Interannual Variation in Seed Traits of Cedrela Species: Implications for Conservation in the Context of Climate Change
Guadalupe Galíndez, Ana Álvarez, Diana Ceccato, Victoria Rivero, Gisela Malagrina, Tania Bertuzzi, Pablo Saravia, Stavros Nicolás Sola, Carol C. Baskin, Luis Fornes

TL;DR
This study examines how climate change affects seed traits and storage potential of two endangered Cedrela species in Argentina, highlighting the importance of seed banking for conservation.
Contribution
The study reveals species-specific responses of seed longevity and traits to climate variables, emphasizing ex situ conservation strategies under climate change.
Findings
Cedrela balansae seeds are medium-lived and suitable for long-term ex situ conservation.
Cedrela fissilis seeds are short-lived and highly sensitive to storage conditions.
Future warming and drying may reduce germination windows, with species-specific impacts.
Abstract
Climate change is altering temperature and precipitation regimes in Argentina, with potential consequences for regeneration and persistence of forest tree species, emphasizing the importance of ex situ seed conservation. We evaluated interannual variation in seed traits, desiccation tolerance, storage behavior, and longevity of Cedrela balansae C. DC. and C. fissilis Vell. (Meliaceae), two endangered native species of subtropical rainforests in Argentina. Both species produced desiccation-tolerant seeds, independently of collection year, seed traits, or climatic conditions. Depending on the species, seed traits and longevity varied across years and showed strong relationships with temperature and precipitation, particularly during seed development. Cedrela balansae seeds are medium-lived seeds and have high longevity under standard seed banking conditions, suggesting strong potential…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
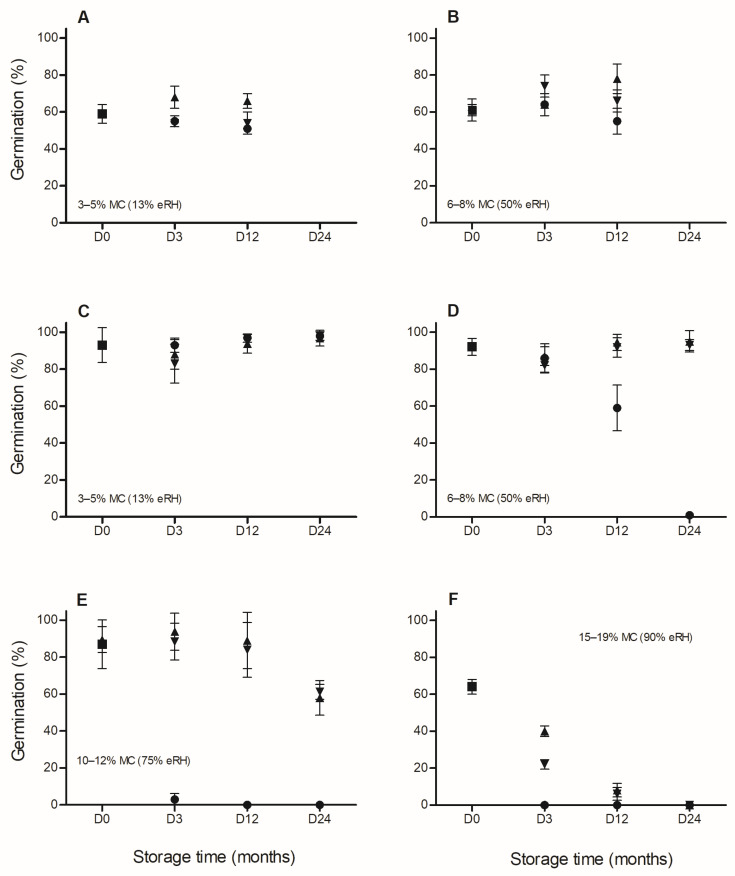 Figure 1
Figure 1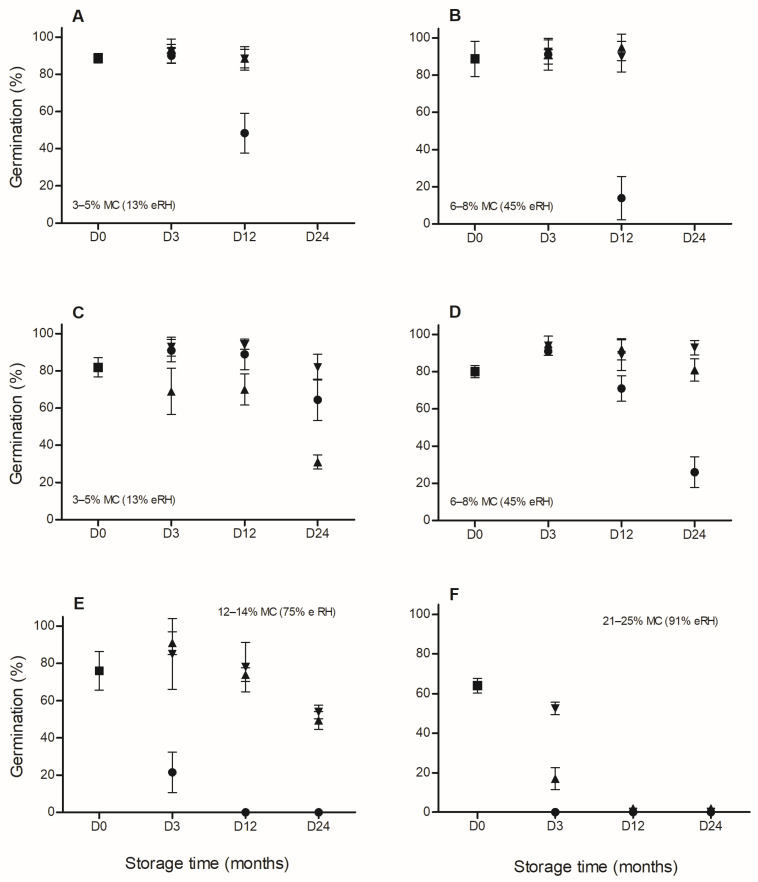 Figure 2
Figure 2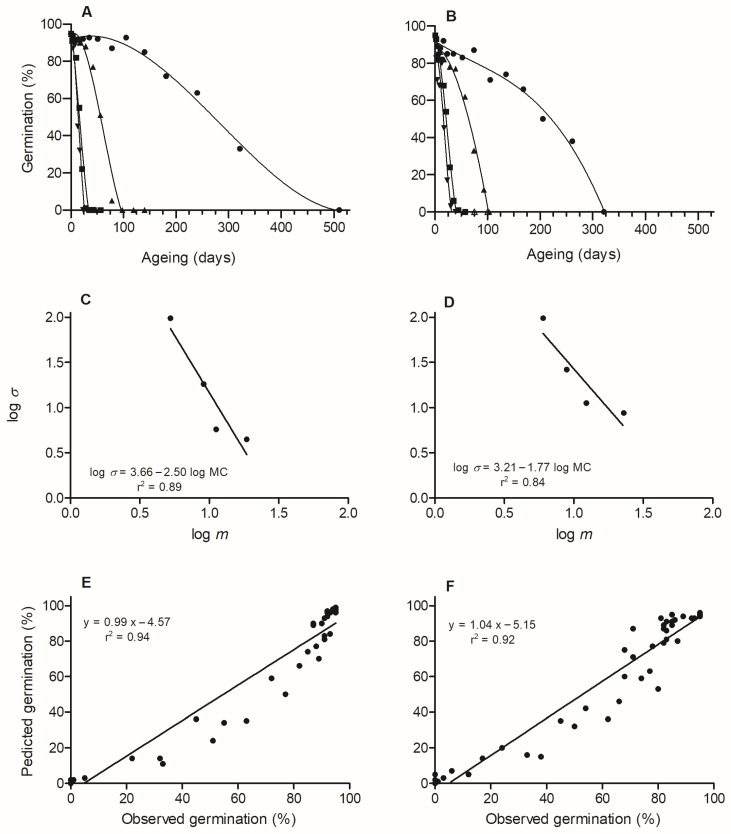 Figure 3
Figure 3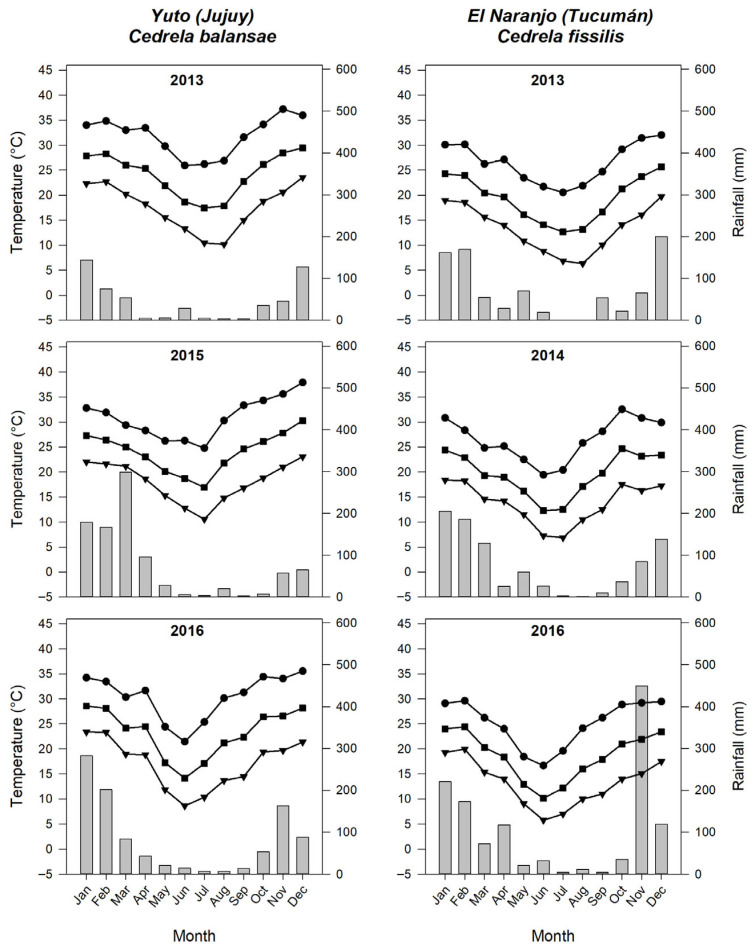 Figure 4
Figure 4- —National Institute of Agricultural Technology (INTA) projects
- —El Consejo de Investigación de la Universidad Nacional de Salta project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSeed Germination and Physiology · Ecology and Vegetation Dynamics Studies · Amazonian Archaeology and Ethnohistory
1. Introduction
Global climate change is affecting plant regeneration from seeds, which may compromise population recruitment and, in the long term, threaten species persistence [1,2]. In Argentina, recent decades have exhibited significant trends in temperature extremes, more frequent heatwaves, increased precipitation, and severe droughts [3]. These climatic changes have, among other consequences, driven substantial variations in forest dynamics and reductions in mean annual net primary productivity [4]. In addition, nearly three million hectares of forest have been lost, positioning Argentina among the ten countries with the highest rates of recent forest loss [4]. This substantial reduction in forest area has been primarily driven by land use change, particularly the