Genome-Wide Identification, Characterization, Expression Analysis, and Interacting Protein Prediction of the GSK3/Shaggy-like Gene Family in Watermelon
Peng Tian, Jingjing Zhang, Bowen Liu, Xiurui Gao, Bing Li, Wei Liu, Yanrong Wu

TL;DR
This study identifies and characterizes the GSK3 gene family in watermelon, revealing their roles in hormone signaling and stress responses.
Contribution
The study provides the first genome-wide analysis of the GSK3 gene family in watermelon, including expression patterns and interacting proteins.
Findings
Eight ClGSK3 genes were identified and grouped into four subfamilies with conserved genomic micro-collinearity.
ClGSK3s are likely involved in hormone and stress responses, with ClSK21 interacting with brassinosteroid signaling components.
BR treatment significantly affects the expression of ClSK21, ClSK31, and ClSK41.
Abstract
Glycogen synthase kinase 3 (GSK3/Shaggy-like) is a highly conserved serine/threonine kinase that orchestrates growth, hormone signaling, and abiotic stress responses in both animals and plants, yet its role in watermelon remains unexplored. In this study, we conducted a whole-genome identification, identifying a total of eight members of the GSK3 gene family (ClGSK3) distributed across seven chromosomes. Phylogenetic and synteny analyses resolved the eight ClGSK3s into four subfamilies that display one-to-one or one-to-many orthology with Arabidopsis and rice GSK3 genes, indicating conserved genomic micro-collinearity across dicots and monocots. Predictions of cis-acting elements and transcriptome data analysis indicate that ClGSK3s may be involved in hormone- and stress-responsive conditions. Protein–protein interaction networks predicted 53 candidate partners for five ClGSK3 proteins;…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
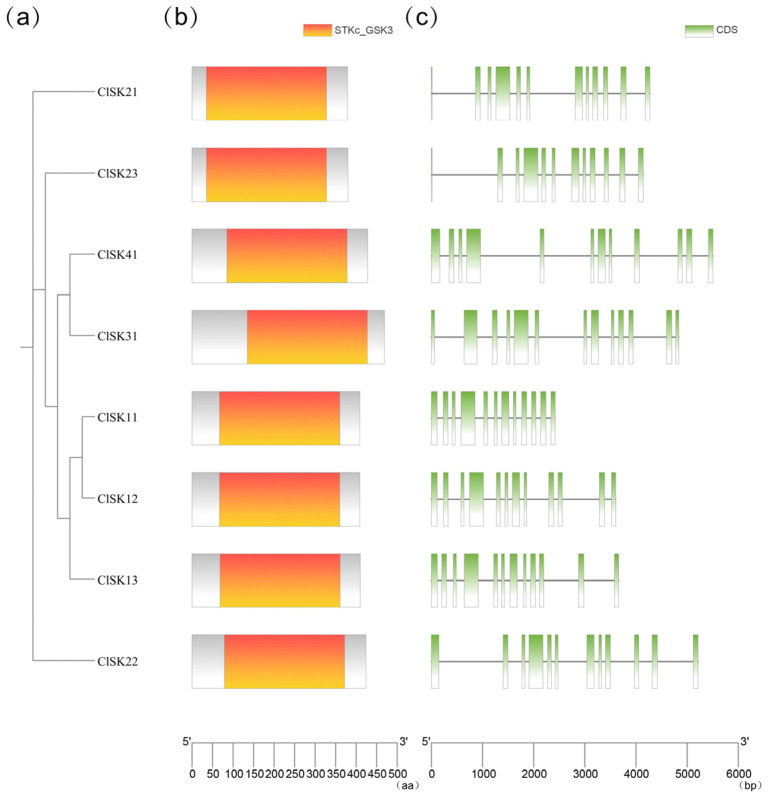 Figure 1
Figure 1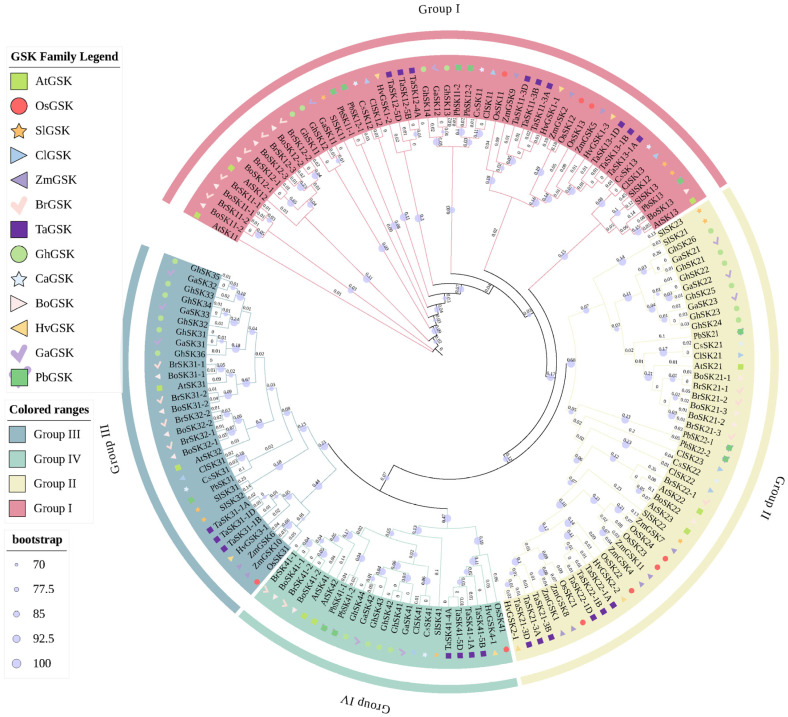 Figure 2
Figure 2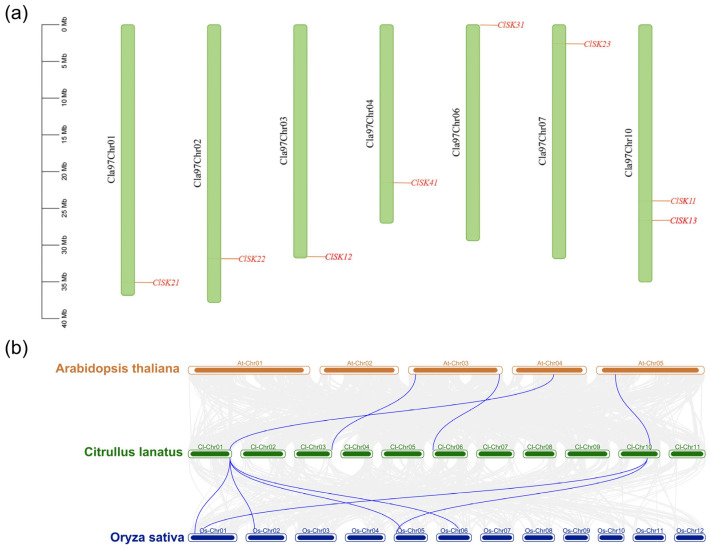 Figure 3
Figure 3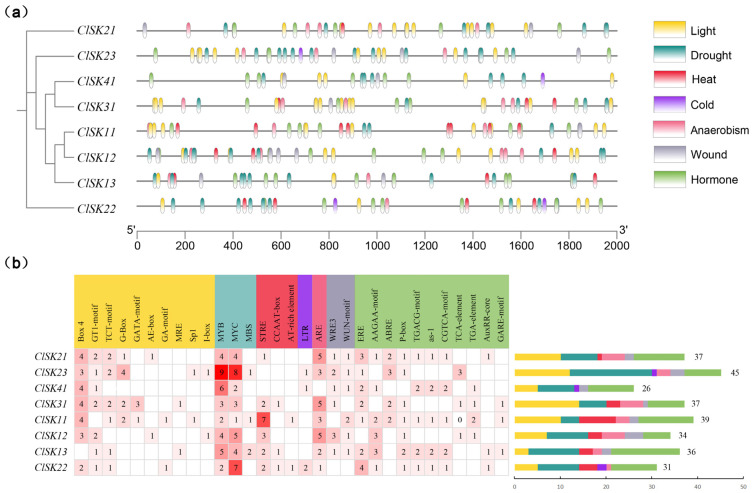 Figure 4
Figure 4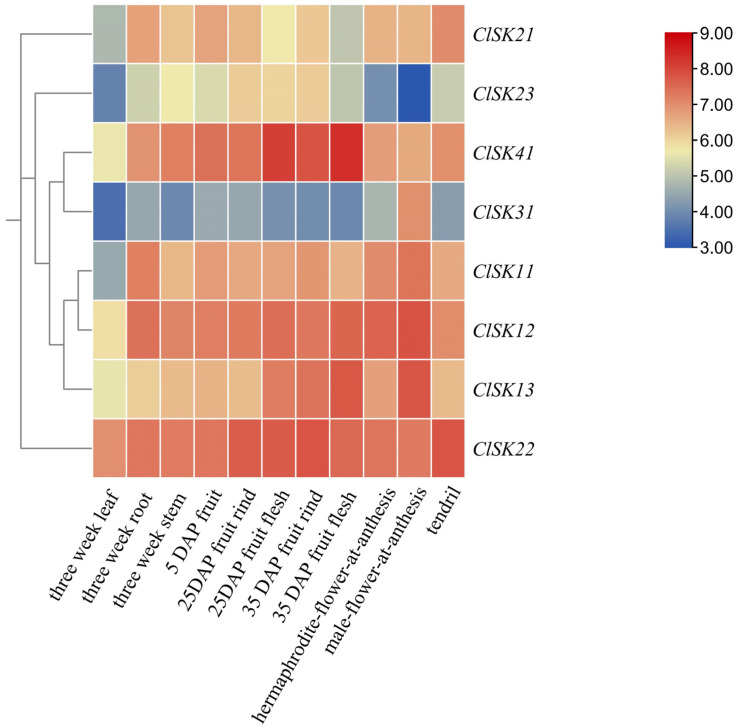 Figure 5
Figure 5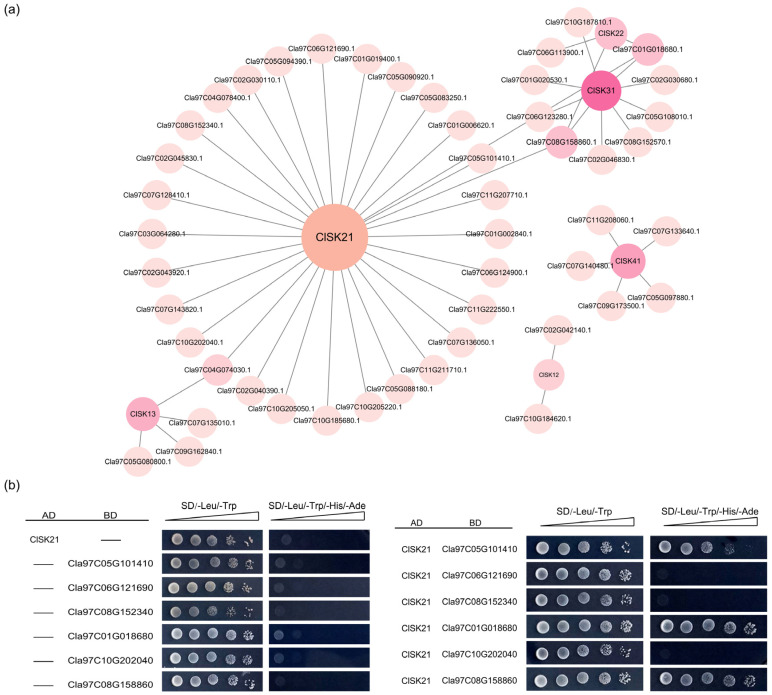 Figure 6
Figure 6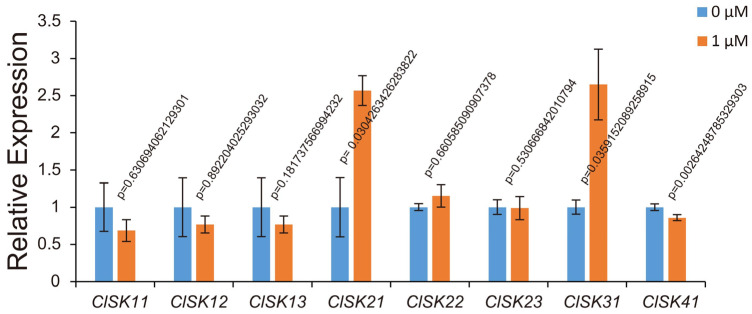 Figure 7
Figure 7- —National Industrial Technology System of Watermelon and Muskmelon
- —Hebei Open-field Vegetable Innovation Team of Modern Agro-industry Technology Research System
- —Key Research and Development Program Project of Hebei Province
- —Basic Research Funds of Hebei Academy of Agriculture and Forestry Sciences
