Enhanced Soybean Immunity to the Soybean Mosaic Virus Through RNA Interference Targeting the CP Gene
Tao Wang, Le Gao, Liqun Wang, Rui Ren, Rui Zhai, Xu Wang, Fuming Xiao, Long Yan, Xiaotong Lei, Tongtong Jin, Haijian Zhi

TL;DR
Scientists used RNA interference to boost soybean resistance to a virus without affecting plant growth.
Contribution
A novel RNAi strategy targeting the CP gene provides broad resistance to multiple viruses in soybeans.
Findings
Four transgenic soybean lines showed moderate to high resistance to the soybean mosaic virus.
Transgenic plants were also resistant to three other related viruses.
RNAi did not negatively impact soybean growth or yield traits.
Abstract
The soybean mosaic virus (SMV), a significant viral pathogen impacting soybean cultivation, leads to substantial yield losses and diminishes seed quality. In a prior study, we developed a targeted silencing vector using RNA interference (RNAi) technology targeting the CP gene, which codes for the viral coat proteins in the SMV genome. This vector was delivered into soybean plants through Agrobacterium-mediated transformation. In our current research, we utilized ongoing molecular characterization and resistance screening to identify four genetically pure lines that display moderate to high resistance to SMV. Additionally, the transgenic plants exhibited resistance to three other potyviruses: the bean common mosaic virus, the recombinant soybean mosaic virus, and the watermelon mosaic virus. Greenhouse and field trials conducted with these lines demonstrated that RNAi-mediated silencing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
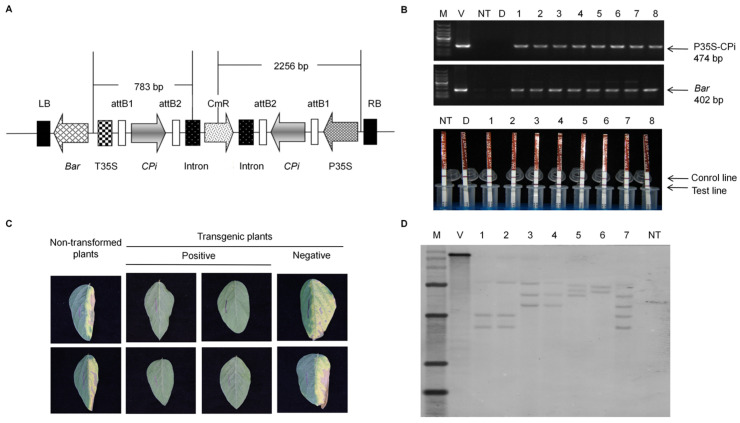 Figure 1
Figure 1 Figure 2
Figure 2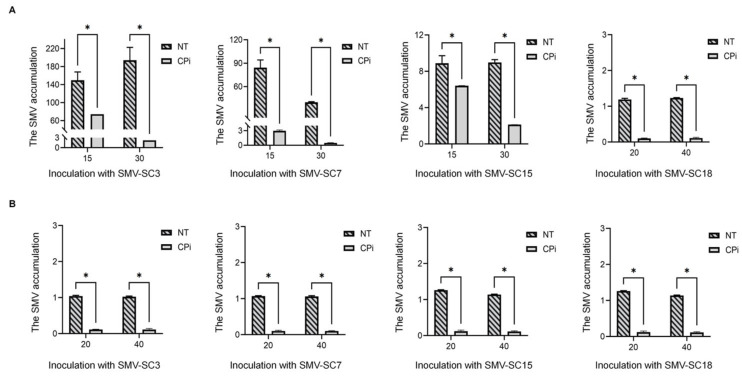 Figure 3
Figure 3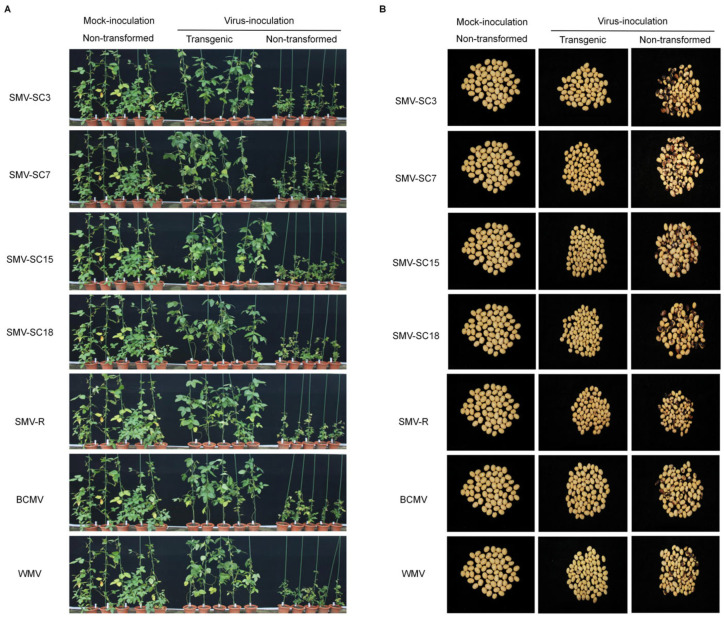 Figure 4
Figure 4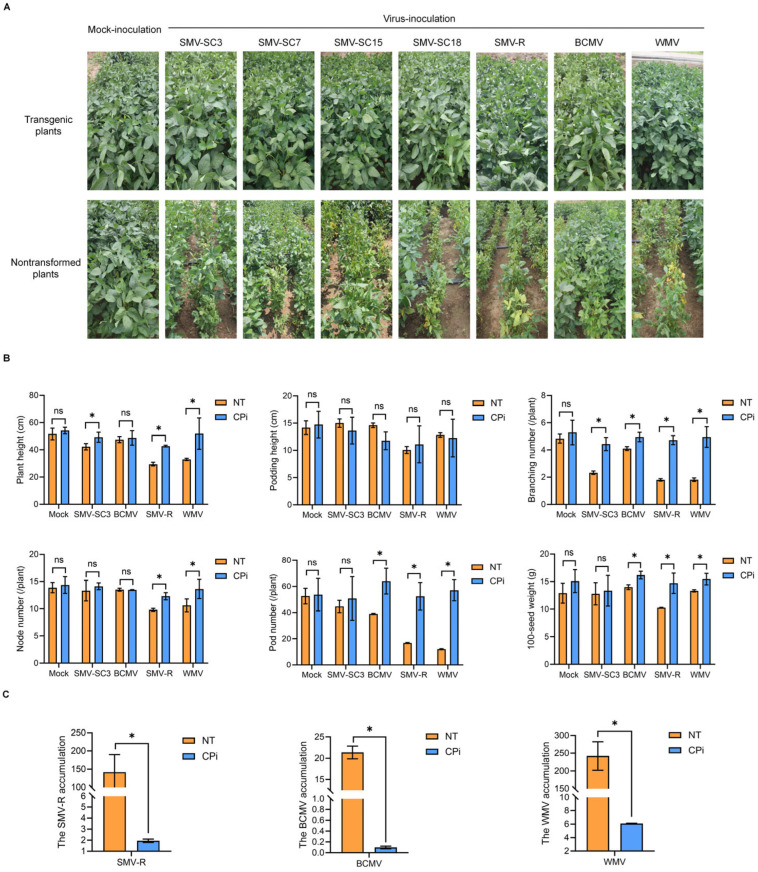 Figure 5
Figure 5- —Science and Technology Innovation Team for the Soybean Modern Seed Industry in Hebei
- —Construction of the Soybean Industry Innovation Team under the Hebei Modern Agricultural Industrial Technology System