expansion of agriculture and grazing, and anthropogenic fires. As a result, the diversity of native forests has declined, and numerous species of high economic value have suffered genetic erosion, local extinction, or are considered endangered or threatened at regional and/or global scales [5,6]. These cumulative threats emphasize the importance of coordinated actions for storing seeds of forest species to guarantee their use in breeding, plant propagation, and ecological restoration of ecosystems [7].
Ex situ seed conservation in germplasm or seed banks is widely recognized as the most efficient and cost-effective strategy for long-term preservation of seeds of many plant species, since it safeguards genetically and geographically representative samples for future use [8,9]. To achieve effective conservation, it is essential to determine seed desiccation tolerance, storage behavior, and longevity [10]. Based on seed desiccation tolerance (DT) and storage behavior, seeds are classified into three categories: orthodox, where seeds can tolerate drying to a moisture content (MC) of approximately 3–7% and subzero temperature storage (−20 °C); recalcitrant, where seeds are sensitive to both desiccation and subzero temperatures; and intermediate, where seeds are sensitive to drying below 10% MC and/or to subzero temperatures [11]. DT has been associated with ecological, physiological, and morphological seed traits [12,13,14,15,16]. Tropical species are more likely to produce desiccation-sensitive seeds than temperate ones, and species with large seeds, thin coats, and high MC at dispersal time are more prone to desiccation than small seeds with thick coats and low MC. However, this classification is based on seed behavior at full maturity, before germination begins, and for many species, the transition from maturation to germination is not clearly defined, complicating seed classification [17]. In this sense, Walters [18] proposed viewing seed responses to desiccation as a continuum—from fully desiccation-tolerant to highly desiccation-sensitive—rather than as discrete categories, highlighting the need for quantitative evaluation of each seed lot.
Seed longevity is defined as the period during which seeds maintain viability at or above a target threshold under air-dry storage conditions [19]. In orthodox seeds, longevity increases as both temperature and seed MC decrease [20]. Consequently, long-term storage of orthodox seeds under low MC and subzero temperatures is the most widely used method for ex situ conservation of plant genetic resources. To estimate and to compare seed longevity in seed banks, accelerated (controlled) ageing tests—conducted at high MC and temperature—are commonly used [21,22,23,24]. Thus, loss of seed viability during accelerated aging or storage can be predicted by the seed viability equations [20]:
where v is viability (in normal equivalent deviates, NED) of the seed lot after p days in storage, K_i_ is the initial viability (NED) of the seed lots, σ is the time (d) for viability to decrease by 1 NED (i.e., the standard deviation of the normal distribution of seed deaths over time), and σ^−1^ is the slope of the transformed survival curve.