- —Hebei Provincial Watermelon Breeding Technology Innovation Center Subsidy Fund Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPolysaccharides and Plant Cell Walls · Plant Molecular Biology Research · Advances in Cucurbitaceae Research
1. Introduction
Watermelon (Citrullus lanatus, 2n = 2× = 22), a member of the Citrullus genus in the Cucurbitaceae family, originated in Africa [1]. It is highly valued for its nutritional content and refreshing properties, making it a significant vegetable crop globally [2,3]. China has long been the leading producer and consumer of watermelon worldwide, with projections indicating its continued dominance in 2023 [4]. The focus of breeding efforts has shifted towards developing varieties with high yield, quality, and resistance [5]. Nevertheless, the molecular mechanisms governing watermelon growth, development, and stress responses are not well elucidated compared to other major crops [6,7].
Glycogen synthase kinase 3 (GSK3)/Shaggy-like kinases are a highly conserved family of serine/threonine protein kinases essential for various cellular and developmental processes in eukaryotes. In animals, GSK3 proteins are involved in signal transduction pathways regulating glycogen synthesis, insulin activity, transcriptional activation, and development [8]. Similarly, in plants, the GSK3 gene family plays a critical role in physiological responses like osmotic stress, wound healing, and Oleandrin lactone signaling [9]. The first plant GSK3 gene was identified in Medicago sativa. With the advancement of genome sequencing in various plant species, comprehensive studies on the identification and functional analysis of GSK3 gene families have been carried out in crops such as wheat, Nicotiana tabacum and pear [10,11,12]. The GSK3 genes are typically categorized into four subfamilies, mirroring the classification of their homologous genes [13,14].
The GSK3 genes have been extensively studied in model species such as Arabidopsis and rice. They are implicated in hormone signaling and environmental adaptation. For instance, in Arabidopsis, BIN2 (AtSK21), a well-studied GSK3 family member, phosphorylates various substrates like EGL3, TTG1, BZR1, and SnRK2.2, influencing root hair formation, Brassinosteroid (BR) and Abscisic Acid (ABA) signaling pathways, among others [15,16,17,18,19,20,21]. AtSK11, AtSK22, and AtSK31 are known to enhance plant salt tolerance [19,22,23]. In rice, OsSK21 is highly expressed in the panicle and plays a crucial role in determining rice plant morphology, panicle architecture, and resistance to abiotic stress [24,25]. OsSK23 