Dietary Use of Hericium coralloides for NAFLD Prevention
Darya Chekushkina, Oksana Kozlova, Elena Vechtomova, Alexander Prosekov

TL;DR
This paper reviews the potential of Hericium coralloides as a dietary supplement for preventing non-alcoholic fatty liver disease due to its bioactive compounds.
Contribution
The paper highlights the use of pure metabolites from H. coralloides for targeted therapeutic effects in functional foods.
Findings
H. coralloides contains bioactive metabolites like lovastatin and ergothioneine effective against NAFLD in vitro and in vivo.
Dietary supplements using pure metabolites allow for standardized dosing and targeted therapeutic benefits.
H. coralloides is a rare species, so in vitro cultivation is necessary for industrial applications.
Abstract
Introduction: Today, scientists are searching for alternative approaches to preventing metabolic diseases, particularly non-alcoholic fatty liver disease, which reduces the healthy life expectancy of the working population. Fungi, such as Hericium coralloides (Scop.) Pers., are promising raw materials for extracting bioactive substances with preventative potential. Materials and Methods: This review covered review and research articles published over the last 42 years and indexed in the databases of the eLIBRARY.RU, the National Center for Biotechnology Information, and Scopus. Results and Discussion: It has been established that H. coralloides is valued for its nutritional properties due to its rich protein, fat, and mineral composition. It is in demand for pharmaceutical purposes due to its content of bioactive metabolites. The most studied metabolites are lovastatin and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
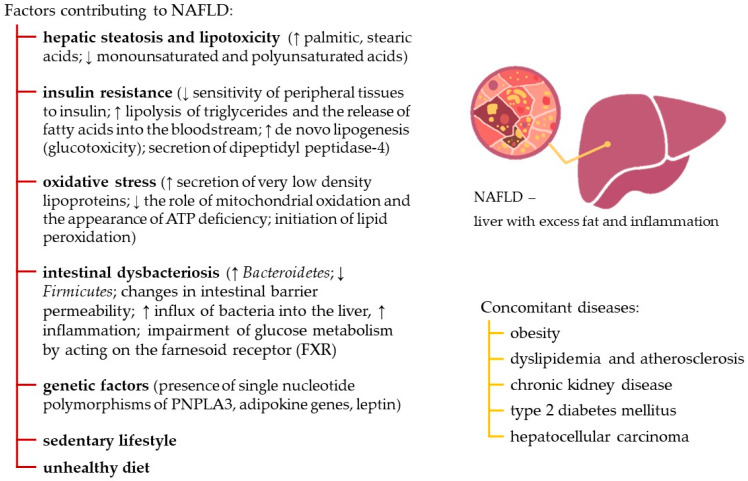 Figure 1
Figure 1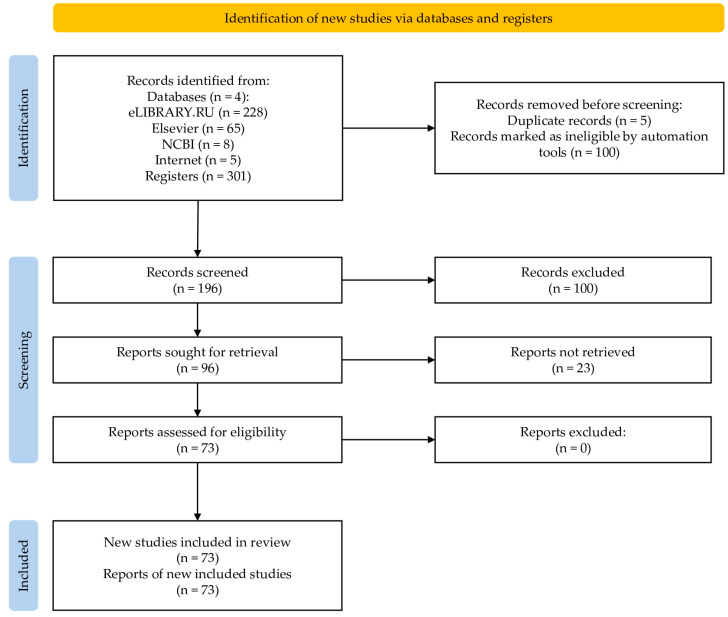 Figure 2
Figure 2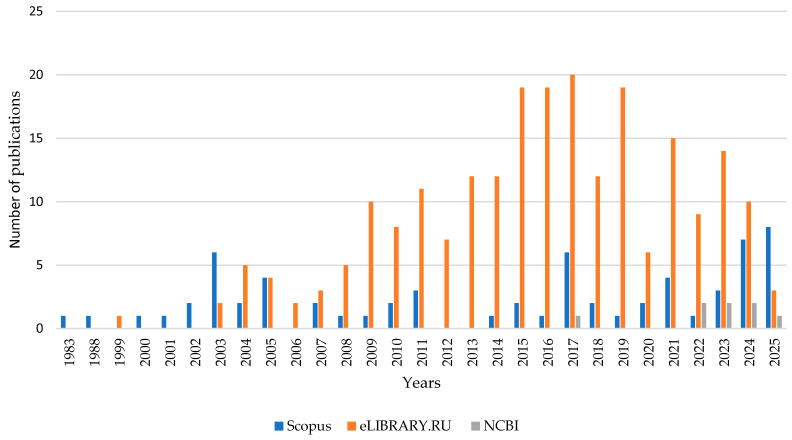 Figure 3
Figure 3 Figure 4
Figure 4- —Kemerovo State University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Biology and Applications · Silymarin and Mushroom Poisoning · Pharmacological Effects of Natural Compounds
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is a multifactorial disease characterized by complex multicomponent pathogenesis and a large number of associated conditions, Figure 1 [1].
NAFLD is characterized by excessive fat accumulation in hepatocytes without evidence of alcohol abuse. Significant factors contributing to the disease include lipid metabolism disorders, oxidative stress, inflammatory processes, intestinal microbiota imbalance, and nutritional disorders (high-calorie diets with excess saturated fats and refined carbohydrates), as well as a sedentary lifestyle [3]. Also important are metabolites of the gastrointestinal microbiota, which affect liver cells through the gut–liver axis [4].