The effects of temperature (t) and MC (m) on seed longevity are species-specific, according to the following equation:
where K_E_ is a species-specific constant for inherent seed longevity; C_W_ is species-specific
MC quantifies the logarithmic relationship between seed longevity (σ) and seed MC (m); C_H_ and C_Q_ are species-specific temperature constants for the quadratic relationship between seed longevity and storage temperature, t, respectively.
Combining Equations (1) and (2) produces the full viability model, i.e., Equation (3):
Considering that the effects of temperature on longevity appear to be the same for all species, C_H_ and C_Q_, take universal values of 0.0329 and 0.000478, respectively [25].
Seed longevity also has been linked to the climate of the area where seeds of the species were collected as well as to seed traits [9,24]. In general, seeds from species inhabiting warmer and drier regions exhibit greater longevity than those from cooler and wetter environments, and small-seeded and non-oil-rich species tend to show longer lifespan than large-seeded or oil-seeded species [15,16,24,26]. Nevertheless, correlations between seed traits and longevity are not consistent across all species [9,24]. In addition, intraspecific and interannual variation in seed longevity have been associated with local environmental conditions during seed development [27,28,29,30]. For example, White [30] showed that accessions collected from the same alpine population for several years differed in longevity, with seeds developed in wetter years being shorter-lived than those from drier years. However, few studies have investigated seed DT, storage behavior, and longevity in native tree species, probably due to their inherent longevity and responses to MC and temperature being less uniform than in crops [26].
The genus Cedrela (Meliaceae) is distributed from Mexico (24° N) to Argentina (28° S) and comprises tropical and subtropical species [31], being a protected genus [5]. In Argentina, four Cedrela species have been reported, all occurring in the northwestern and northeastern regions of the country. Thus, C. angustifolia, C. balansae, and C. saltensis are distributed at the different altitudinal levels of the Yungas Rainforest ecoregion in the northwest, while C. fissilis is present in the Paranaense Rainforest ecoregion in the northeast. Cedrela balansae and C. fissilis are considered important forest resources due to their fast development in forest enrichment systems and their quality timber, making them the most commercially valuable species in both local and international markets [32]. As a result, they have been subjected to intensive logging, which has led to reductions in genetic diversity and alterations in population structure [33,34,35]. At present, C. balansae is classified as endangered and C. fissilis as vulnerable species [5]. Mayrinck [6] also cited C. fissilis as an endangered species for Cerrado and Atlantic Forest Biome of Brazil. In addition, C. fisilis also is included in CITES (https://cites.org/eng/app/appendices.php; accessed on 30 December 2025).
Cedrela balansae C. DC. and C. fissilis Vell. are long-lived, deciduous canopy trees that can reach heights of up to 20 m. Flowering of C. balansae occurs between November and December, whereas that of C. fissilis is from September to December. Fruiting begins immediately after flowering, and capsules require approximately 6–8 months to reach maturity. Fruit development extends until August for C. balansae and July for C. fissilis, when the dehiscent capsules mature and release numerous winged seeds that are primarily dispersed by wind during the dry season [36,37].
At present, ex situ conservation of these species is primarily achieved through living collections established in clonal banks and clonal seed orchards [38]. However, the growing demand for seeds to support propagation, breeding, reforestation, and ecological restoration programs highlights the need for additional conservation strategies. While living collections can provide seeds in the short term, seed banks are essential as a complementary approach to safeguard genetic variability, particularly in the face of changing climatic conditions. Species of Cedrela are reported to have orthodox seeds [39,40], but Silva et al. [39] found that germination and vigor of C. fissilis seeds were greatly reduced during storage. Identifying seed desiccation tolerance, storage behavior, longevity, and temporal variation in these two Cedrela species is essential for prioritizing immediate post-harvest processing, selecting appropriate storage techniques, and planning viability monitoring and regeneration cycles.
Over the last two decades, collection sites (populations) of C. balansae and C. fissilis have been experiencing significant increases in temperatures and decreases in precipitation (Figure S1). Thus, we examined interannual variation in seed traits, desiccation tolerance, storage behavior, and longevity of these two species and their association between each other and with climatic conditions of seed provenance. Additionally, we predict longevity under different storage conditions of MC and temperature using the viability equation. For this, we used three accessions collected from the same population for each species between 2013 and 2016. Specifically, we hypothesized that interannual variation in seed traits and/or in climatic conditions affect desiccation tolerance, storage behavior, and/or seed longevity. We expected that seeds with lower seed mass, MC or lipid content, and/or produced in warmer and drier years would show higher tolerance to desiccation and longevity than seeds with higher mass, MC or lipid content, and/or produced in cooler and moist years.
2. Results
2.1. Seed Traits
Seed mass, lipid content, viability, MC, and/or germination percentage (GP) of fresh seeds varied across years, depending on the species (Table 1). In C. balansae, seeds collected in 2013 showed lower seed mass, viability, MC, and GP than those collected in the other years, while lipid content did not vary among years. Seeds of C. fissilis collected in 2013 showed lower seed mass and MC than in the other years, whereas lipid content in 2014 was higher than in the other years. Viability and GP did not vary over the years. For both species, the time to 50% germination of viable seeds (t50) did not vary among years, being on average 7.38 d for C. balansae and 7.78 d for C. fissilis.
2.2. Seed Desiccation Tolerance
For both species, MC, GP, and t50 varied significantly among study years and/or treatments (Table 1). For C. balansae, the highest MC values were for seeds collected in 2016, while GP was higher in 2015 and 2016 than in 2013. No significant differences were detected in t50 or VLI among years for any treatment. For all years of study, MC was significantly lower in the LiCl treatment than in the other treatments. In 2013, no significant differences in GP or t50 were registered among treatments, whereas in 2015 and 2016, GP was lower and t50 higher in seeds hydrated with KNO_3_ compared to other treatments. In 2013, VLI did not vary between treatments, whereas in 2015 and 2016 VLI values were significantly higher in KNO_3_ than in the other treatments.
For C. fissilis, MC was higher in 2014 in CaNO_3_ than in the other treatments. No significant differences in GP or VLI were registered among years for any treatment, whereas t50 was lower in 2014 than in the other years across all treatments. In all study years, MC was significantly lower in the LiCl treatment than in the other treatments. GP decreased significantly after drying with LiCl and after hydration with CaNO_3_, NaCl, and KNO_3_, with the lowest values registered in KNO_3_. The t50 values increased with increasing MC, reaching their highest values in KNO_3_ in 2014 and 2016 and in NaCl in 2016. In 2013, VLI did not vary between treatments, whereas VLI values were significantly higher in KNO_3_ in 2015 and 2016 and in NaCl in 2016.