functions as a negative regulator of BR signaling, acting synergistically with factors such as OsPPKL1 and OsBSK3 [26,27]. OsSK22 serves as the central kinase in BR signaling, regulating traits such as plant architecture, panicle shape, and disease resistance through the phosphorylation of various downstream factors [28,29,30,31]. Additionally, it interacts with pathways involving jasmonic acid, strigolactone, and others [32,33,34]. OsSK41 regulates rice grain shape and weight via the auxin pathway and is associated with salt stress resistance [35,36,37,38]. Despite their evolutionary conservation, the biological roles of GSK3 genes can vary among species and contexts. To date, no comprehensive study has explored the GSK3 gene family in watermelon, limiting our understanding of its potential roles in fruit development and abiotic stress tolerance.
Given the crucial functions of GSK3 genes, we hypothesize that the GSK3 gene family in watermelon is evolutionarily conserved and plays analogous roles in mediating fruit development, plant architecture, and tolerance to abiotic stresses by integrating hormone signaling pathways, particularly BR signaling. The aim of this study is to conduct a genome-wide identification and systematic characterization of the ClGSK3 gene family. By achieving this aim, we seek to provide the first comprehensive overview of the ClGSK3 family, clarify their potential functional roles in watermelon, and identify key molecular targets that can be utilized in breeding programs to develop climate-resilient and high-quality watermelon varieties. This work lays a foundation for further functional validation of ClGSK3 genes and their practical application in watermelon genetic improvement.
2. Results
2.1. Identification of GSK3/Shaggy-like Family Members in Watermelon
To identify GSK3/Shaggy-like family genes in watermelon, we conducted BLAST (https://blast.ncbi.nlm.nih.gov/, accessed on 3 November 2025) searches using the protein sequences of all 10 members from Arabidopsis and 9 members from rice as queries. To ensure the reliability of the BLAST results, we obtained the kinase domain model Pkinase (PF00069.23) and the hidden Markov model (HMM) of AtSKs protein kinase from the Pfam website. We then employed these models to further search for and confirm the members of the GSK3 family in watermelon. We identified a total of eight members, which we designated as ClSK11, ClSK12, ClSK13, ClSK21, ClSK22, ClSK23, ClSK31, and ClSK41, based on their closest phylogenetic relationships with Arabidopsis and the rice family (Table S1).