In recent decades, NAFLD has become one of the most common chronic liver diseases worldwide. According to the latest epidemiological data, its prevalence reaches 25–30% of the population in developed countries and continues to grow due to the increasing incidence of obesity, metabolic syndrome [5], and type 2 diabetes mellitus [6,7].
Currently, conventional treatments for NAFLD include lifestyle modification (diet and exercise) and drug therapy aimed at reducing the risk factors such as hyperlipidemia and insulin resistance [8]. However, there is still a lack of effective and safe drugs for the treatment of NAFLD [9]. This necessitates the development of new effective approaches to prevention and therapy, including nutritional support in the form of nutritional agents and natural bioactive substances [10]. Recent studies have emphasized the role of natural compounds, primarily polysaccharides, antioxidants, and anti-inflammatory metabolites, in regulating lipid metabolism, reducing hepatocellular stress, and modulating intestinal microbiota [11].
The nutraceutical market has recently seen an active development of bioactive supplements and functional foods with preventative and therapeutic effects on NAFLD [12]. These products include polysaccharides, flavonoids, phytosterols, vitamins, and microelements that affect fat and glucose metabolism, as well as the state of the intestinal microbiota [13,14]. There is evidence that plant metabolites, such as resveratrol, exert a protective effect against NAFLD by modulating the intestinal microbiota. The study observed an improvement in the density of intercellular contacts of enterocytes reduces the translocation of lipopolysaccharides into the portal bloodstream and, accordingly, reduces the activation of the TLR4/NF-kB-dependent inflammatory cascade in the liver, which is considered as a key factor in the transition of steatosis to steatohepatitis [15]. Thus, the regulation of the “gut–liver” axis may be one of the most promising areas of application of polysaccharide fractions of H. coralloides in preventive nutrition. Of great importance are plant and fungal extracts rich in polyphenols, as well as microbial metabolites containing organic acids, amino acids, vitamins, and other bioactive substances [16]. These metabolites promote health by preventing metabolic syndrome and maintaining a healthy lifespan [15,17].
The fungus Hericium coralloides (Scop.) Pers., a member of the Hericiaceae family, is highly valuable. It contains a wealth of bioactive compounds, such as polysaccharides, hericenones, ericenes, and other metabolites with pronounced antioxidant, anti-inflammatory, and neuroprotective properties [18].
In experimental models, Hericium fungal polysaccharides demonstrate the ability to reduce the expression of lipogenic transcription factors (SREBP-1c, FAS) and activate the AMPK signaling pathway, which leads to a decrease in triglyceride accumulation in hepatocytes and an improvement in insulin sensitivity [19].