2.3. Seed Storage Behavior
For both species, seed storage behavior under different conditions of MC and temperature varied between years (Figure 1 and Figure 2). Thus, for C. balansae, in 2013, GP and t50 were not significantly affected by MC, temperature, storage period, or their interactions (p > 0.05; Figure 1A,B). In contrast, GP in 2015 was significantly affected by all these factors and their interactions (p < 0.05; Figure 1C–F). At 25 °C, GP was significantly reduced after 12 months in seeds stored with 6–8% MC and after 3 months in seeds stored with 10–12% MC. At 5 °C and −20 °C, significant decreases in GP were detected after 24 months in seeds with 10–12% MC and after 3 months in seeds with 15–19% MC, respectively. No germination (dead seeds) was recorded at 25 °C after 24, 12, and 3 months in seeds stored with 6–8%, 10–12%, and 15–19% MC, respectively, or at 5 °C and −20 °C after 24 months in seeds with 15–19% MC. The t50 values were significantly higher in seeds stored at 15–19% MC across all storage temperatures than in the other treatments (on average 11 vs. 6–8 days, respectively).
For C. fissilis, GP and t50 in 2013 were significantly affected by temperature, storage period, and temperature × storage period and MC × temperature × storage period interactions (p < 0.05; Figure 2A,B). Thus, GP at 25 °C was significantly lower after 12 months in seeds stored with 3–5% and 6–8% MC, and t50 was higher (8 vs. 6 days) than in the other treatments. In 2014, GP and t50 were significantly affected by all these factors and their interactions (p < 0.05; Figure 2C–F). At −20 °C, GP decreased significantly after 24 months in seeds stored with 3–5% and after 3 months in seeds with 21–25% MC. At 25 °C, GP was significantly reduced after 3 and 24 months in seeds stored with 12–14% and 6–8% MC, respectively, and no germination (dead seeds) was recorded after 3 and 12 months in seeds stored with 21–25% and 12–14% MC, respectively. At 5 °C and −20 °C, significant decreases in GP were detected after 24 months in seeds with 12–14% MC, and no GP was registered after 12 months in seeds with 21–25% MC. The t50 values were significantly (p < 0.05) higher in seeds stored at 21–25% MC across all storage temperatures than in the other treatments (on average 10 vs. 6–7 days, respectively).
2.4. Seed Longevity
2.4.1. Comparative Longevity
For both species, seed longevity parameters significantly varied between study years (p < 0.05; Table 2). For C. balansae, K_i_, σ^−1^ and p50 were higher in 2015 than in 2013, whereas σ was higher in 2013 than in 2015. For C. fissilis, K_i_ did not differ significantly between years, but σ and p50 were higher and σ^−1^ lower in 2014 than in 2013.
2.4.2. Estimation and Validation of CW and KE Constants
For both species, seed survival curves exhibited a typical sigmoid pattern on the percentage scale, with seeds at 45% eRH (5.2–6% MC) showing a longer lag period, followed by seeds at 60% eRH (8.9–9% MC) and finally by those at ≥75% eRH (≥11.2% MC; Figure 3A,B). For both species, there was no significant difference in K_i_ among MC, being on average 2.22 for C. balansae and 1.66 for C. fissilis (Table 2). However, mean values of σ, p50, and σ^−1^ differed significantly between MC (p < 0.05), with both σ and p50 decreasing and σ^−1^ increasing as MC increased. A negative relationship was observed between seed longevity (log σ) and MC (log m; Figure 3C,D).
The estimated K and C_W_ values were 3.6618 (s.e. = 0.0409) and 2.4983 (s.e. = 0.0395), respectively, for C. balansae and 3.2039 (s.e. = 0.0419) and 1.7706 (s.e. = 0.0489), respectively, for C. fissilis. Using these K values and the universal constants C_H_ (0.0329) and C_Q_ (0.000478), the derived K_E_ values were 5.3988 ± 0.0409 for C. balansae and 3.8677 ± 0.3680 for C. fissilis. A positive relationship was observed between predicted GP by the viability equation and observed GP of storage behavior tests (Figure 3E,F), which validated the estimated constants.
2.4.3. Prediction of Seed Longevity Under Different Storage Conditions
Predictions of seed longevity (p50) indicated that seeds stored with MC > 6.8% at −20 °C ranged between 16 and 27 years for C. balansae and between 2 and 3 years for C. fissilis (Table 3). In contrast, when stored at ambient room temperature (22 °C), p50 values were ≤1 year for C. balansae and <2 months for C. fissilis. However, under standard seed bank conditions (3–5% MC at −20 °C), C. balansae seeds showed a predicted p50 of 115 years, with a σ of 67 years. For C. fissilis, predicted p50 under the same conditions was only 7 years, with a σ of 4 years.
2.4.4. Associations Between Seed Traits, Desiccation Tolerance and Longevity Parameters, and Climatic Variables of Seed Provenance
For both species, Spearman’s correlation analysis revealed significant associations among seed traits, longevity parameters, and/or climatic variables, whereas lipid content and VLI were not correlated with any of the variables considered (p < 0.05; Table 4). In C. balansae, seed mass, MC, K_i_, and p50 were positively intercorrelated, while σ was negatively correlated with these variables. These traits also were positively associated with both precipitation variables (APP and PP_J-J_) and negatively associated with temperature variables, including annual temperatures (MaAT and MeAT) and temperatures during seed development (MaT_J-J_, MeT_J-J_, and MiT_J-J_). Conversely, σ showed negative correlations with both precipitations and positive correlations with all these temperature variables. In C. fissilis, only seed mass was positively correlated with MC. Seed mass, K_i_, and p50 were positively correlated with all annual temperature variables. K_i_ and p50 also were positively correlated with both precipitations and negatively with temperatures during the seed development period.
3. Discussion
Temporal variation in seed traits and functional responses has received comparatively less attention than inter- and intraspecific variability [27]. However, understanding how seed mass, desiccation tolerance, storage behavior, and longevity vary among years is essential for effective seed bank management [42] and for predicting natural regeneration of species in environments increasingly affected by climate change [26,30,43]. In this study, our hypothesis was partially supported, i.e., interannual variation in seed traits and in the climatic conditions of seed provenance did not affect seed desiccation tolerance in C. balansae and C. fissilis, but it did influence seed storage behavior and longevity.
3.1. Seed Traits
Environmental conditions experienced by mother plants strongly influence seed development and, consequently, seed traits [44,45]. High temperatures and/or reduced water availability during seed development frequently affect seed mass, MC, dormancy, and germination, although responses are not always consistent across species [12,46]. In both Cedrela species, the lowest seed mass and MC were recorded in 2013, coinciding with lower accumulated precipitation during seed development compared with other years. Similar patterns have been reported in alpine species, where seed mass is more closely linked to soil water availability than to temperature [47]. In C. balansae, seed viability and germination also were reduced in 2013, whereas in C. fissilis, no interannual differences in these traits were found. This apparently neutral effect of warmer and drier maternal environments on seed viability and germination is consistent with previous reports for threatened Acacia species in Western Australia [27] and alpine Australian species [48].