- —Jiangsu Collaborative Innovation Center for Modern Crop Production (JCIC-MCP)
- —Core Technological Development for Breeding Program of Jiangsu Province
- —R&D Program of Beijing Municipal Education Commission
- —the Scientific Research Innovation Team Project of Beijing Vocational College of Agriculture
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Virus Research Studies · Transgenic Plants and Applications · Plant tissue culture and regeneration
1. Introduction
Soybean mosaic virus (SMV) disease is a significant biotic stress affecting the global soybean industry. The disease typically results from a complex of viruses and has a severe impact [1]. The primary pathogen, SMV, is a member of the Potyviridae family and comprises a single-stranded RNA of the positive sense [2]. This virus spreads primarily through non-persistent aphid transmission and seed-borne vectors [3,4]. In major soybean-producing regions, SMV causes an average annual yield loss of 15–30%, with severe localized outbreaks exceeding 50% in some years, making it a critical constraint on stable and high-yield production [5,6]. Due to the lack of effective chemical control agents, traditional management heavily relies on breeding disease-resistant varieties. However, SMV exhibits complex strain differentiation and high mutation rates [7,8], posing potential risks that could allow new variants to overcome existing resistance.
RNA interference (RNAi) technology, which induces the degradation of target gene mRNA through a mechanism involving specific double-stranded RNA [9], has opened new avenues for plant virus-resistant breeding. In controlling tomato yellow leaf curl virus, constructing RNAi vectors targeting multiple viral genes increased disease resistance in transgenic plants by 80% [10]. For rice stripe virus resistance improvement, interference vectors targeting viral coat protein and movement protein genes enabled transgenic plants to exhibit full life-cycle immunity against the virus [11]. These findings demonstrate that RNAi technology can achieve cross-species and multi-target viral resistance enhancement with stable and long-lasting protective effects.