Further analysis of GSK3 proteins entailed evaluating their physical and chemical properties. Predicted amino acid counts varied from 379 (ClSK21) to 469 (ClSK31), corresponding to molecular weights ranging from 43.03 kDa (ClSK21) to 52.75 kDa (ClSK31) (Table S2). The predicted isoelectric point (pI) suggested that, apart from ClSK31 (pI = 6.61), all family members were expected to be basic proteins with pI values above 7. Moreover, all the 8 proteins displayed an average grand average of hydropathy (GRAVY) score below 0, indicating their hydrophilic nature. Examination of the instability index indicated that ClSK21, ClSK22, and ClSK23 proteins were anticipated to be unstable, with instability index values exceeding 40 (Table S2).
2.2. Structural and Phylogenetic Analysis
To delve into the functional differentiation and evolutionary path of the watermelon GSK3 gene family, we initially generated a maximum likelihood (ML) phylogenetic tree using IQ-TREE software (v2.4.0) based on full-length amino acid sequences (Figure 1a). Subsequently, we delineated the conserved protein domains (Figure 1b) and the exon-intron architecture of the genes (Figure 1c).
Subsequently, an ML phylogenetic tree encompassing 154 GSK3 proteins from 13 species, including Arabidopsis, rice, wheat and maize (Table S2), was constructed using full-length sequences in IQ-TREE software. The analysis unveiled four distinct clusters (I–IV), each containing members from the three species, indicating an early differentiation before speciation. Notably, Group I included ClSK11/12/13, Group II contained ClSK21/22/23, Group III had ClSK31, and Group IV included ClSK41 (Figure 2).
Our findings indicated that the phylogenetic distinct clusters of ClGSK3s correlate with both protein structure and gene structure. At the protein level, while each member contains a single characteristic Pkinase domain, the domain composition and length among members of each group are highly consistent (Figure 1b, Figure 2 and Figure S1). At the gene level, the number of exons ranged from 11 to 13: Group I genes had 12 exons, Group II had 11 exons (except for ClSK22), with substantial diversity in intron phase and length, which might reflect ancestral indel events or possible transcript isoforms. This “phylogenetic-structural” association pattern implies that post-gene duplication, structural reconfiguration, and functional diversification may follow a progressive trajectory of “initial conservation followed by subsequent diversification.” These insights offer pivotal clues for future functional validation and molecular engineering endeavors.
2.3. Chromosomal Localization and Collinearity Analysis
To ascertain the chromosomal localization of GSK3 genes in watermelon, we conducted an analysis of their physical distribution across chromosomes. Among the 8 GSK3 genes, they are distributed evenly across 7 of the 11 watermelon chromosomes. Chromosome 10 contains 2 genes, while chromosomes 1, 2, 3, 4, 6, and 7 each harbor 1 gene (Figure 3a).
Gene duplication, including whole-genome duplication (WGD), segment duplication, and tandem duplication, is widespread in plant genomes and is a significant factor in genome evolution, resulting in the substantial enlargement of plant gene families [39]. A collinearity analysis of GSK3 genes in Arabidopsis, rice, and watermelon was performed to clarify the evolutionary mechanism of the GSK3 family. The analysis identified 4 GSK3 genes in rice, 4 in watermelon, and 6 homologous genes in Arabidopsis, resulting in the formation of 10 collinear pairs (Figure 3b, Table S3), which suggests a shared ancestry among these genes.
The synonymous substitution rate (Ks) of all colinear gene pairs was estimated with the MCScanX module implemented in TBtools. Among the GSK3 pairs, the four Arabidopsis–watermelon colinear fragments exhibited Ks < 1, whereas the six watermelon–rice fragments yielded Ks > 1 (Table S3). This distribution indicates a closer evolutionary distance between watermelon and the dicot model Arabidopsis than between watermelon and the monocot rice, consistent with the divergence of dicotyledonous and monocotyledonous lineages. Importantly, all colinear GSK3 gene pairs displayed Ka/Ks ratios significantly < 1, reflecting strong purifying selection that has acted to conserve GSK3 function across these taxa.
The Orthogroup assignment was derived from OrthoFinder (v2.5, Emms DM et al., United Kingdom) default settings, grouping the 10 collinear genes into 3 orthogroups (Table S4). These 10 pairs show a mix of one-to-one and one-to-many orthology (Table S4), suggesting ancient duplications within the watermelon lineage after the monocot–dicot split.
2.4. Promoter Cis-Regulatory Elements Analysis
Regulation of plant gene transcription relies on controlling cis-regulatory elements (CREs) within promoter regions, influencing gene expression in response to environmental stresses and tissue-specific contexts. To explore the transcriptional regulatory network of the GSK3 gene family in watermelon, we analyzed the 2000 bp upstream promoter sequence and predicted CREs using the PlantCARE tool. Our analysis identified enriched core elements related to hormone, light, anaerobism, drought, heat, wound, and cold signaling pathways in the promoter region of the GSK3 gene family (Figure 4, Table S5), suggesting a key role in integrating light signals, hormone cues, and stress responses.
Group I (ClSK11/12/13) encompasses the most diverse array of elements, comprising 55 types, and includes numerous photoresponsive elements and stress-related elements. This suggests a robust capacity for stress adaptation and potential involvement in light-dependent processes, such as photomorphogenesis and the regulation of photosynthesis in plants. Group II (ClSK21/22/23) exhibited a high percentage of abundant hormone-responsive elements (35.2%) and photoresponsive elements (27.8%). These elements potentially play a role in light-dependent mechanisms, such as regulating flowering time. Group III (ClSK31) comprises an equal proportion of photoresponsive and hormone-responsive elements, each accounting for 33.3%. This group contains fewer element types, totaling 18, and does not include complex stress-responsive elements. It may play a role in regulating specific tissues or developmental stages, as well as in hormone regulation under particular photoperiods. Group IV (ClSK41) has the highest proportion of hormone-responsive elements (46.2%), includes unique cold-responsive elements (LTR), and possesses the smallest total number of elements (26). This group may play a crucial role in sustaining growth in low-temperature environments.