Despite the substantial body of research on Hericium erinaceus, data on Hericium coralloides remain fragmented and are mainly focused on its neurotrophic properties, whereas its potential in the context of metabolic liver diseases has not been systematically evaluated. Transcriptomic and chemical profiling studies indicate species-specific differences in secondary metabolite composition within the genus Hericium, including higher expression of terpenoid biosynthesis–related genes in H. coralloides, which may be relevant to the regulation of hepatic lipid metabolism and inflammatory responses. Therefore, the present review aims to bridge the gap between fundamental mycological research and applied nutraceutical approaches for the prevention of NAFLD [20].
Our aim was to conduct a systematic review of the scientific literature on the use of Hericium coralloides for the prevention of NAFLD. To achieve this aim, we set the following objectives:
- (1)Review general information about Hericium coralloides;
- (2)Examine the metabolic composition of the fungus, methods for its extraction, and extract yield;
- (3)Analyze the market of dietary supplements containing Hericium coralloides (or its metabolites).
Thus, we sought to gather up-to-date information on the use of Hericium coralloides and determine the feasibility of using its metabolites to produce dietary supplements for preventing NAFLD.
2. Materials and Methods
The search covered review and research articles published in Russian and English over the last 42 years and indexed in the Russian Scientific Electronic Library (eLIBRARY.RU), the National Center for Biotechnology Information (NCBI) and Scopus (Elsevier). The market of products, which included Hericium coralloides, was studied, for which Internet resources were used.
The search keywords were “Hericium coralloides metabolites,” “Hericium coralloides-based dietary supplements,” “Hericiaceae,” “Hericium coralloides,” “metabolic diseases,” and “NAFLD prevention” (thematically related to the food industry, medicine, and biotechnology).
We focused on the papers describing the results of clinical and preclinical studies (regardless of the model objects), as well as review articles. We excluded the articles describing the results obtained in silico, conference proceedings, preprints and monographs.
The results of the literature search are shown in Figure 2.
3. Results and Discussion
The search of publications using the keyword “Hericium coralloides” showed a limited number of articles in the databases, Figure 3.
The first article on H. coralloid was published in 1983 [22].
The largest numbers of publications in Elsevier were 6 in 2003 (1983–2010), 6 in 2017 (2010–2020), and 8 in 2025 (2020–2025).
The largest numbers of publications in eLIBRARY.RU were 10 in 2009 (1983–2010), 20 in 2017 (2010–2020), and 15 in 2021 (2020–2025).
A total of 301 publications were returned by the search using the keyword “Hericium coralloides.” However, having screened all of them, we selected for this review 10 articles from eLIBRARY.RU, 9 articles from NCBI (excluding those also included in Elsevier), and 42 articles from Elsevier (excluding those also included in PubMed).
3.1. Information on Hericium coralloides
The genus Hericium belongs to the family Hericiaceae, the order Russulales, and the class Agricomycetes. Its 15 species are distinguished macroscopically by the presence of branched and unbranched hymenophore structures supporting spines of varying lengths, as well as by their occurrence in single or multiple clusters. Microscopically, they are distinguished by the presence of amyloid-ornamented basidiospores [23], Figure 4.
The species Hericium coralloides (Scop.) Pers., commonly known as the “coral tooth,” is a striking representative of this genus. Superficially resembling coral, the fungus has bright white coloration, a narrow base, and coral-like branches densely covered with spines. H. coralloides primarily grows on the trunks of broad-leaved trees or rotten wood in northeast, northwest, and southwest China [24]. This rare edible and medicinal mushroom is valued for its beneficial nutritional and pharmacological properties.
Recent years have seen a growing interest in using H. coralloides in the food industry and biotechnology. Importantly, its cultivation methods play a key role in determining the quality of the resulting product and its commercial potential [24].
There are two main methods of cultivating H. coralloides: in natural conditions, the forests where the fungus grows on wood, and in vitro. Both methods have their advantages and limitations, which determine the future use of the fungus’s fruiting bodies in research and industry. In nature, Hericium coralloides can be found on old and decaying deciduous trees in temperate climates. The fungus grows slowly and requires rather specific conditions in terms of humidity, temperature, and wood composition. The influence of these factors on the chemical profile and biological activity of the fungus’s fruiting bodies is largely determined by the ecosystem in which it grows [25]. Such growing conditions have limited scalability and can also expose the fungus to external factors, such as competition with other species and wood diseases. This limits the commercial use of Hericium coralloides grown in natural conditions. Moreover, due to the high demand for the fungus, it has become a rare species in some regions. In Russia, for example, it is listed in the Red Data Books of 46 constituents [26,27].