Recent advancements in RNAi technology for SMV research have achieved new breakthroughs in targeted interference of viral proteins and host susceptibility genes. The HC-Pro protein, serving as a viral gene-silencing suppressor [12], has become a key target for RNAi applications. Transgenic soybean lines developed with RNAi vectors targeting HC-Pro gene demonstrated significantly enhanced disease resistance against multiple SMV strains [13,14]. Notable advancements have also been realized in bolstering resistance to P3 protein interference; by employing RNAi to target the P3 gene, the virus accumulation in transgenic soybeans was diminished by 72% [15]. Additionally, RNAi studies on the host susceptibility gene GmVma12 revealed that transgenic soybeans exhibited stronger resistance to the SMV-SC15 strain, with markedly reduced seed mottling symptoms [16].
The coat protein (CP) of SMV serves as the primary structural component of viral particles. It plays crucial roles in viral particle assembly, stabilization, and mediating viral movement between host cells, while also acting as a key factor in viral–host interactions [17,18]. Sequence variations in the CP gene directly influence viral pathogenicity and strain differentiation, making it a critical target for antiviral research. In previous research, we constructed a vector that interfered with CP and conducted genetic transformation. In the present study, viruses were inoculated into transgenic soybean plants. The findings indicated that the transgenic plants exhibited moderate to high resistance against the prevalent SMV strains, resulting in a significant reduction in the accumulation of viruses within the plants and the appearance of symptoms. This provides an efficient technical solution for molecular breeding of virus-resistant soybeans.
2. Results
2.1. Generation of Transgenic Soybean Plants Containing the SMV CP Hairpin
In previous studies, a 264-bp fragment (8732–8995 nt) was selected as the target sequence for constructing an RNAi expression vector containing its reverse repeat fragments, based on the conserved sequence of the SMV CP gene identified in GENBANK (Figure 1A) [19]. The SMV-CP RNAi (CPi) fragment was introduced into the soybean cultivar Huachun 6 using Agrobacterium-mediated transformation. A total of 2963 explants were transformed, and following differentiation, screening, and regeneration, 89 herbicide-resistant regenerated plants were obtained.
To obtain stable transgenic lines, continuous screening and purification of T_0_–T_3_ generation transgenic plants were conducted using PCR detection, LibertyLink^®^ strip analysis, and herbicide resistance testing. The PCR results indicated that transgenic plants exhibited the expected amplification for the CPi fragment (474 bp) and the bar gene (402 bp), corresponding to the predicted sizes. In contrast, no such bands were detected in the non-transformed plants (Figure 1B). The LibertyLink^®^ strip test results indicated that the transgenic plants were able to detect both the control and test lines. In contrast, only the control line was detected in the non-transformed plant (Figure 1B), confirming the presence of the bar protein in these plants, rather than its absence in the transgenic ones. The herbicide tolerance assay results demonstrated that the leaves of transgenic plants exhibited a distinct herbicide-resistant phenotype, following treatment with 200 mg/L PPT. In contrast, non-transformed plants exhibited significant yellowing and curling after herbicide application (Figure 1C). These results indicate that the RNAi-CP vector has been successfully transformed and is normally expressed in the plant.
Southern blot analysis was conducted to confirm that the CP hairpin was successfully integrated into the genome of T_1_ soybean plants. Seven PCR-positive transgenic soybean plants were selected for Southern blot hybridization, using the CPi sequence as the probe. The results indicated that all seven transgenic plants exhibited hybridization signals, whereas no bands were observed in the non-transformed plants (Figure 1D), signifying successful integration of the foreign fragment into the soybean genome. Additionally, the transgenic plants displayed two to four copies of T-DNA insertions.