Our findings have important implications for the resilience of both species under changing climatic conditions. In C. balansae, smaller-winged seeds may favor dispersal to new habitats, but their reduced viability and germination could limit seedling recruitment. In contrast, stability of seed viability and germination in C. fissilis may confer an advantage under changing environmental conditions and during colonization of new habitats.
3.2. Seed Desiccation Tolerance
Seed mass and MC at shedding are commonly used predictors of desiccation tolerance [11,15,49], with desiccation-tolerant seeds typically having a thousand-seed weight (TSW) < 2500 g and MC ≤ 23% [11]. In addition, in environments with dry seasons, such as in the Yuto and El Naranjo localities, most species had low seed MC (<20%), since they usually tolerate dry periods better than those with high MC [14,50,51]. In this case, although seed mass and MC varied among years, seeds of both species consistently showed low TSW (20–27 g) and low MC (5.5–7.5%) and were dispersed in the dry season (in winter months, July and August), strongly indicating desiccation tolerance.
In C. balansae, no differences in germination between fresh and dried seeds (3–5% MC) were detected, and viability loss index (VLI) values were ≤0.05, confirming desiccation tolerance. In C. fissilis, drying reduced germination by 9–14%, with VLI values between 0.09 and 0.15, classifying the seeds as potentially desiccation-tolerant [49]. Similar partial reductions in germination after drying have been reported for other tree species [52,53], supporting the idea of a continuum in desiccation tolerance rather than discrete categories, in which some species and/or populations are more susceptible to desiccation than others [18]. On the other hand, seed hydration above 18% MC reduced germination and increased t50, probably due to enhanced metabolic damage and fungal proliferation [42]. Nevertheless, even under these conditions, VLI values remained within the range of potential desiccation tolerance, reinforcing the overall orthodox behavior of both species.
3.3. Storage Behavior
Standard seed banking protocols involve drying seeds to 3–7% MC and storing them at −20 ± 2 °C (base collection) or 5 ± 2 °C (active collection) [10]. While effective for most crops, wild tree species often show heterogeneous responses to storage conditions [26]. Moreover, storage behavior is frequently inferred from single-seed lots, limiting broader inference. In our study, germination and t50 were influenced by seed MC, storage temperature, storage duration, and collection year, with species-specific responses. In C. balansae, seeds dried to 3–5% MC consistently maintained viability across all storage temperatures, whereas seeds with 6–8% MC showed year-dependent responses, with seeds collected in 2015 remaining viable only when stored at 5 and −20 °C. In C. fissilis, seeds with both 3–5% and 6–8% MC maintained viability at 5 and −20 °C in both years; however, seeds collected in 2014 with 3–5% MC lost viability at −20 °C after 24 months of storage. This loss of viability was probably associated with the higher lipid content of these seeds, consistent with reports of lipid crystallization damage during freezing in oily seeds [54].
On the other hand, seeds stored at MC > 10–12% retained viability for up to 1 year at low temperatures but deteriorated thereafter, while seeds with MC > 15–21% lost viability rapidly, probably due to physical damage caused by lethal ice formation, reinforcing the advice against storing seeds with high MC [55]. At 25 °C, viability loss at MC ≥ 6% was attributable to physiological ageing processes [56], including lipid peroxidation, as previously described for C. fissilis [39,57].
Overall, both species can be classified as orthodox, in agreement with previous reports [6,40,41,58]. However, for C. fissilis, lipid content should be carefully considered prior to long-term storage at −20 °C, since viability may decrease if seeds are stored at −20 °C. Hamilton et al. [16] suggested that for oily seeds with variable responses and storage limitations, cryopreservation is the most reliable long-term conservation strategy. For this species a preliminary study showed that cryopreservation of seeds tolerated freezing in LN without any loss in viability when MC was in the range of 3–5%, indicating that this technique would be a reliable strategy. Further studies are needed to define the optimal hydration window for cryogenic storage, which can be very wide for orthodox seeds [59].
3.4. Seed Longevity
Seed longevity is central to seed bank management for planning viability monitoring intervals and regeneration cycles, and it is also key to understanding soil seed persistence [60]. In our study, accelerated ageing tests revealed interannual variation in longevity, with both species showing the lowest p50 values in seeds collected in 2013. White et al. [30] demonstrated that such interannual variation in seed longevity may arise from differences in K_i_, in the time required for viability to fall one NED (σ), or from variation in both parameters. Consistent with this, the reduced p50 observed in C. balansae was primarily associated with lower initial viability (K_i_), whereas in C. fissilis it was mainly linked to lower σ values. Because p50 is inherently dependent on K_i_, σ has been proposed as a more appropriate measure of intrinsic seed longevity, since it is independent of initial seed quality [61]. Based on p50 values ranging from 28 to 47 days, seeds of both Cedrela species would be classified as medium-lived (p50 > 10 to ≤100 d) [22], similar to what has been reported for other tree species [21,53]. However, relative longevity classifications are valid only under the specific ageing conditions under which they are derived [54]. For example, these authors reported σ values of 30.21 days for Xanthorrhoea preissii seeds aged at 35 °C with 10% MC but only 6.63 days when seeds were aged at 45 °C with the same MC. Therefore, additional ageing tests conducted under standardized conditions (45 °C and 60% RH) would be required to determine more robustly whether the Cedrela species should be classified as medium- or short-lived.
The seed viability equation can provide accurate predictions of seed longevity when applied within a valid range of conditions, and it is widely used to describe how orthodox seeds lose viability during storage under different combinations of seed MC and temperature [20,23,54,62]. However, reliable application of this model requires an estimation of species-specific constants [20]. In this study, we used four MC levels at 35 °C to estimate these constants and associated longevity parameters. For both species, the constant K_i_ did not differ significantly among storage environments, reflecting the use of a single-seed lot and satisfying a key assumption of the seed viability equation [20]. Survival curves differed among MC treatments, with seeds stored at lower MC exhibiting greater longevity (higher p50) than those stored at higher MC. Consistent with this pattern, a negative relationship was observed between seed longevity (log σ) and seed MC (log m), highlighting the importance of seed drying for prolonging longevity. The estimated C_W_ constant, which describes the relative effect of MC on longevity, was 2.4983 for C. balansae and 1.7706 for C. fissilis. These values are lower than those reported for C. odorata (C_W_ = 3.80) [62], possibly reflecting its higher seed oil content (21%) compared with our Cedrela species (13–19%), although relationships between oil content and C_W_ are not consistent across studies [23]. The K_E_ constant, which represents inherent seed longevity [20], was estimated at 5.3988 for C. balansae and 3.9729 for C. fissilis. These values also are lower than those reported for C. odorata (K_E_ = 6.90) [62], indicating higher inherent longevity in C. odorata, followed by C. balansae and then C. fissilis. Moreover, the C_W_ and K_E_ constants were validated using independent data from the storage behavior experiments, confirming their suitability for predicting longevity in other seed lots of each species.