In vitro cultivation of Hericium coralloides significantly expands the possibilities for its industrial production with controlled quality. In laboratories, the fungus can be grown in sterile media at optimal temperatures and humidity, with its chemical composition thoroughly controlled. Researchers can improve the concentration of certain active substances, such as polysaccharides and phenolic compounds, which is important for their use in the pharmaceutical and food industries [28].
The fungi of the genus Hericium boast a diversity of structurally complex bioactive compounds both in their fruiting bodies and in the mycelium. However, their chemical profile can vary significantly under the influence of cultivation conditions. According to Guan Y. et al., the fruiting bodies of H. coralloides cultured in vitro contained about 33% polysaccharides, 16% crude protein, 12% total ash, 8% reducing sugars, 6% crude fat, 5% crude fiber, 3% total triterpenoids, 1% total saponins, 0.88% total flavonoids, and 0.5% total alkaloids. In addition, the total contents of sterols and phenols were found at 0.43% and 0.18%, respectively [27,29].
Nutritionally, H. coralloides has about 8–24% protein, a moderate fat content, and a rich mineral composition [30,31].
According to [32,33], the fungus’s genome was 43.7 Mb in size, consisted of 13 chromosomes, and had an N50 of 3.60 Mb. Its chromosome assembly efficiency was 96.9%. The transcriptor analysis revealed high expression of genes responsible for the terpenoid backbone and diterpenoid biosynthesis in the mycelium. In addition, the researchers identified the key genes involved in terpenoid biosynthesis. Their results also suggest the potential for commercial exploitation of H. coralloides terpenoid metabolites.
3.2. H. coralloides Metabolites, Methods of Their Extraction, and Extract Yield
Zhang et al. [34] aimed to optimize the extraction of polysaccharides from Hericium coralloides by comparing the efficiencies of various extraction methods. They employed both traditional and modern methods, including:
- High-reflux extraction (HRE-P);
- Acid-base extraction;
- Enzymatic extraction;
- Ultrasonic-assisted extraction (UAE-P);
- Cold water extraction (CWE-P);
- Pressurized hot water extraction (PHE-P).
The researchers also investigated hydrogen peroxide/ascorbic acid systems (HAE-P) and acid–chlorite delignification.
Their results demonstrated significant intergroup differences in extract yield, chemical composition, monosaccharide profile, and molecular weight. All nine polysaccharide fractions obtained showed similar characteristics when analyzed by Fourier transform infrared (FTIR) spectroscopy and nuclear magnetic resonance (NMR) spectroscopy. Among the methods, high-reflux extraction produced the second-highest polysaccharide yield, as well as the highest polyphenol content and the highest molecular weight of polysaccharides. The metabolites obtained by high-reflux extraction demonstrated the most pronounced antioxidant activity against ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) and OH radicals. Cold water extraction revealed the highest antioxidant activity against ABTS, DPPH (2,2-diphenyl-1-picrylhydrazyl), and superoxide radicals, while ultrasonic-assisted extraction showed the best results against DPPH radicals [35].
Furthermore, the polysaccharides obtained by UAE-P, CWE-P, and HAE-P exhibited more pronounced cytoprotective activity against L929 cells compared to the other fractions. According to the correlation analysis, monosaccharide composition and total polyphenol content are key variables determining the bioactivity of H. coralloides polysaccharides [36].
Corallocins are a group of meroterpenoids that are benzofuranone and isoindolinone derivatives. Wittstein et al. [36] isolated three corallocins, namely A (2.8 mg), B (29.4 mg), and C (3.4 mg), from 7.88 g of crude ethyl acetate extract of H. coralloides fruiting bodies, whereas Ryu et al. [37] isolated corallocins from H. erinaceus fruiting bodies. Their chemical structures were examined by various spectral methods, including one-dimensional (1D) and two-dimensional (2D) nuclear magnetic resonance (NMR) spectroscopy, as well as high-resolution electrospray ionization mass spectrometry (HR-ESIMS) [37]. Subsequently, the same research group discovered two previously undescribed isoindolinone derivatives, which were named corallocin D and corallocin E. The crude ethyl acetate extract (272 mg) was purified by preparative reversed-phase liquid chromatography to isolate 1 mg of corallocin C and 0.7 mg of corallocin D.