2.2. The Transgenic Plants Exhibiting Resistance to Herbicides Also Demonstrated Significant Resistance to SMV-SC3
To determine whether transgenic plants exhibit significantly different resistance to SMV compared to non-transformed plants, we conducted a resistance evaluation experiment. The visual responses of the fully expanded first (V_1_), second (V_2_), third (V_3_), and fourth (V_4_) trifoliate leaves of T_2_ and T_3_ transgenic plants and non-transformed plants inoculated with SMV-SC3 were observed. The results were as follows: 14 T_2_ generation plants and 32 T_3_ generation plants exhibited no symptoms of SMV infection and were classified as highly resistant (HR); 3 T_2_ generation plants showed mild early-stage symptoms that later resolved, classified as delayed resistance (DR); 44 T_2_ generation plants and 83 T_3_ generation plants displayed delayed mosaic or mild mosaic symptoms, classified as mildly resistant (MR); and 23 T_2_ generation plants and 2 T_3_ generation plants showed mosaic and curling symptoms throughout the four growth stages, classified as susceptible (S). The 90 non-transformed control plants exhibited susceptibility at all four stages (Table 1). Then, the symptoms of both non-transformed and transgenic plants inoculated with SMV-SC3 at the V_4_ stage were observed. Figure 2A shows that non-transformed plants displayed mosaic and curling symptoms in their V_4_ leaves. In contrast, the highly resistant T_2_ and T_3_ generation transgenic plants exhibited no mosaic symptoms and were indistinguishable from the mock-inoculated control plants. The mildly resistant T_2_ and T_3_ generation transgenic plants displayed relatively mild mosaic and curling symptoms.
Additionally, we conducted follow-up observations on the subsequent symptoms of the infected plants and performed virus content tests. The severity of mosaic disease in the top trifoliate leaves was evaluated at 8 weeks post-inoculation, as depicted in Figure 2B. Grade 0 indicates an absence of symptoms; Grade 1 signifies mild mosaic symptoms; Grade 2 denotes noticeable mosaic symptoms; Grade 3 features mosaic symptoms along with leaf curling; and Grade 4 indicates severe mosaic symptoms and leaf curling. The results showed that 84 T_2_ generation plants had an average disease severity of Grade 1, whereas 117 T_3_ generation plants exhibited no symptoms (Grade 0). In comparison, non-transformed plants averaged around Grade 4 in severity (Table 2 and Figure 2C). The accumulation of SMV in transgenic plants inoculated with SMV-SC3 and non-transformed plants inoculated with SMV-SC3 was detected using DAS-ELISA. The results indicated that the viral accumulation in T_2_ and T_3_ plants was less than double that of the negative control (non-transformed plants inoculated without SMV), whereas the viral accumulation in non-transformed plants was 10.6 times that of the negative control (Table 3). The results demonstrated that the transgenic positive plants exhibited resistance to herbicides and a high level of disease resistance to SMV-SC3.
2.3. The Transgenic Plants Exhibited Robust Resistance to Numerous Strains of SMV
Under dual testing conditions involving herbicides and SMV-SC3, four transgenic lines exhibiting high resistance to SMV-SC3 were identified. T_3_ generation plants from these lines were inoculated with other SMV strains (SMV-SC7, SMV-SC15, and SMV-SC18) to evaluate their resistance responses. The results indicated that at 15 and 30 dpi, the transgenic plants displayed significantly lower levels of SMV CP gene transcription compared to non-transformed plants. In particular, following inoculation with SMV-SC7 and SMV-SC18, the transgenic plants showed less than 0.1% of the CP gene transcription levels observed in non-transformed plants. Over time, by day 30 post-inoculation, the SMV CP gene transcription levels in the transgenic plants gradually decreased, whereas the non-transformed plants maintained relatively high levels (Figure 3A). Subsequently, the levels of viral accumulation in both transgenic and non-transformed plants were measured at 20 and 40 dpi. The results indicated that non-transformed plants sustained high viral accumulation at 20 and 40 dpi after inoculation with various strains of SMV, reaching levels 9.5 to 11.9 times higher than those observed in transgenic plants (Figure 3B). These findings suggest that transgenic plants demonstrate robust resistance to multiple strains of SMV.
2.4. The Transgenic Plants Infected with Potyviruses Maintained the Original Agronomic Traits and Yield of the Recipient Plants
Transgenic T_4_ generation plants and non-transformed plants were planted both in greenhouses and fields, and inoculated with multiple potyviruses—four SMV strains, BCMV, SMV-R, and WMV—to investigate differences in agronomic traits between the two groups. These traits included maturity period, plant height, podding height, branching number, node number, pod number, and 100-seed weight. The results indicated that non-transformed plants in both greenhouse and field settings exhibited leaf curling and stunted growth after inoculation with different viruses, whereas the transgenic plants did not display stunting symptoms similar to those of their non-transformed counterparts (Figure 4A and Figure 5A). A morphological analysis of the harvested seeds indicated that non-transformed seeds inoculated with viruses primarily developed brown seed-coat mottling, whereas transgenic seeds inoculated with these viruses rarely showed such symptoms (Figure 4B).