2.5. Tissue-Specific Expression of ClGSK3s in Watermelon
The temporal and spatial expression of genes is closely linked to the manifestation of biological functions. In this study, ClGSK3 genes were analyzed using the transcriptome database (Accession no. PRJNA676179) of watermelon across various tissues and developmental stages, including vegetative tissues (3-week-old leaves, roots, and stems), flower organs (bisexual and male flowers at anthesis), and fruit developmental series from 5 to 35 days after pollination [40]. The analysis revealed a generally consistent expression pattern across different tissues (Figure 5, Table S6). ClSK41 exhibited a pronounced specificity for fruit, particularly in mature pulp, suggesting its potential involvement in glucose metabolism and accumulation. ClSK23 demonstrated high expression during the mid-stage of fruit development, indicating a possible role in development and sugar transport. ClSK22/21 showed elevated expression in the pericarp and tendrils, which may relate to plant morphology, fruit shape, and stress responses. ClSK12/13/31 were predominantly expressed in floral organs, potentially linked to pollen development or sex determination. ClSK11 was highly expressed in both flowers and roots, suggesting its involvement in basal energy metabolism.
We re-analyzed the published RNA-seq dataset (SRP143549) of four-leaf-stage drought-tolerant M20 and susceptible Y34 watermelon seedlings under 4 and 8 days of water-withholding (drought) treatment. Relative to fully irrigated controls, ClSK12 and ClSK41 were markedly up-regulated in both genotypes at both time points, pointing to a conserved, early role in drought perception. By contrast, ClSK11 and ClSK21 exhibited pronounced up-regulation only at 8 d, while ClSK13 was significantly down-regulated at the same stage in both genotypes (Figure S2, Table S7), implying that these three genes participate in later-acting drought responses. ClSK31 was constitutively higher in M20, suggesting a basal role in drought resilience. Then we reanalyzed the published RNA-seq dataset (GSE146087) obtained from six-week-old seedlings of the Crimson Sweet variety exposed to 300 mM NaCl [41]. Compared to deionized-water controls, all ClGSK3s, except for ClSK31, exhibited significant differential expression (Figure S3, Table S8), indicating their involvement in the salt-stress response.
2.6. Prediction and Validation of the Interacting Proteins of the ClGSK3s
Protein–protein interactions (PPIs) involving GSK3 proteins were examined to elucidate potential functions, signaling pathways, and metabolic processes. The STRING database facilitated the prediction of interactions among the eight ClGSK3 proteins, utilizing ortholog information primarily derived from Arabidopsis. A total of 53 potential interacting partners with five ClGSK3 proteins were identified (Figure 6a, Table S9). Among them, ClSK21 was predicted to interact with 23 proteins, ClSK31 with 9 proteins, ClSK41 with 5 proteins, ClSK13 with 4 proteins, ClSK22 with 3 proteins, and ClSK12 with 2 proteins (Figure 6a). KEGG analysis of these interacting proteins revealed their involvement in multiple metabolic pathways, including ribosome biogenesis and circadian rhythm, with the plant hormone signaling pathway containing the largest number of members (Table S9). This finding suggests a close relationship between the ClGSK3 protein and plant hormone signaling, particularly in the case of ClSK21, which has 16 of its 23 predicted interacting proteins associated with this pathway.
Only ClSK21 exhibited high-confidence interactions (Required score > 0.900) with six proteins: Cla97C05G101410, Cla97C06G121690, Cla97C08G152340, Cla97C01G018680, Cla97C10G202040, and Cla97C08G158860 (Table S9). These high-confidence interactions were validated through yeast two-hybrid experiments. The results indicate that ClSK21 has genuine interactions with Cla97C05G101410, Cla97C01G018680, and Cla97C08G158860 (Figure 6b). These three genes encode orthologs of the key BR signaling factors BKI1 (Cla97C05G101410) and BZR1 (Cla97C01G018680 and Cla97C08G158860) in watermelon. These findings suggest that ClSK21 may play a regulatory role in BR signaling within this species. Further research is necessary to provide additional evidence.
2.7. Expression Pattern of GSK3s Response to BR Treatment
Promoter profiling, spatiotemporal expression atlases and predicted interactors all pointed to a BR link for ClGSK3s. To test this, we monitored the transcript abundance of all eight family members after 0 and 1 µM exogenous brassinolide (BL), a type of BR, treatment. Within the same time window, ClSK21 and ClSK31 transcripts rose 2.57- and 2.65-fold, respectively (p < 0.05), whereas ClSK41 dropped to 85.8% of the control level (p < 0.01) (Figure 7). These reciprocal responses implicate ClSK21, ClSK31 and ClSK41, which may participate in BR signaling.
3. Discussion
Glycogen synthase kinase 3/Shaggy-like kinases are highly conserved serine/threonine kinases that regulate plant growth, development, and stress responses through the integration of hormone signaling [10,11,12]. Prior to this study, the GSK3 gene family in watermelon, an economically significant horticultural crop, had not been characterized. In this research, we systematically analyzed the identification, phylogeny, chromosomal distribution, cis-regulatory elements, transcriptome data, protein–protein interactions, and BR responsiveness of ClGSK3s. This work provides the first comprehensive insight into their potential functions and evolutionary dynamics.