The structures of these compounds were identified using HR-ESIMS and NMR analysis. The two compounds had a similar single chiral center and similar positive specific optical rotation values ([α]^20^D +24 and +26, respectively). Their absolute configuration was determined as an R-configuration by electron circular dichroism (ECD) [38].
The comparative structural analysis revealed that corallocins B–E have a common isoindolinone structure differing in the modification of the substituent at the nitrogen atom 2. Corallocin C, in turn, is a rare derivative of indoleisoindolinone. In contrast, corallocin A has a meroterpenoid structure of the benzofuranone type, with a carboxyl group located far from the geranyl fragment [23].
Studies have revealed significant neurotrophic potential of the compounds isolated from Hericium coralloides. Corallocins A, B, and C were examined for their ability to stimulate differentiation of PC12 cells (a rat pheochromocytoma cell line) by stimulating the astrocytoma cell line 1321N1 [36].
Corallocins A and C were found to increase the nerve growth factor (NGF) production in astrocytoma cells (line 1321N1) and stimulate neurite outgrowth in PC12 cells. Corallocins B and C also increased the expression of the brain-derived neurotrophic factor (BDNF) [33,37].
In another study, corallocin A isolated from H. erinaceus demonstrated its ability to induce strong survival and neurotrophic responses in cultured hippocampal neurons (DIV3) without the addition of serum [38]. Furthermore, corallocins D and E showed weak to moderate cytotoxicity against HeLa (KB 3.1), Mucor hiemalis, and Bacillus subtilis cells in vitro [39].
Lovastatin and ergothioneine (often referred to as the “longevity vitamins”) are valuable metabolites found in both the mycelia and fruiting bodies of Hericium fungi [40].
In [41], the scientists compared the contents of ergothioneine and lovastatin, as well as those of bioelements and glucan, in the fruiting bodies and mycelia of H. erinaceus, H. coralloides and H. americanum. They found that lovastatin was present in higher concentrations in the biomass obtained from the in vitro cultures compared to the fruiting bodies [23]. Among the three species, lovastatin had the highest content in the mycelium of H. coralloides (21.6 mg/100 g dry weight) and the lowest in that of H. erinaceus (5.81 mg/100 g dry weight). Unlike lovastatin, ergothioneine accumulated predominantly in the fruiting bodies of the fungi rather than in the in vitro cultures. In particular, its content was high in the fruiting bodies of H. erinaceus (305 mg/100 g dry weight) and lower in those of H. coralloides (155–177 mg/100 g dry weight) [41,42].
The presence of naturally occurring lovastatin in the biomass of H. coralloides adds further significance to this species as a potential source of hypolipidemic compounds. Inhibition of HMG-CoA reductase may contribute to reduced cholesterol biosynthesis and attenuation of systemic inflammation, which is clinically relevant in metabolic syndrome—a key risk factor for the development of NAFLD [43].
The main secondary metabolites of Hericium fungi are listed in Table 1.
The metabolites of Hericium fungi exhibit a wide range of biological activity. In particular, polysaccharides possess neuroprotective, immunomodulatory, and antioxidant properties, while erinacerins, hericerins, and other aromatic compounds exhibit neurotrophic and anticancer effects. Lovastatin and ergothioneine provide additional antioxidant, anti-inflammatory, and neuroprotective effects. Overall, the combination of these compounds makes the genus Hericium a promising source of bioactive substances for the development of functional foods and therapeutic agents [23].
In [33], the authors conducted a transcriptomic analysis to identify differential regulation of secondary metabolites of H. coralloides terpenoid fungi. The researchers used strain 77 of H. coralloides (Fujian, China). They found that this strain had dikaryotic hyphal morphology. The morphological analysis of its protoplasts (100× magnification) showed numerous clamp connections in the mycelium. These are mycelial bridges and anastomoses where fungal hyphae are tightly pressed or merge with each other, forming a stronger and more complex structure. These connections strengthen and support the mycelial strand, allowing the fungus to effectively transport nutrients throughout its network.