A comparative agronomic analysis was conducted between non-transformed plants and transgenic plants subjected to mock treatment. The results demonstrated that no significant differences were observed between the transgenic and non-transformed plants in key traits such as plant height, podding height, branching number, nod number, pod number, 100-seed weight, and duration of maturity (Figure 5B, Table S2). However, it is noteworthy that non-transformed plants inoculated with different viruses exhibited reduced plant height, branching number, pod number, and 100-seed weight compared to transgenic plants inoculated with the same virus (Figure 5B).
Following inoculation with four viral strains (SMV-SC3, BCMV, SMV-R, and WMV), the agronomic performance of non-transformed plants was compared with that of transgenic lines, to evaluate the differences in viral resistance. The results showed that non-transformed plants suffered significant reductions in multiple agronomic traits relative to transgenic plants, with the damage severity varying substantially among virus treatments. Specifically, after SMV-SC3 inoculation, non-transformed plants exhibited a 14.24% decrease in plant height and a 47.40% reduction in branching number. Under BCMV infection, the branching number, pod number and 100-seed weight of non-transformed plants declined by 17.17%, 39.11% and 13.54%, respectively. Notably, SMV-R caused the most severe damage to non-transformed plants, leading to reductions of 30.96% in plant height, 61.86% in branching number, 20.33% in nod number, 67.73% in pod number, and 30.27% in 100-seed weight. In the WMV treatment group, non-transformed plants showed the largest declines in plant height (36.69%) and pod number (78.74%), accompanied by decreases of 63.43% in branching number, 22.08% in nod number, and 11.78% in 100-seed weight. Collectively, these findings indicated that transgenic soybean plants had stronger resistance to the tested viral strains than non-transformed plants.
Additionally, both non-transformed and transgenic plants exhibited no significant differences in podding height, regardless of their status of inoculation with different viruses (Table S2). The findings suggest that transgenic plants do not display significant differences in key agronomic traits compared to non-transformed plants, and their inherent resistance to potyviruses contributes to more effectively stabilizing soybean yield and quality.
3. Discussion
Current research has identified major resistance genes against SMV located on chromosomes 2, 13, and 14 [20,21,22]. The resistance gene on chromosome 2 encodes an RNase that interacts with SMV’s P3 protein, degrading double-stranded RNA during viral replication to trigger resistance responses [23]. However, this resistance has been overcome by emerging variants [24]. Resistance genes on chromosomes 13 and 14 are confirmed as nucleotide-binding domain and leucine-rich repeat genes [25,26], which mediate resistance through interactions with SMV’s HC-Pro and CI proteins, respectively [27,28]. Nevertheless, the large genomic size of these two resistance genes makes it challenging to develop resistant varieties through conventional genetic engineering techniques [29]. Compared to the introduction of resistance genes into host plants, RNAi technology has demonstrated numerous significant advantages in the field of resistance breeding research. Firstly, RNAi technology requires relatively simple operations, eliminating the need for complex gene cloning and transformation procedures. By simply designing and introducing appropriate interference sequences, research cycles can be significantly shortened and costs reduced [30]. Secondly, RNAi technology has high specificity, allowing for precise targeting of specific viral genes, which effectively inhibits viral replication and transmission without adversely affecting other host plant genes. In contrast, introducing resistance genes may trigger unforeseen genetic interactions that could disrupt the normal growth and development of the host plant [31]. Furthermore, RNAi technology offers broad-spectrum resistance coverage. A single interference vector can be engineered to target multiple viral genes or different virus genera simultaneously, achieving multi-resistance or broad-spectrum protection [32]. These advantages ensure that RNAi technology continues to play a significant role in the field of resistance research and breeding applications.
The CP gene of the soybean mosaic virus is crucial for its replication, assembly, and movement between cells. To enhance plant defenses against pathogens, researchers have employed two primary strategies—gene silencing and gene overexpression—to investigate the operational mechanisms of the CP. The technique of gene silencing adeptly curtails CP expression via targeted RNAi pathways, leading to a diminished production of viral coat proteins and a marked decrease in the virus’s reproductive efficiency within the plant [33,34]. In contrast, the strategy of gene overexpression increases CP transcription in plant cells through genetic modification, thereby enhancing the plants’ ability to resist viral infections [35]. Findings have revealed that silencing the CP in plants significantly increased the resistance of transgenic soybeans to the SMV, effectively halting viral replication and transmission. Similarly, the overexpression of CP also endowed transgenic plants with enhanced viral resistance, presumably due to the surplus CP disrupting the viral lifecycle or provoking plant immune responses, thus restraining viral growth and dissemination [36]. In summary, these findings offer a robust theoretical framework and technical support for the development of novel virus-resistant soybean cultivars.