Watermelon possesses only eight GSK3 genes, fewer than most model plants and crops reported to date [10,42,43,44,45,46,47]. Nevertheless, all four canonical sub-clades (I–IV) are represented, suggesting a “minimal but complete” GSK3 toolkit shaped by dicot-specific duplication and strong purifying selection. The evolutionary trajectory of the GSK3 family was systematically examined by identifying all family members in 13 species (Table S2), as documented in previous studies [10,42,43,44,45,46,47]. Phylogenetic clustering of ClGSK3s into four subfamilies, each encompassing members from all examined species (Figure 2), indicates that the major lineages diverged prior to the monocot–dicot split and have since maintained structural conservation. Synteny and Ks analyses identify ten collinear ClGSK3–AtGSK3/OsGSK3 pairs (Table S3); dicot–dicot blocks exhibit Ks < 1, while dicot–monocot blocks present Ks > 1, thereby calibrating the evolutionary timeline of Cucurbitales. All collinear pairs exhibit Ka/Ks ≪ 1 (Table S3), indicating persistent purifying selection that maintains core kinase function. One-to-one and one-to-many orthologies trace the modern ClGSK3s complement to (i) ancient pre-eudicot duplications shared with Arabidopsis and (ii) a watermelon-specific tandem duplication event on chromosome 10, generating two paralogs (ClSK11 and ClSK13). This duplication is absent in Arabidopsis and rice, indicating it occurred after the Cucurbitaceae lineage diverged from other dicots. Each ClGSK3 contains the Pkinase domain signature, and closely related genes exhibit similar exon–intron architecture and domain layout (Figure 1). Subfamily members share highly consistent domain composition and length at the protein level. At the gene level, exons vary from 11 to 13 (Group I comprises 12 exons, while Group II mostly comprises 11 exons except ClSK22). The diversity in intron phase and length may indicate ancestral indel events or the presence of transcript isoforms, underscoring a “phylogenetic–structural” correlation in which post-duplication rearrangements facilitate functional differentiation. Most proteins are basic and hydrophilic, making them suitable for cytoplasmic signaling, whereas ClSK21/22/23 show high instability indices (>40) (Table S2), facilitating rapid hormonal or stress responses through conformational switching or targeted degradation.
Watermelon is distinguished by its high sugar content and large fruit size [2,48,49], and our data identify ClGSK3s as key regulators of these species-specific traits. Tissue profiles and promoter dissection converge to support a model in which ClGSK3s connect hormonal signals to developmental outcomes and fruit quality, with distinct functional specializations aligned to their subfamily-specific cis-acting element features. Group I (ClSK11/12/13) harbors the most diverse cis-elements and a high abundance of light-responsive elements (Figure 4, Table S5), endowing this subgroup with robust stress adaptation capacity and potential involvement in light-dependent processes. Consistently, ClSK12/13 are predominantly expressed in floral organs, while ClSK11 shows high expression in both flowers and roots (Figure 5, Table S6), suggesting their coordinated roles in pollen maturation, sex determination, and basal energy metabolism. Group II (ClSK21/22/23) exhibits a high proportion of abundant hormone-responsive elements and photoresponsive elements (Figure 4, Table S5), which may underpin their function in light-dependent developmental processes. ClSK21/22 are highly expressed in the pericarp and tendrils (Figure 5, Table S6), with their auxin-responsive AuxRR-core and light-responsive elements reflecting a conserved role in modulating plant architecture and longitudinal growth analogous to OsSK21/22 in rice [50,51,52], while ClSK23 is upregulated during mid-fruit development (Figure 5, Table S6), coinciding with maximal cell expansion and sucrose import, implying involvement in fruit development and sugar transport. Group III (ClSK31) features equal proportions of photoresponsive and hormone-responsive elements with fewer total element types and no complex stress-responsive motifs (Figure 4, Table S5), suggesting specialization in regulating specific tissues or developmental stages under particular photoperiods; its floral-restricted expression (Figure 5, Table S6) further supports potential roles in pollen development or sex determination, alongside auxin, gibberellin, and ethylene response elements. Group IV (ClSK41) has the highest proportion of hormone-responsive elements and unique cold-responsive LTR elements (Figure 4, Table S5), indicating a key role in mediating low-temperature adaptation. Notably, ClSK41 exhibits pronounced fruit-specific expression, especially in ripe, sugar-rich pulp (Figure 5, Table S6), paralleling known carbohydrate regulators such as ClVST2, ClSWEET3, and ClAGA2 [48,53], which suggests that GSK3-mediated phosphorylation contributes to terminal sugar loading in watermelon fruits. The unique LTR element further facilitates fruit ripening and sugar accumulation under low temperatures. The validation of predicted interactions identified 53 potential interacting partners with five ClGSK3 proteins; excluding unannotated hits, 21 of the 29 functionally annotated interactors are linked to hormone pathways (Table S9). Among these, ClSK21 interacts with the watermelon orthologs of BR-signaling transcription factor BZR1 (Figure 6b and Figure S4), recapitulating the canonical BIN2/AtSK21 interactions characterized in Arabidopsis [54,55,56]. Notably, ClSK21 interacts with the BKI1 ortholog (Figure 6b and Figure S4), a conserved component of BR signaling, which differs from the previously reported signaling networks in Arabidopsis and rice [54,55,56]. This finding highlights the unique characteristics of BR signaling in watermelon and offers a significant advancement for future research, providing a new entry point for exploring BR-mediated regulatory pathways in this crop. qRT-PCR analysis demonstrates that ClSK21/31/41 transcripts shift significantly following BL treatment (Figure 7), confirming the involvement of these kinases in BR signaling [16,21,54]. Given that BR promotes fruit enlargement, sugar allocation, stem elongation, and stress tolerance [57,58], the cloned ClGSK3s—particularly the BR-responsive triad—provide valuable gene targets for marker-assisted selection aimed at optimizing plant architecture and enhancing soluble solids in watermelon lines.
