, the application of AI-based models to alkaloid antivirals against RNA viruses remains largely prospective. Model performance is strongly dependent on the quality, size, and chemical diversity of training datasets, which are often biased toward synthetic compounds and well-characterized viral targets. Moreover, AI predictions require experimental validation, as computational confidence does not necessarily translate into antiviral efficacy or acceptable toxicity profiles. At present, AI-based approaches should therefore be viewed as complementary hypothesis-generating tools rather than substitutes for experimental screening, and their integration into alkaloid-based antiviral research will depend on the generation of standardized, high-quality biological datasets.
In addition, natural source diversity remains underexplored. While terrestrial plants dominate the current research landscape, marine organisms and endophytic fungi represent rich reservoirs of chemically unique alkaloids featuring halogenation, sulfur bridges, or complex polycyclic structures [137,138]. Advances in metagenomics, genome mining, and synthetic biology offer powerful tools to identify and optimize these underused scaffolds, potentially unlocking new antiviral pharmacophores with mechanisms distinct from those found in traditional plant-derived compounds.
Complementary to these discovery-driven approaches, late-stage functionalization (LSF) strategies, including photochemical transformations, provide a powerful means to expand chemical diversity directly from complex natural alkaloid scaffolds. By enabling selective modification at advanced synthetic stages, LSF allows the generation of novel scaffold variants and unexplored chemical space that may not be accessible through biosynthesis or de novo synthesis alone, thereby broadening the pool of potential antiviral pharmacophores.
Resistance considerations also shape the translational landscape. Traditional antiviral development often suffers from the “one drug- one bug” limitation, where each compound is tailored to a single virus. Because many alkaloids act on conserved host pathways or multiple viral targets, they may avoid this bottleneck, reducing the likelihood of rapid resistance. Combination therapies represent another promising avenue [134]. Berberine’s ability to enhance antibiotic activity via efflux inhibition and immunomodulation suggests that pairing alkaloids with established antivirals could yield synergistic effects [139]. Still, such strategies require thorough pharmacokinetic and toxicological evaluation before advancing to clinical stages.
Major methodological and infrastructural challenges persist. Many RNA viruses of public health concern require high-containment biosafety laboratories, which remain limited globally. As a result, many studies rely on pseudoviruses or computational models rather than live virus assays. RT-qPCR is used to quantify viral RNA, but it does not measure infectious virus. Viral titration assays which directly quantify infectious particles remain the gold standard for assessing antiviral efficacy. Pharmacokinetic analyses and tissue distribution studies are rare, and in vivo experiments often rely on short-term endpoints rather than detailed immunological or virological profiling. More robust preclinical and clinical work is needed to establish dosing, optimize structures, and generate high-quality efficacy data [135,140].
Vaccination remains a central pillar of RNA virus control. Yet rapid antigenic evolution, as seen in influenza A virus and SARS-CoV-2, limits vaccine durability [141]. Newer mRNA and vector-based platforms improve adaptability, but they cannot replace the need for broad-spectrum antivirals. Alkaloids, with their diverse mechanisms and host-targeted effects, may serve as complementary tools alongside vaccination to strengthen preparedness for future outbreaks.
These findings highlight both the promise and the challenges of alkaloid-based antiviral development. Their structural diversity, multi-target mechanisms, and activity across unrelated RNA viruses make them compelling candidates for next-generation therapeutics. Yet meaningful progress will require deeper mechanistic validation, expanded in vivo research, improved pharmacokinetic and toxicity profiling, and better access to biosafety level laboratories infrastructure. By combining modern drug development strategies with the chemical richness of alkaloids, it may be possible to generate broad-spectrum antivirals capable of responding quickly and effectively to future RNA virus threats.
6. Conclusions
Overall, the evidence shows that alkaloids form a rich and versatile source of antiviral candidates, acting through multiple conserved mechanisms including RdRp inhibition, blockade of viral entry, disruption of glycoprotein maturation, and modulation of host immune pathways. Several compounds, such as lycorine, emetine, homoharringtonine, cepharanthine, berberine, and palmatine, demonstrate broad spectrum activity across diverse RNA viruses, suggesting that alkaloid scaffolds are well suited for addressing the rapid evolution and cross species transmission typical of these pathogens.
Yet significant gaps hinder clinical translation. Most studies depend on in vitro assays without confirming true viral suppression, in vivo evidence remains limited, and toxicity, bioavailability, and pharmacokinetic profiles are poorly characterized. Many alkaloids also have narrow therapeutic windows, and the lack of high containment biosafety facilities restricts more rigorous evaluation. To advance these compounds meaningfully, future research must prioritize standardized antiviral testing, comprehensive toxicology, improved delivery strategies, and exploration of combination therapies. With such efforts, alkaloids could evolve from promising laboratory findings into antivirals for future RNA virus threats.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Forterre P. Prangishvili D. The origin of viruses Res. Microbiol.200916046647210.1016/j.resmic.2009.07.00819647075 · doi ↗ · pubmed ↗
- 2Baltimore D. Expression of animal virus genomes Bacteriol. Rev.19713523524110.1128/br.35.3.235-241.19714329869 PMC 378387 · doi ↗ · pubmed ↗
- 3Gebreyes W.A. Dupouy-Camet J. Newport M.J. Oliveira C.J.B. Schlesinger L.S. Saif Y.M. Kariuki S. Saif L.J. Saville W. Wittum T. The Global One Health Paradigm: Challenges and Opportunities for Tackling Infectious Diseases at the Human, Animal, and Environment Interface in Low-Resource Settings P Lo S Neglected Trop. Dis.20148 e 325710.1371/journal.pntd.0003257 PMC 423084025393303 · doi ↗ · pubmed ↗
- 4Ludwig B. Kraus F.B. Allwinn R. Doerr H.W. Preiser W. Viral zoonoses—A threat under control?Intervirology 200346717810.1159/00006974912684545 · doi ↗ · pubmed ↗
- 5Tao K. Tzou P.L. Nouhin J. Gupta R.K. de Oliveira T. Kosakovsky Pond S.L. Fera D. Shafer R.W. The biological and clinical significance of emerging SARS-Co V-2 variants Nat. Rev. Genet.20212275777310.1038/s 41576-021-00408-x 34535792 PMC 8447121 · doi ↗ · pubmed ↗
- 6V’kovski P. Kratzel A. Steiner S. Stalder H. Thiel V. Coronavirus biology and replication: Implications for SARS-Co V-2Nat. Rev. Microbiol.20211915517010.1038/s 41579-020-00468-633116300 PMC 7592455 · doi ↗ · pubmed ↗
- 7Mlcochova P. Kemp S.A. Dhar M.S. Papa G. Meng B. Ferreira I.A.T.M. Datir R. Collier D.A. Albecka A. Singh S. SARS-Co V-2 B.1.617.2 Delta variant replication and immune evasion Nature 202159911411910.1038/s 41586-021-03944-y 34488225 PMC 8566220 · doi ↗ · pubmed ↗
- 8Karim S.S.A. Karim Q.A. Omicron SARS-Co V-2 variant: A new chapter in the COVID-19 pandemic Lancet 20213982126212810.1016/S 0140-6736(21)02758-634871545 PMC 8640673 · doi ↗ · pubmed ↗