This study successfully developed an efficient RNAi vector targeting the SMV CP gene and obtained genetically stable resistant lines through Agrobacterium-mediated soybean transformation. When inoculated with multiple SMV strains, these resistant lines demonstrated enhanced SMV resistance compared to non-transformed plants. Previous studies have also demonstrated that interfering with the CP gene in SMV can produce similar resistance-enhancement effects [33,34], further confirming that RNAi-CP is an effective strategy for resistance improvement and breeding. Compared with similar studies, our work innovatively maintains key agronomic traits in resistant lines, while effectively preserving soybean yield and seed quality compared to susceptible plants.
Through sequence alignment (Figure S1), four SMV strains, SMV-R, BCMV, and WMV, were found to exhibit high sequence homology with one another. Among these strains, SMV-SC3 shared 100% homology with the CP interference fragment, whereas BCMV showed the lowest homology, at 75%; the remaining viral strains displayed homology levels ranging from 86% to 97%. This high degree of sequence similarity suggests that the CP genes of these viruses are susceptible to RNAi-mediated silencing upon invading the transgenic plants, which consequently confers broad-spectrum viral resistance to the transgenic plants.
Notably, non-transformed plants inoculated with BCMV, SMV-R, or WMV all exhibited typical dwarfing phenotypes, but the extent of agronomic trait damage varied distinctly among the viral treatments. Specifically, non-transformed plants infected with SMV-R and WMV suffered severe and comprehensive damage to multiple agronomic traits, including plant height, branching number, node number, pod number, and 100-seed weight. In contrast, BCMV infection only affected the branching number, pod number, and 100-seed weight of non-transformed plants, with no significant alterations detected in plant height and node number.
Consistent with the phenotypic observations, qRT-PCR analysis revealed that the viral mRNA accumulation levels in non-transformed plants were significantly higher than those in transgenic plants across all three viral treatments (Figure 5C). Interestingly, the viral mRNA abundance in BCMV-infected non-transformed plants was notably lower than that in plants inoculated with SMV-R or WMV. Based on these findings, we hypothesize that BCMV exhibits relatively lower virulence in the soybean cultivar Huachun 6 compared with SMV-R and WMV, which accounts for the less severe alterations in agronomic traits observed in BCMV-infected plants.
4. Materials and Methods
4.1. Materials
The viruses used in this study were SMV-SC3, SMV-SC7, SMV-SC15, SMV-SC18, recombinant soybean mosaic virus (SMV-R), bean common mosaic virus (BCMV), and watermelon mosaic virus (WMV), as well as Agrobacterium EHA105 and interference expression vector pB7GWIWG2 (II), which were provided by the National Soybean Improvement Center of Nanjing Agricultural University (Nanjing, China). The transgenic recipient Huachun 6 was kindly provided by Professor Hai Nian from South China Agricultural University (Guangzhou, China). The Escherichia coli DH5α used for vector construction and the intermediate vector pDONR221 were purchased from TianGen Biochemical Technology Co., Ltd. (Beijing, China) and Invitrogen Corporation, Inc. (Carlsbad, CA, USA), respectively.
4.2. Identification of Transgenic Plants
In our initial studies, we effectively assembled an interference vector, pB7GWIWG2(II)-CPi, which includes the CP gene fragment. The recombinant vector pB7GWIWG2(II)-CPi contains a plant-selection marker, namely the herbicide resistance gene bar, which confers resistance to phosphinothricin (PPT) (Figure 1A). By employing a cotyledonary node–Agrobacterium-mediated soybean transformation system, we successfully generated T_0_ transgenic soybean plants. For comprehensive details on vector construction and tissue culture methods, please consult the research by Gao et al. [19].
The T_1_–T_3_ generation of transgenic plants was cultivated in the greenhouse at Pailou, Nanjing Agricultural University. At the first trifoliate leaf stage, the transgenic plants were identified using the leaf-painting method, the LibertyLink^®^ strip detection method, and the PCR detection method. Pure lines were then selected for further study.
The leaf-painting method involved diluting the PPT solution to a concentration of 200 mg/L. One half of a leaf was marked using a marker pen, with the main leaf vein serving as the boundary, and the other half was brushed with the diluted solution. After one week of cultivation, the reaction of the herbicide on the leaves was observed.
LibertyLink^®^ strip detection was conducted using a QuickStix^TM^ Kit (EnviroLogix, Inc. (Portland, ME, USA)) to verify the expression of the bar gene. The process includes the following steps: place 100 mg of fresh leaf tissue into a 1.5 mL centrifuge tube. Using a grinding rod, rotate the tube to extract the juice, then add 0.5 mL of extraction buffer for dilution and mixing. Insert the test strip into the centrifuge tube, following the arrow direction. After a 5 min incubation period, observe whether the detection line develops coloration.