Promoter dissection, tissue profiles, and validation of predicted interactions of the ClGSK3 family converge to implicate the potential roles in stress tolerance. To systematically dissect the abiotic stress roles of the ClGSK3s, we re-mined the published RNA-seq dataset (SRP143549) (Figure S2), profiling four-leaf-stage seedlings of drought-tolerant M20 and drought-susceptible Y34 under 4 d and 8 d water-withholding [59]. The results showed conserved and stage-specific drought response patterns of ClGSK3s, such as early up-regulation of ClSK12/41 and late response of ClSK11/21/13. Then we reanalyzed the published RNA-seq dataset (GSE146087) (Figure S3) obtained from six-week-old seedlings of the Crimson Sweet variety exposed to 300 mM NaCl [39], revealing differential expression of ClGSK3s under salt stress. This finding is consistent with the established functions of GSK3 proteins in other plant species (e.g., AtSK11/12 in Arabidopsis for drought tolerance [60], AtSK31 for salt tolerance [23], and OsSK21 in rice for multi-stress tolerance [24,25]). Evolutionary conservation suggests that ClGSK3s may play analogous roles in stress response. The interaction of ClSK21 with BR signaling components further substantiates its involvement in stress tolerance, as BR signaling enhances resistance to stress through the regulation of stress-responsive genes; for instance, rice GSK3-regulated BR signaling promotes submergence tolerance [61]. The expression of ClSK21 and ClSK22 in pericarp and tendril tissues, which serve as the first line of stress defense, reinforces this potential.
To enhance the understanding of the functional mechanisms of ClGSK3s in watermelon and to broaden their applications in breeding, the following targeted recommendations are proposed: First, verify the specific roles of key genes (ClSK21, ClSK31, ClSK41, etc.) in BR signaling, fruit development, and stress tolerance through CRISPR-Cas9 knockout/overexpression techniques, along with phenotypic and physiological analyses of transgenic plants. Second, identify phosphorylation substrates of core proteins, particularly ClSK21, utilizing immunoprecipitation-mass spectrometry (IP-MS) and construct downstream signaling networks via transcriptomic and metabolomic analyses. Third, investigate the response patterns of ClGSK3s to multiple hormones, such as auxin and jasmonic acid, and elucidate their crosstalk mechanisms to clarify the integration of hormone signals. Fourth, evaluate the breeding potential of these genes through field trials aimed at developing varieties with optimized plant architecture, improved fruit quality, and enhanced stress resistance using genetic engineering or marker-assisted selection. Finally, explore the regulatory roles of ClGSK3s under combined stresses to facilitate the breeding of climate-resilient varieties.
4. Materials and Methods
4.1. Genome-Wide Identification of GSK3 Genes in Watermelon
To identify GSK3 family genes in watermelon, we employed the protein sequences of GSK3s from Arabidopsis and rice as queries to search the watermelon reference genome (version: 97103_v2 Genome) protein database on NCBI (https://www.ncbi.nlm.nih.gov/) using BLASTp with specific criteria: E-value ≤ 1 × 10^−10^ and identity ≥ 40% (accessed on 3 November 2025). Subsequently, we utilized the kinase domain model Pkinase (PF00069) and the hidden Markov model (HMM) of AtSKs protein kinase from the InterPro database (https://www.ebi.ac.uk/interpro/, accessed on 3 November 2025) to search and confirm watermelon GSK3 family members. The protein sequences obtained from both methods were merged, and the conserved domains were validated using SMART (http://smart.embl-heidelberg.de/, accessed on 3 November 2025) and PFAM (http://pfam.janelia.org/, accessed on 3 November 2025). Candidate genes were selected after removing redundancies, and following the nomenclature of Arabidopsis, we standardized the names of the identified watermelon GSK3s based on their respective subfamilies and position on the phylogenetic tree. The sequence alignment was generated utilizing the clustalw platform (https://www.genome.jp/tools-bin/clustalw, accessed on 5 November 2025) and enhanced with espript 3 (https://espript.ibcp.fr/ESPript/ESPript/, accessed on 5 November 2025).
4.2. Chromosomal Localization and Collinearity Analysis
The chromosomal localization of GSK3 genes was analyzed by retrieving their loci information from the annotation gff3 files (version: 97103_v2 Genome). The chromosomal positions of GSK3 genes and their collinearity across watermelon, Arabidopsis, and rice were visualized through the functions ‘Gene Location Visualize’ and ‘Dual Synteny Plot for MCScanX’ with default parameters in TBtools (v2.322, Chen C et al., China) [62].
4.3. Phylogenetic Tree Analysis
Phylogenetic trees were constructed using IQ-TREE software (v2.4.0, Minh BQ et al., International collaboration (led by researchers from Denmark, Germany, Australia, etc.)) with maximum likelihood (ML) [63]. Sequences of GSK3 family proteins were retrieved from previous studies [10,42,43,44,45,46,47] and aligned using the MAFFT online tool (https://www.ebi.ac.uk/Tools/msa/mafft/, accessed on 12 January 2026) with default parameters. The aligned sequences were utilized as input with the following settings: Q.plant+G4 model, 1000 bootstrap replicates, and 1000 SH-aLRT tests for branch support. The phylogenetic tree was enhanced using the online tool iTOL (https://itol.embl.de/, accessed on 12 January 2026).
4.4. Expression Profiles of ClGSK3 Genes
Transcriptome data from previous investigations were obtained from the NCBI database. The raw sequencing reads were processed to obtain high-quality clean reads, which were then re-mapped to the updated watermelon reference genome (97103v2). Subsequently, gene expression levels were normalized (TPM), converted to log2 ratios, and visualized using TBtools software (v2.322, Chen C et al., China).
4.5. Cis-Acting Element Analysis of GSK3 Genes in Watermelon
The 2000 bp upstream sequence information of the GSK3 genes was retrieved from the gff3 annotation file for analysis of cis-acting elements in the gene promoters. An analysis was conducted utilizing the online tool PlantCARE with default parameters (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 19 November 2025), and the outcomes were visualized through the ‘simple biosequence viewer’ function of TBtools (v2.322) with default parameters.