Identifying the period during which seed viability can be maintained under different storage conditions is particularly relevant for forest researchers working with species that have abundant seed production in some years and scarcity in others, as is the case for C. balansae. Using the viability tools of the Seed Information Database [41] together with our estimated constants, we assessed the potential longevity of seeds stored under different combinations of MC and temperature. C. balansae exhibited high longevity, with a p50 on the order of hundreds of years and a loss of one NED of viability occurring after 67 years, indicating strong potential for long-term ex situ conservation. In contrast, C. fissilis showed much lower longevity, with a p50 of around 7 years and a comparable decline in viability occurring after approximately 4 years, highlighting its greater sensitivity to temperature and MC during storage despite being desiccation-tolerant. Moreover, under uncontrolled ambient conditions (≈22 °C and MC > 6%), both p50 and σ declined sharply, reaching 1 year or less for C. balansae and less than 2 months for C. fissilis. According to these estimations, and consistent with results from comparative longevity tests, C. balansae can be classified as having medium-lived seeds, whereas C. fissilis appears to have short-lived seeds, contrary to comparative longevity test classification but in agreement with reports for this species by Carvalho [63]. Furthermore, our results for C. fissilis support the hypothesis proposed by Long et al. [43] that seed lifespan estimated under laboratory conditions reflects persistence under natural conditions, since this species also has been reported to form a transitory soil seed bank [64].
3.5. Associations Between Seed Traits, Desiccation Tolerance, and Longevity Parameters and Climatic Variables of Seed Provenance
Climate change is increasingly influencing tree reproductive processes by altering phenology, seed production, seed functional traits, and seedling establishment, with potential long-term consequences for species dispersal, persistence, and conservation [1,2]. Our correlation analyses showed that lipid content for both species was not associated with other seed traits, including desiccation tolerance and seed longevity, or with climatic variables, in agreement with previous findings across a wide range of species [9,24]. Furthermore, seed desiccation tolerance, estimated using the viability loss index (VLI), was not related to seed traits, seed longevity, or climatic variables, suggesting that desiccation tolerance may be a heritable trait and therefore species-specific [13].
Conversely, for C. balansae, seed mass and MC were positively associated with each other and with precipitation and negatively associated with temperature, both at the annual scale and during seed development. These results are consistent with those of Cochrane [27] and Vázquez-Ramírez and Venn [48], who reported that seed mass tends to decrease under warmer and drier environmental conditions. Regarding seed longevity, both K_i_ and p50 were positively associated with seed mass, MC, and precipitation and negatively associated with temperature (both at the annual scale and during seed development). These trends contrast with findings from other studies reporting that larger seeds and/or seeds produced in warmer and drier years exhibit lower p50 values than smaller seeds and/or seeds produced in cooler and wetter years [9,24,30]. In contrast, σ showed an inverse pattern, increasing with temperature and decreasing with precipitation, suggesting that seeds produced under warmer and drier conditions may exhibit greater resistance to aging than those produced in colder and wetter years. This pattern is consistent with Mondoni et al. [29], who demonstrated that warmer conditions during seed development enhanced resistance to aging in alpine species, supporting the idea that seed longevity may exhibit adaptive responses under future warming scenarios. As proposed by these authors, adaptive adjustments in seed longevity driven by transgenerational plasticity may therefore play a fundamental role in species survival and persistence under future environmental challenges, such as climate warming.
In contrast, for C. fissilis, seed mass was the only trait positively correlated with MC and with annual temperature, a pattern similar to that reported for some alpine species [65]. In addition, K_i_ and p50 were positively correlated with annual temperature and precipitation (both at annual scale and during seed development) but negatively correlated with temperatures during seed development period. These results indicate that the thermal conditions experienced by maternal plants during the reproductive phase are particularly critical for determining seed longevity. In this context, several studies have emphasized that environmental conditions during seed development are decisive in the determination of seed longevity [2,24,30]. Similarly, the positive relationship between seed longevity and precipitation is consistent with findings by Merritt et al. [66] for Australian species; however, the mechanisms linking precipitation to seed longevity remain poorly understood and require further investigation [30]. However, if we consider that C. fissilis is naturally distributed in the subtropical Paranaense Rainforest (Atlantic Forest) of northeastern Argentina, which is characterized by a humid subtropical climate without a dry season, an average annual rainfall of approximately 2.000 mm, and a mean annual temperature of 20.2 °C, it is therefore not surprising that seed traits and longevity increased with temperature and precipitation and that temperature during seed development can be a critical factor influencing initial seed quality and longevity. Further research, including seed collections from populations across the species’ natural distribution, is required to establish the relative contributions of genetic and environmental factors in determining seed traits and longevity.
Therefore, as climate change progresses with increasing temperatures and decreasing precipitation, the window of opportunity for successful germination and seedling establishment is likely to become shorter for both species, as has been reported for other tree species [67]. For C. balansae, reductions in seed mass, MC, initial seed quality, and longevity are likely to negatively affect seed germination and seedling establishment; however, an increase in seed deterioration (σ) may enhance seed persistence in the soil, thereby increasing the likelihood of germination when environmental conditions become favorable. In contrast, C. fissilis may remain unaffected or may even exhibit an increase in seed mass, initial seed quality, and longevity under higher annual temperatures; however, if temperatures increase and precipitation decreases during seed development, both initial seed quality and longevity are expected to decline, reducing seed persistence in the soil and limiting germination and seedling establishment. In this context, these findings highlight the importance of ex situ conservation through seed banking for both species.
4. Materials and Methods
4.1. Seed Collection
Mature seeds were collected at the end of July for C. balansae and June for C. fissilis from 20 individuals per species in two seed production stands located in the northwestern provinces of Argentina (Jujuy and Tucumán). Seeds of C. balansae were obtained in 2013, 2015, and 2016 from Yuto locality in the Jujuy province (23°38′ S, 64°28′ W; 349 m a.s.l.). Seeds of C. fissilis were collected in 2013, 2014, and 2016 from El Naranjo locality in the Tucumán province (26°39′ S, 65°02′ W; 665 m a.s.l.) as a seed-source provenance introduced from Guaraní reserve (26°55′ S, 54°13′ W; 482 m a.s.l.) in the Misiones province of Argentina. The climate in both collection sites is characterized by two well-defined seasons: a dry season from late April to September and a wet season during the remainder of the year, with more than 80% of annual precipitation occurring between October and March (Figure 4).