The PCR detection protocol involves extracting DNA from fresh leaves at 100 mg/L using an efficient plant genomic DNA extraction kit (DP350, TianGen Biochemical Technology Co., Ltd. (Beijing, China)). The insertion fragments of CPi and bar were amplified using Premix Taq (R004A, TaKaRa Bio, Inc. (Beijing, China)), with detection primers 35S-P/attB2-CPi-R and bar-F/bar-R synthesized by General Biotech (Anhui) Co., Ltd. (Anhui, China). The reaction system consisted of 25 μL Premix Taq, 1 μL template (<500 ng), 1 μL each of upstream and downstream primers, and 22 μL sterilized water. The reaction was amplified on a T100^TM^ gradient PCR instrument (Bio-Rad Laboratories, Inc. (Hercules, CA, USA)) with the following program: 98 °C pre-denaturation 10 s, 55 °C denaturation 30 s, 72 °C extension 1 min, for 30 cycles. The products were analyzed by 1% agarose gel electrophoresis, and the DNA molecular markers used were purchased from Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China).
4.3. Southern Blot Hybridization Analysis
Genomic DNA from T_1_ generation transgenic plants was extracted using an efficient plant genome DNA extraction kit. Approximately 30 μg of DNA samples were digested with EcoRI enzyme (Thermo Fisher Scientific, Inc. (Waltham, MA, USA)), which specifically cleaves the T-DNA region. The digested DNA was electrophoretically separated on a 0.8% agarose gel and transferred to Hybond-N+ nylon membrane (Amersham, (Buckinghamshire, UK)). A 264-bp fragment containing the CP gene coding region of pB7GWIWG2(II)-CPi plasmid was amplified using attB1-CPi-F/attB2-CPi-R primers. Primer sequence information is shown in Table S1. The product was subsequently labeled with digoxin (DIG) for high-intensity detection (Hoffmann-La Roche, Inc. (Indianapolis, IN, USA)), and the labeled product was used as the probe. Subsequent hybridization, film washing, and signal detection were performed according to the instructions provided in the DIG High Prime DNA Labeling and Detection Kit II (Hoffmann-La Roche, Inc. (Indianapolis, IN, USA)).
4.4. SMV Inoculation and Resistance Assessment
T_2_–T_3_ generation plants and non-transformed plants were grown in greenhouses. After the young leaves unfolded, SMV-SC3 was inoculated using the methodology described by Gao et al. (2015) [13]. The responses of the T_2_ and T_3_ plants were visually observed during the V_1_–V_4_ stages. These responses were classified into four categories: HR-type (highly resistant to SMV, with no viral symptoms observed during the study period); DR-type (delayed resistance manifestation, with early viral symptoms that later disappeared); MR-type (mildly resistant, with delayed viral symptoms or mild symptoms compared to susceptible controls); and S-type (highly susceptible, with viral symptoms consistent with controls across all four stages). Additionally, the severity ratings for SMV were assessed on the top three fully developed leaves at 8 weeks post-inoculation (the topmost, second, and third layers), with the average leaf ratings indicating the overall plant infection levels. The disease severity was categorized into five grades: Grade 0 (asymptomatic); Grade 1 (mild mosaic symptoms); Grade 2 (mosaic symptoms); Grade 3 (mosaic symptoms with leaf curling); and Grade 4 (mosaic symptoms with severe leaf curling).
4.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
Four SMV strains (SMV-SC3, SMV-SC7, SMV-SC15, and SMV-SC18) were inoculated into T_3_ generation transgenic plants and non-transformed plants. Leaves were collected at 15 and 30 days post-inoculation (dpi), and the viral accumulation in the plants was detected using qRT-PCR. Samples were transferred to centrifuge tubes, ground into powder with liquid nitrogen, and processed for RNA extraction and cDNA synthesis. Amplification was performed using the primers CP-testF/CP-testR and Tubulin-F/Tubulin-R (as the internal reference gene), with the CFX96 Touch real-time PCR detection system (Bio-Rad Laboratories, Inc. (Hercules, CA, USA)). In addition, leaves were collected from both non-transformed and transgenic plants at 30 dpi with SMV-R, BCMV, and WMV, respectively. The viral accumulation levels of these three viruses in the leaves were then detected using the primers CP-testF/CP-testR, BCMV-CP-testF/BCMV-CP-testR, and WMV-CP-testF/WMV-CP-testR, which correspondes to each virus. The data were analyzed employing the 2^−ΔΔCT^ method.