4.6. PPI Network Prediction
To identify potential proteins that may interact with GSK3 family proteins in watermelon, we obtained protein–protein interaction data for Arabidopsis from the STRING database (https://string-db.org/, accessed on 1 December 2025), applying a medium confidence threshold (Required score > 0.700). A BLASTP search of the watermelon protein database was then conducted using the interaction proteins from Arabidopsis via the ‘PPI Prediction’ feature in TBtools. The results were subsequently visualized using Cytoscape software (v3.10.3, Cytoscape Consortium, San Diego, CA, United States).
4.7. Y2H Assays
For the GAL4-based Y2H assay, the CDS of ClSK21 and the six high-confidence interacting proteins were cloned into the pGADT7 vector at the EcoRI and BamHI restriction sites through gene synthesis by GenScript Corporation (Nanjing, China). The gene-synthesized fragments were subsequently excised from pGADT7 with EcoRI and BamHI and ligated into the matching sites of pGBKT7 using T4 DNA ligase. The interaction assays were conducted with a Yeast Transformation Kit (YT0010, Beijing LABLEAD Trading Co., Ltd., Beijing, China) following the manufacturer’s instructions. The DMSO (ID9011) used in the experiment was purchased from Beijing Solarbio Science & Technology Co., Ltd. (Beijing, China). Both the bait and prey constructs were co-transformed into the yeast strain AH109 and cultured on SD/-Leu/-Trp medium, followed by selection on SD/-Leu/-Trp/-His/-Ade medium.
4.8. Quantitative Real-Time PCR
The homozygous inbred line TP101 was used in the exogenous BR treatment experiment, which was established by the Institute of Cash Crops, Hebei Academy of Agriculture and Forestry Sciences. The exogenous BL treatment experiment was performed as described previously [5]. The 3-day-old hydroponically grown seedlings were treated with 1 µM exogenous BR or a solvent-only control (0 µM BR). After 4 days of treatment, total RNA was isolated from all seedlings of both treated and control groups (three biological replicates each) using Trizol reagent (15596026CN, Invitrogen, Carlsbad, CA, USA), and the quality was checked by agarose (CA1341, Beijing Coolaber Science & Technology Co., Ltd., Beijing, China) gel electrophoresis. Subsequently, cDNA was generated with a QuantiTect Reverse Transcription Kit (JKR23014, GeneCreate, Wuhan, China) and employed as a template for quantitative real-time PCR (qRT-PCR) analysis. qRT-PCR assays were conducted on an Applied Biosystems 7500 real-time PCR system, employing the 2 × SYBR Green qPCR Mix (AH0105-B, Shandong Sparkjade Biotechnology Co., Ltd., Jinan, China), following the manufacturer’s recommended protocol. Each trial was replicated three times using distinct biological samples. Expression levels were normalized to the internal control gene ClCAT [64] using the 2^(−ΔΔCt)^ method, while all quantitative primers were listed in Table S10.
5. Conclusions
This study presents the first comprehensive identification and characterization of the watermelon GSK3 gene family, which consists of eight evolutionarily conserved members. ClGSK3s integrate BR and other hormone signaling pathways to regulate watermelon fruit development, plant architecture, and abiotic stress tolerance. Notably, ClSK21 serves as a key regulator of BR signaling through its interactions with BKI1/BZR1 orthologs. These findings enhance the understanding of the diversity within the plant GSK3 family and provide prioritized molecular targets for breeding high-quality, stress-resistant watermelon varieties, thereby establishing a foundation for further functional research and genetic improvement.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yu Y. Guo S. Ren Y. Zhang J. Li M. Tian S. Wang J. Sun H. Zuo Y. Chen Y. Quantitative Transcriptomic and Proteomic Analysis of Fruit Development and Ripening in Watermelon (Citrullus lanatus)Front. Plant Sci.20221381839210.3389/fpls.2022.81839235392508 PMC 8980866 · doi ↗ · pubmed ↗
- 2Guo S. Zhao S. Sun H. Wang X. Wu S. Lin T. Ren Y. Gao L. Deng Y. Zhang J. Resequencing of 414 cultivated and wild watermelon accessions identifies selection for fruit quality traits Nat. Genet.2019511616162310.1038/s 41588-019-0518-431676863 · doi ↗ · pubmed ↗
- 3Wang Y. Wang J. Guo S. Tian S. Zhang J. Ren Y. Li M. Gong G. Zhang H. Xu Y. CRISPR/Cas 9-mediated mutagenesis of Cl BG 1 decreased seed size and promoted seed germination in watermelon Hortic. Res.202187010.1038/s 41438-021-00506-133790265 PMC 8012358 · doi ↗ · pubmed ↗
- 4FAO Food and Agriculture Organization of the United Nations Agriculture Databases 2023 Available online: https://www.fao.org/faostat/zh/#data/QCL/visualize(accessed on 3 December 2025)
- 5Tian P. Zhang L. Zhang J. Liu B. Liu W. Li B. Gao X. Zhang J. Wu Y. Xu Y. Genome-Wide Identification of GW 5-LIKE Family Revealed the Function of Cl GL 1 Involved in Fruit and Seed Shape by Mediating Brassinosteroid Signaling in Watermelon Horticulturae 202511132610.3390/horticulturae 11111326 · doi ↗
- 6Kaur S. Oliveira M.M. Kaundal A. Understanding Salt Stress in Watermelon: Impacts on Plant Performance, Adaptive Solutions, and Future Prospects Int. J. Plant Biol.2025169310.3390/ijpb 16030093 · doi ↗
- 7Niu H. Tan J. Yan W. Liu D. Yang L. Advances in Functional Genomics for Watermelon and Melon Breeding: Current Progress and Future Perspectives Horticulturae 202511110010.3390/horticulturae 11091100 · doi ↗
- 8Frame S. Cohen P. GSK 3 takes centre stage more than 20 years after its discovery Biochem. J.200135911610.1042/bj 359000111563964 PMC 1222116 · doi ↗ · pubmed ↗