4.2. Seed Traits
For each species and year of collection, immediately after seed collection, seed mass, lipid and moisture content (%), viability and germination (%), and the time to 50% germination were determined in fresh seeds. The mass of 1000 seeds (TSW) was estimated by weighing five replicates of 100 seeds each with a precision balance to 0.0001 mg accuracy (Denver Instrument APX-200, Denver Instrument Company, Denver, CO, USA). Total lipid content was determined using a modified protocol [68]. Three pre-weighed samples of ground seeds (2.0 g) were extracted overnight with petroleum ether. Then, the solvent was removed via evaporation, and the extract was further purified by dissolving it in chloroform and filtering through Whatman No. 1 paper. The combined filtrate and washings then were dried and weight. The amount of lipids per gram of seed dry weight was calculated (expressed in % dry weight basis). Seed moisture content (MC) was determined gravimetrically by drying three replicates of 10 seeds each at 103 °C for 17 h (expressed in % fresh weight basis). Seed viability (%) was determined in 100 seeds using the tetrazolium chloride staining technique [69].
Seed germination was determined in four replicates of 25 seeds each for each year. Seeds were sown in Petri dishes on two sheets of filter paper moistened with distilled water and incubated in a chamber at 25 °C with a photoperiod of 12 h light/12 h darkness (optimal conditions). The number of germinated seeds (i.e., with radicle emergence) was recorded daily for 30 days. At the end of all germination tests, when no additional germination had occurred for 2 weeks, a cut test was conducted to determine the viability of the non-germinated seeds (soft or firm, i.e., dead or viable, respectively). The number of viable seeds per replicate was the number of firm non-germinated seeds + the germinated seeds. The final germination percentage (GP) was calculated based on the total number of viable seeds. The accumulated germination data were adjusted using the sigmoidal dose–response equation, and time to 50% germination of viable seeds (t50) for each treatment was estimated by linear interpolation between points on the germination curve adjacent to 50% germination.
4.3. Seed Desiccation Tolerance
In 2013, for both species, two sub-lots of 130 seeds each were placed in tulle bags and stored inside a sealed glass container (desiccator) at ambient temperature (22 °C), and MC was adjusted by drying over saturated solutions of LiCl (13% equilibrium relative humidity, eRH) to 3–5% MC and CaNO_3_ (50% eRH) to 6–9% MC [51]. The eRH inside each desiccator was monitored weekly using dataloggers (HOBO Temp/RH logger UX100-011, USA). After approximately 4 weeks, when the eRH of each desiccator was stabilized, GP and MC were determined. GP of fresh (untreated) seeds was used as a control. In 2014 for C. fissilis and in 2015 for C. balansae, two additional salt solutions were included: NaCl (75% eRH; 10–14% MC) and KNO_3_ (91% eRH; 15–25% MC), following the procedure described for LiCl and CaNO_3_.
Seed desiccation tolerance was estimated using the viability loss index (VLI) [49]:
VLI values theoretically vary from 0 (desiccation-tolerant seeds) to 1 (desiccation-sensitive seeds). Thus, seeds with VLI > 0.95 were considered as desiccation-sensitive, with 0.95 > VLI > 0.5 as potentially desiccation-sensitive, with VLI < 0.05 as desiccation-tolerant, and with 0.05 < VLI < 0.5 as potentially desiccation-tolerant. In addition, we also estimated VLI for seeds submitted to CaNO_3_, NaCl, and KNO_3_ saturated salt solution to evaluate tolerance.
4.4. Seed Storage Behavior
In 2013, a factorial design experiment combining two MC levels (3–5 and 6–8%), three temperatures (−18, 5, and 25 °C), and two storage periods (3 and 12 months) was conducted for each species. Two sub-lots of 1700 seeds each were equilibrated in desiccators containing LiCl and CaNO_3_ and kept at 22 °C until the eRHs were reached. After approximately 4 weeks, MC and GP were determined for each seed lot. Then, 12 samples of 130 seeds each were sealed in aluminum-foil bags and stored at the specified temperatures [51]. In 2014 or 2015, depending on the species, two additional MC ranges were included (NaCl: 11–14% MC and KNO_3_: 15–25%MC). Accordingly, a factorial design was implemented combining four MC levels (3–5%, 6–8%, 10–14%, and 15–25%), three storage temperatures (−20°, 5°, and 25 °C), and three storage periods (3, 12, and 24 months). For each species, 5000 seeds were equilibrated in desiccators with LiCl, CaNO_3_, NaCl, and KNO_3_ and kept at 22 °C until eRHs were reached. After 4 weeks, MC and GP were determined, and 36 samples of 130 seeds each were sealed in aluminum-foil bags and stored at the specified temperatures. Each treatment (MC × temperature × storage period) was represented by one sample per bag. In all study years, MC and GP were evaluated for each treatment.
4.5. Seed Longevity
4.5.1. Comparative Seed Longevity
In 2013, 2014, and 2015 (depending on the species), 11 samples of 100 seeds each were stored in tulle bags and placed into a desiccator with a saturated salt solution of NaNO_2_ (60% eRH) and kept in a chamber at 35 °C. One sample for each species was removed after each of 2, 5, 9, 20, 33, 54, 75, 100, 125, 140, and 152 days for germination testing. Germination data were plotted as a seed survival curve, where seed viability (GP) was plotted against ageing period. Time for viability to decline to 50% (p50) was estimated using the following equation:
Species were ranked by p50 and assigned to the categories of short-lived (p50 > 1 and ≤10 days), medium-lived (p50 > 10 and ≤100 days), and long-lived (p50 > 100 and ≤1000 days), based on [22].
4.5.2. Estimation and Validation of CW and KE Constants
In 2014 or 2015 (depending on the species), 14 samples of 100 seeds each were stored in tulle bags and placed into desiccators containing saturated salt solutions of CaNO_3_ (45% eRH), NaNO_2_ (60% eRH), NaCl (75% eRH), and KNO_3_ (88% eRH) at 35 °C. One sample per treatment was removed at scheduled intervals (10–14 dates) depending on how quickly seeds were losing viability and tested for germination. Loss of viability ranged between 42 days in KNO_3_ and 510 days in CaNO_3_ for C. balansae and between 39 days in KNO_3_ and 321 days in CaNO_3_ for C. fissilis.
The GP data from serial germination tests at the respective seed MC were fitted by probit analysis according to the viability equation (Equation (1)) [20]. This generated the fitted seed survival curves from which K_i_, σ, σ^−1^, and p50 were determined. To estimate K_E_ and C_W_ constants, the relationship between storage seed MC and seed longevity was quantified using linear regression, where log σ was plotted against the log of seed MC (m) to obtain the value of C_W_ by fitting the following equation:
The value of K_E_ was estimated by using the universal values of C_H_ (0.0329) and C_Q_ (0.000478) as well as K (Equation (3)) at ageing temperature (t) equal to 35 °C, as presented in the following equation:
For validating the newly derived viability constants (C_W_ and K_E_), we used the GP from the seed storage behavior experiments (see Section 2). Thus, we estimated initial viability (K_i_) of stored seeds under each combination of MC and temperature, with C_W_ and K_E_, and with the viability equation. We predicted the GP for each combination and storage interval. The observed GP values were regressed against predicted GP values using linear regression.