4.6. Serological Determination
Four SMV strains (SMV-SC3, SMV-SC7, SMV-SC15, and SMV-SC18) were inoculated into the T_3_ generation of transgenic plants and non-transformed plants. Leaves were collected at 20 and 40 dpi, and the viral accumulation in the plants was detected using a double-antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) kit (V094, NanoDian, Inc. (Los Angeles, CA, USA)). The collected leaves were placed in sampling tubes, ground into a powder, and analyzed for viral content using the Infinite^®^ 200 PRO (Tecan Group Ltd. (Männedorf, ZH, Switzerland)) with an OD405 nm reading. The values of transgenic and non-transformed plants were converted to multiples of the negative control. Samples with relative values exceeding twice the negative control were classified as positive.
4.7. Agronomic Traits of Transgenic Plants Under Greenhouse and Field Conditions
Transgenic T_4_ plants and non-transformed plants were cultivated under greenhouse and field conditions, respectively. Four strains of soybean mosaic virus and three other potyviruses—bean common mosaic virus (BCMV), recombinant soybean mosaic virus (SMV-R), and watermelon mosaic virus (WMV)—were inoculated into the transgenic lines and non-transformed plants. Forty days post-inoculation, both transgenic and non-transformed plants were photographed simultaneously. During harvest, at least nine plants from each treatment group were randomly selected for agronomic trait evaluation. Measurements were taken for maturity period, plant height, podding height, branching number, node number, pod number, and 100-seed weight (Table S2). Seed samples from both inoculated and non-inoculated plants were also collected for seed-coat mottling analysis.
4.8. Statistical Analysis
The Jamovi software (Version 1.6) [37] was utilized to analyze the differences in resistance levels between transgenic soybean plants and control recipient varieties, using a two-tailed Student’s t-test. Multiple comparisons were conducted using the least significant difference method when examining the variance of agronomic traits across various treatment groups. Variations were deemed statistically significant if the p-value was less than 0.05.
5. Conclusions
This study successfully generated four genetically stable, high-resistance lines by employing RNAi-mediated specific silencing of the SMV CP gene. The primary conclusions are as follows: firstly, the constructed pB7GWIWG2(II)-CPi vector can efficiently induce post-transcriptional silencing of the CP gene in soybean cells, significantly inhibiting SMV replication. Secondly, the screened resistant lines maintained normal growth and development under viral stress, with their critical agronomic traits consistent with those of the recipient. These findings not only provide new genetic resources and breeding materials for molecular breeding of soybean resistance to SMV, but also offer a replicable technical system for utilizing RNAi technology to control plant viral diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Widyasari K. Alazem M. Kim K.H. Soybean resistance to soybean mosaic virus Plants 2020921910.3390/plants 902021932046350 PMC 7076706 · doi ↗ · pubmed ↗
- 2Cho E.K. Goodman R.M. Evaluation of resistance in soybeans to soybean mosaic virus strains 1Crop Sci.1982221133113610.2135/cropsci 1982.0011183 X 002200060012 x · doi ↗
- 3Hill J.H. Alleman R. Hogg D.B. Grau C.R. First report of transmission of soybean mosaic virus and alfalfa mosaic virus by Aphis glycines in the new world Plant Dis.20018556110.1094/PDIS.2001.85.5.561C 30823148 · doi ↗ · pubmed ↗
- 4Pasin F. Daròs J.A. Tzanetakis I.E. Proteome expansion in the Potyviridae evolutionary radiation FEMS Microbiol. Rev.202246 fuac 01110.1093/femsre/fuac 01135195244 PMC 9249622 · doi ↗ · pubmed ↗
- 5Ren Q. Pfeiffer T.W. Ghabrial S.A. Soybean mosaic virus incidence level and infection time: Interaction effects on soybean Crop Sci.1997371706171110.2135/cropsci 1997.0011183 X 003700060005 x · doi ↗
- 6Hill J.H. Whitham S.A. Control of virus diseases in soybeans Adv. Virus Res.20149035539010.1016/B 978-0-12-801246-8.00007-X 25410106 · doi ↗ · pubmed ↗
- 7Seo J.K. Ohshima K. Lee H.G. Son M. Choi H.S. Lee S.H. Sohn S.H. Kim K.H. Molecular variability and genetic structure of the population of soybean mosaic virus based on the analysis of complete genome sequences Virology 20093939110310.1016/j.virol.2009.07.00719716150 · doi ↗ · pubmed ↗
- 8Gao L. Sun S. Li K. Wang L.W. Hou W.S. Wu C.X. Zhi H.J. Han T.F. Spatio-temporal characterisation of changes in the resistance of widely grown soybean cultivars to soybean mosaic virus across a century of breeding in China Crop Pasture Sci.20186939540510.1071/CP 17296 · doi ↗