3.3. The Market of Dietary Supplements Containing Hericium coralloides (or Its Metabolites)
Dietary supplements containing Hericium coralloides and its metabolites lie at the intersection of nutraceuticals, functional mushrooms, and preventative medicine. Recent years have seen a growing interest in Hericium species as a source of compounds with neurotropic, anti-inflammatory, and metabolic-modulating properties [53]. Their potential hepatoprotective effects can help fight the increasing prevalence of non-alcoholic fatty liver disease (NAFLD). These effects of Hericium fungi are associated with their antioxidant activity, as well as their ability to reduce systemic inflammation and improve lipid and glucose metabolism.
The market of dietary supplements containing Hericium coralloides or its metabolites combines the interests in neurotropic, anti-inflammatory, and metabolically active natural compounds. These supplements are therefore considered as promising nutraceutical agents that can interact with the pathophysiological links of the disease and reduce the risk of its development in susceptible groups [54]. The products containing Hericium coralloides, which are available on the market, are presented in Table 2.
Most dietary supplements (Table 2) are based on dried and powdered H. coralloides mycelium or extract, rather than pure active metabolites. This leads to unstable concentrations of active ingredients and reduces the predictability of their effects. As a result, higher doses might be needed to achieve the desired result.
The analysis of the data collected during the literature review is reflected in Table 3.
The authors found no clinical studies confirming or refuting the effect of H. coralloides metabolites on NAFLD. They did find an in vivo study confirming the effect of ergothioneine on metabolic dysfunction-associated steatosis liver disease (MASLD) [71]. In the experiment, ergothioneine, administered daily at a dose of 35 mg/kg body weight to male mice C57BL/6J, improved lipid metabolism and reduced liver dysfunction in mice fed a high-fat diet.
The databases used in this study primarily include in vivo studies in rodents [46,66,72] and laying hens [19], which describe an indirect association with metabolic disorders, including NAFLD. In [19], the scientists studied the mechanisms by which polysaccharides isolated from H. erinaceus alleviate the symptoms of NAFLD in old laying hens, focusing on the regulatory function of the metabolites in the intestinal microbiome. They found that the polysaccharides ameliorated liver damage and metabolic disorders, as well as improved the intestinal barrier function. This reduced the transport of lipopolysaccharides from the intestine to the liver, inhibiting the activation of the hepatic pathway and slowing down the liver’s inflammatory response and apoptosis. Thus, H. erinaceus metabolites can mitigate liver damage and metabolic disorders associated with NAFLD by regulating the gut–liver axis [19].
These results demonstrate the value of further research examining the effects of H. coralloides metabolites on metabolic diseases in vitro, in vivo, and in humans. This literature review suggests that H. coralloides metabolites demonstrate potential for the prevention of NAFLD. Similar metabolites (atorvastatin, simvastatin) have also shown potential for the treatment of NAFLD in randomized, placebo-controlled trials [68].
Most of the data on the biological activity of H. coralloides metabolites were obtained in in vitro and experimental models of neurodegeneration, whereas direct models of NAFLD for this species are practically absent. Extrapolation of antioxidant and anti-inflammatory effects on liver tissue is possible only indirectly, through common pathogenetic mechanisms, including activation of the Nrf2 pathway, suppression of NF-kB, and decreased production of proinflammatory cytokines (TNF-α, IL-6), which play a key role in the progression of steatohepatitis. Similar mechanisms have previously been demonstrated for H. erinaceus polysaccharides in NAFLD models, which indicates the potential relevance of these pathways for H. coralloides, but requires direct experimental confirmation.
4. Conclusions
The data on Hericium coralloides metabolites suggest expanding the uses of this fungus as an ingredient in functional foods, nutraceuticals (dietary supplements), and cosmeceuticals due to its antioxidant properties. In addition to its traditional use as dried and powdered biomass in dietary supplements, the fungus’s individual bioactive metabolites (corallocins, erinacerins, and other neurotrophic compounds) could be introduced into functional foods and specialized dietary supplements. This will enable the scientists to standardize the composition of the products and increase their effectiveness, as well as to specifically implement their neuroprotective, immunomodulatory, and antioxidant effects for NAFLD prevention. The use of pure metabolites, or their concentrates, can open up new possibilities for developing products with proven pharmacological activity and more predictable and controllable biological effects compared to the traditional biomass [23].
Given the growing scientific interest in fungi of the genus Hericium, including the well-studied Hericium erinaceus [73], the biological potential of H. coralloides is relevant for both research and practical applications. Studying its composition, biological activity, cultivation methods, and technological applicability can contribute to expanding the range of functional ingredients and creating innovative nutraceutical products. This review covered the chemical composition, nutritional value, and biological activity of H. coralloides, as well as its potential use in the food industry, specifically in products supporting metabolic health and liver function.