4.5.3. Prediction of Seed Longevity Under Different Storage Conditions
For both species, we used the estimated parameters of the seed viability equation to predict σ and p50 for a collection with an initial viability of 95% (1.64 NED) under a range of MCs and storage conditions. The MCs evaluated were the (1) MC of seeds at time of dispersal (e.g., 6.8% for both species), (2) MC of seeds equilibrated at standard seed bank drying conditions of 15% RH and 15 °C (C. balansae: 3.8% and C. fissilis: 4.3%), and (3) estimated MC of seeds equilibrated at ambient conditions with 50% RH and 22 °C (C. balansae: 8.3% and C. fissilis: 8.9%). Storage temperatures considered were −20 °C and 5 °C, standard seed bank storage conditions [10], and 22 °C (ambient room temperature of storage at clonal seed orchard facilities). Predictions were obtained using the viability tools available in the Seed Information Database [41].
4.5.4. Climate Data
Climate information for the collection sites was sourced from NASA POWER. Maximum (Ma), mean (Me), and minimum (Mi) daily temperatures, as well as total precipitation (PP), were downloaded for the period 2000–2024. To assess long-term trends, daily data was aggregated to an annual level (MaAT, MeAT, MiAT, and APP; Figure S1). For correlation analyses, for each year of the study, MaAT, MeAT, MiAT, and APP and maximum, mean, and minimum temperatures and total precipitation (MaT_J-J_, MeT_J-J_, and MiT_J-J_, PP_J-J_, respectively) during seed development (January–July for C. balansae and January-June for C. fissilis) were also considered (Figure 4).
4.6. Statistical Analysis
General linear mixed models were used to analyze seed mass, lipid content, MC, and t50, K_i_, σ, σ^−1^, and p50 and to compare among years and/or treatments for each species. Variance heterogeneity was considered, and the AIC and BIC information criteria were used to select the best-fitted model. General linear mixed models for binomial distribution and logit link functions were used to explain variations in number of germinated seeds. As the number of viable seeds was not the same for all experimental units, this number was entered as a covariable. For all analysis, year, desiccation tolerance, and storage behavior treatment factors were considered as fixed effects. In all cases, all factor interactions were included in the models, and same information criteria were used to select the most parsimonious model, and the residual deviance/degrees of freedom ratio was calculated to assess if the goodness-of-fit of the model was reasonable and if there was over-dispersion. Treatments with no variance (none or all of the seeds germinated in each replicate) were excluded from the analyses. The post hoc DGC test of multiple comparisons of means was used to locate differences among factor levels or their combinations of means when the effect of the factor or their interaction was significant (p < 0.05). We explored the associations of seed traits (seed mass, lipid content, and MC), desiccation tolerance (VLI), and seed longevity parameters (K_i_, σ, and p50) between each other and with climatic variables (MaAT, MeAT, MiAT, AP, MaT_J-J_, MeT_J-J_, MiT_J-J_, and PP_J-J_), evaluating the significance of the Spearman correlation coefficient. All statistical analyses were conducted using the InfoStat software package (version 2017) [70].
5. Conclusions
This study demonstrates that interannual climatic variability during seed development plays a central role in shaping seed traits, storage behavior, and longevity in C. balansae and C. fissilis, with important implications under ongoing climate change. While both species produce desiccation-tolerant seeds, desiccation tolerance itself was largely independent of seed traits and climatic conditions, supporting the idea that it is primarily a species-specific characteristic. In contrast, seed mass, MC, initial seed quality, and longevity were strongly influenced by temperature and precipitation, particularly during the reproductive phase, highlighting the importance of maternal environmental effects on seed performance. Marked interspecific differences were observed in seed longevity and storage potential. C. balansae exhibited medium-lived seeds and comparatively high longevity under optimal storage conditions, indicating strong potential for long-term ex situ conservation, whereas C. fissilis showed short-lived seeds, much shorter longevity and greater sensitivity to storage environment, consistent with its transitory soil seed bank behavior. Correlations between climatic variables and longevity parameters suggest that future increases in temperature and reductions in precipitation may shorten the window for germination and seedling establishment in both species, although species-specific responses are expected depending on the timing of climatic stress. Overall, our findings highlight that seed longevity is a dynamic trait influenced by both environmental conditions and transgenerational plasticity, which may mediate species responses to future climate scenarios. Given the predicted shifts in temperature and precipitation regimes and the contrasting longevity observed between species and year of collection, ex situ conservation through seed banking represents a key strategy to safeguard genetic diversity and support the long-term persistence of these endangered forest species. Additionally, cryopreservation should be considered as a complementary approach for their long-term conservation. To our knowledge, this is one of the first studies to integrate interannual climatic variability, seed trait expression, and longevity parameters in subtropical tree species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baskin C.C. Baskin J.M. Plant Regeneration from Seeds: A Global Warming Perspective Academic Press London, UK 2022
- 2Bernareggi G. Carbognani M. Petraglia A. Mondoni A. Climate warming could increase seed longevity of alpine snowbed plants Alp. Bot.2015125697810.1007/s 00035-015-0156-0 · doi ↗
- 3Long Q. Wang F. Ge W. Jiao F. Han J. Chen H. Cai L. Temporal and spatial change in vegetation and its interaction with climate change in Argentina from 1982 to 2015 Remote Sens.202315192610.3390/rs 15071926 · doi ↗
- 4Peri P.L. Uso Sostenible del Bosque. Aportes Desde la Silvicultura Argentina 1st ed. Peri P.L. Martinez Pastur G.J. Schlichter T.M. Ministerio de Ambiente y Desarrollo Sostenible Buenos Aires, Argentina 2021
- 5IUCN The IUCN Red List of Threatened Species, Version 2025–2Available online: https://www.iucnredlist.org(accessed on 1 December 2025)
- 6Mayrinck R.C. Vilela L.C. Pereira T.M. Rodrigues-Junior A.G. Davide A.C. Vaz T.A. Seed desiccation tolerance/sensitivity of tree species from Brazilian biodiversity hotspots Trees 20193377778510.1007/s 00468-019-01815-8 · doi ↗
- 7Gallo L. El bien común genético forestal de la República Argentina Instituto Nacional de Tecnología Agropecuaria (INTA)Buenos Aires, Argentina 2024
- 8Hay F.R. Probert R.J. Advances in seed conservation of wild plant species: A review of recent research Conserv. Physiol.20131 cot 03010.1093/conphys/cot 03027293614 PMC 4806614 · doi ↗ · pubmed ↗