Significant limitations of the current evidence base should be noted, including the lack of direct in vivo models of NAFLD using H. coralloides specifically, as well as the lack of clinical studies evaluating the effect of its metabolites on biochemical markers of liver function. Therefore, the use of this mushroom in nutraceutical products should be considered as a potentially preventive rather than therapeutic intervention. Future research will focus on developing standardized extracts with known levels of polysaccharides, ergothioneine, and terpenoids, using cell lines, animal models of NAFLD, and conducting randomized clinical trials evaluating biochemical and imaging markers of steatosis.
At the initial stage, in vitro studies will be conducted using cell lines relevant to the pathogenesis of NAFLD and metabolic disorders, including human hepatocyte cell lines (HepG2 and Huh7) and macrophage cell lines (THP-1) to evaluate anti-inflammatory activity. The investigations will focus on the regulation of genes involved in lipid metabolism (PPARα, SREBP-1c, FAS), oxidative stress (Nrf2, SOD, CAT), inflammatory responses (NF-κB, TNF-α, IL-6), and programmed cell death pathways, including apoptosis and autophagy (caspase-3). In addition, the effects of extracts and individual metabolites on mitochondrial function, reactive oxygen species levels, and cellular insulin sensitivity will be assessed. At the next stage, in vivo studies using animal models of NAFLD are considered appropriate, including diet-induced models (high-fat diet-fed mice or rats), which allow for the reproduction of key metabolic and morphological features of the disease.
Thus, the combination of studies on cell lines and validated animal models will enable the substantiation of the biological activity of Hericium coralloides at the level of molecular mechanisms and provide a scientific basis for the subsequent transition to clinical studies and the development of nutraceutical products with proven preventive potential against NAFLD and related metabolic disorders.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cheremukhina Y.V. Borovkova N.Y. Vasilkova A.S. Vlasova T.V. Akhmedzhanov N.M. Non-alcoholic fatty liver disease: A view formation on the problem and its significance for the clinical practice Russ. J. Prev. Med.202528127132(In Russian)10.17116/profmed 202528071127 · doi ↗
- 2Kytikova O.Y. Novgorodtseva T.P. Denisenko Y.K. Kovalevsky D.A. Metabolic and Genetic Determinants of Lipid Metabolism Disruption in Non-Alcoholic Fatty Liver Disease Russ. J. Gastroenterol. Hepatol. Coloproctol.202030152510.22416/1382-4376-2020-30-2-15-25 · doi ↗
- 3Meroni M. Longo M. Rustichelli A. Dongiovanni P. Nutrition and Genetics in NAFLD: The Perfect Binomium Int. J. Mol. Sci.202021298610.3390/ijms 2108298632340286 PMC 7215858 · doi ↗ · pubmed ↗
- 4Reshetova M.S. Zol’nikova O.Y. Ivashkin V.T. Ivashkin K.V. Appolonova S.A. Lapina T.L. Rol’ kishechnoj mik-robioty i ee metabolitov v patogeneze nealkogol’noj zhirovoj bolezni pecheni Ross. Zhurnal Gastroentero-Logii Gepatologii Koloproktol.2022327588(In Russian)10.22416/1382-4376-2022-32-5-75-88 · doi ↗
- 5Chekushkina D.Y. Fedorova A.M. Kovalenko S.V. Milentyeva I.S. Altshuler O.G. Aksenova L.M. Anti-Metabolic Syndrome Effect of Trans-Cinnamic Acid Food Process. Tech. Technol.20255513614710.21603/2074-9414-2025-1-2563 · doi ↗
- 6Kosobyan E.P. Smirnova O.M. Modern concepts of pathogenesis of non-alcoholic fatty liver disease Diabetes Mellit.2010135564
- 7Vesnina A. Le V. Ivanova S. Prosekov A. Antidiabetic Potential of Mangiferin: An In Silico and In Vivo Approach Pharmaceutics 202517126210.3390/pharmaceutics 1710126241155899 PMC 12567361 · doi ↗ · pubmed ↗
- 8Chalasani N. Younossi Z. Lavine J.E. Charlton M. Cusi K. Rinella M. Harrison S.A. Brunt E.M. Sanyal A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases Hepatology 20186732835710.1002/hep.2936728714183 · doi ↗ · pubmed ↗