Bioinformatic Prediction of Activation States in Molecular Network Pathways of Eukaryotic Initiation Factor 2 (EIF2) Signaling and Coronavirus Pathogenesis
Shihori Tanabe, Sabina Quader, Ryuichi Ono, Hiroyoshi Y. Tanaka, Horacio Cabral

TL;DR
This paper explores how the EIF2 signaling pathway interacts with coronavirus infection, finding that they have opposite activation states and share key molecules.
Contribution
The study identifies inverse activation states and overlapping molecular components between EIF2 signaling and coronavirus pathogenesis.
Findings
EIF2 signaling and coronavirus pathogenesis have inverse activation states.
EIF2 signaling interacts with multiple miRNAs like let-7 and miR-1292-3p.
Eight molecules and one complex overlap between the two pathways.
Abstract
Eukaryotic initiation factor 2 (EIF2) signaling plays a crucial role in regulating mRNA translation and initiating eukaryotic protein synthesis. Computational molecular network pathway analysis of the canonical pathways of the coronaviral infection revealed that EIF2 signaling is inactivated when the coronavirus pathogenesis pathway is activated and vice versa. Our computational analyses indicated that the coronavirus pathogenesis pathway and EIF2 signaling had inverse activation states. Computational investigation of upstream or downstream microRNA (miRNA) revealed that EIF2 signaling directly interacted with miRNAs, including let-7, miR-1292-3p (miRNAs with the seed CGCGCCC), miR-15, miR-34, miR-378, miR-493, miR-497, miR-7, miR-8, and MIRLET7. A total of 36 nodes, including 8 molecules (ATF4, BCL2, CCND1, DDIT3, EIF2A, EIF2AK3, EIF4E, and ERK1/2), 1 complex (the ribosomal 40s…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
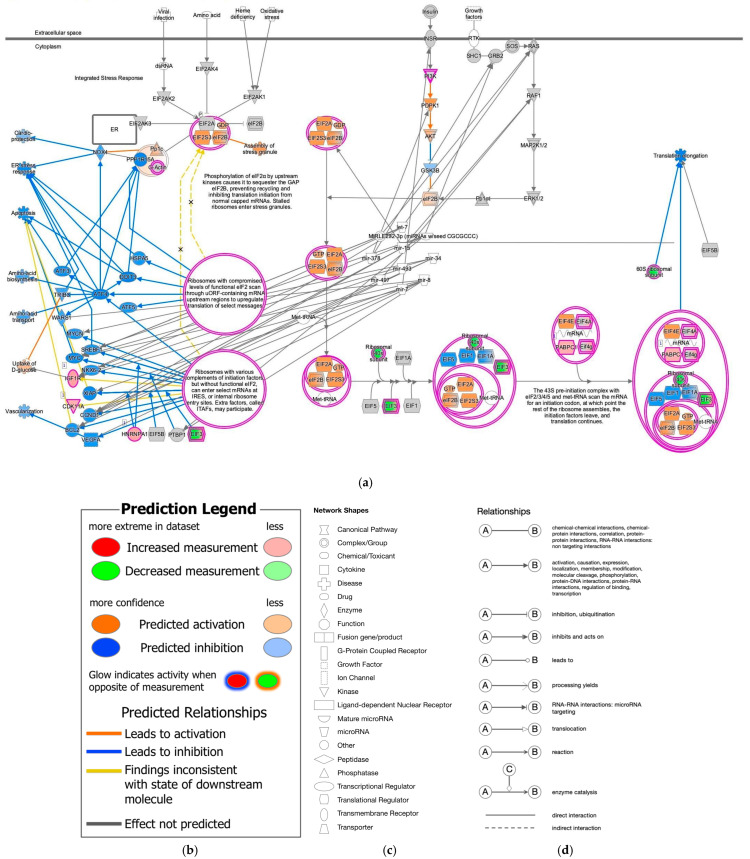 Figure 1
Figure 1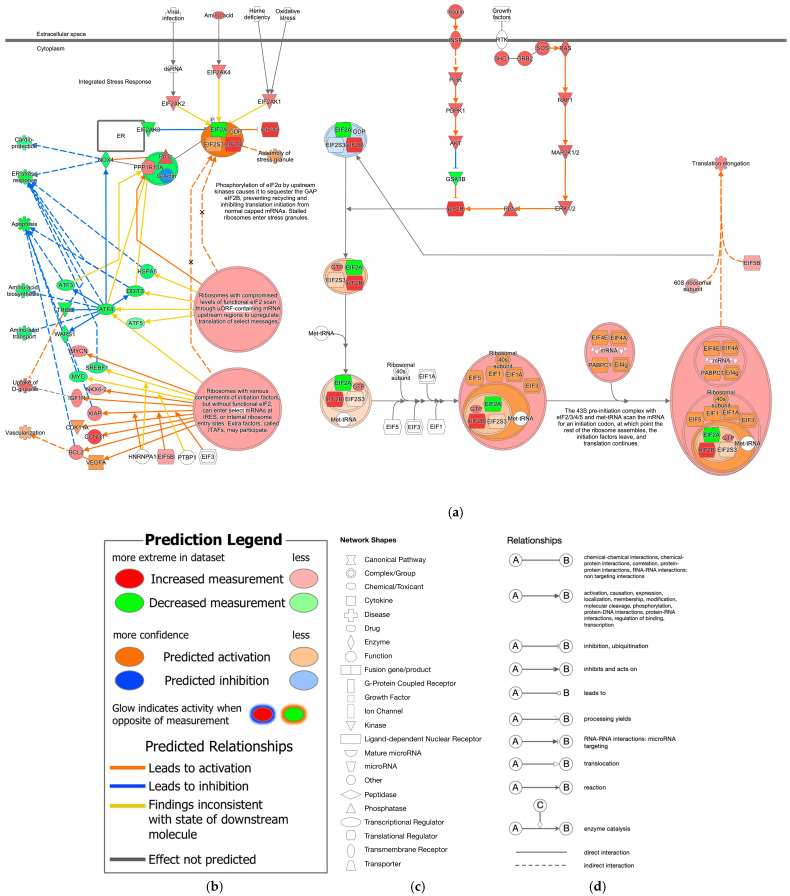 Figure 2
Figure 2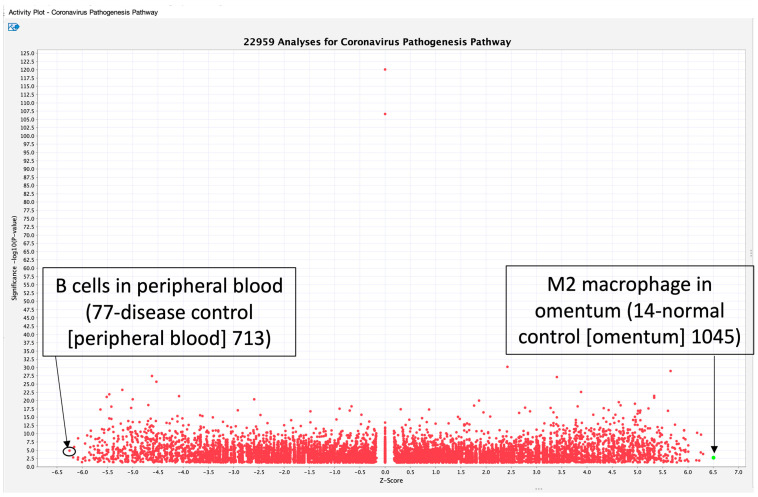 Figure 3
Figure 3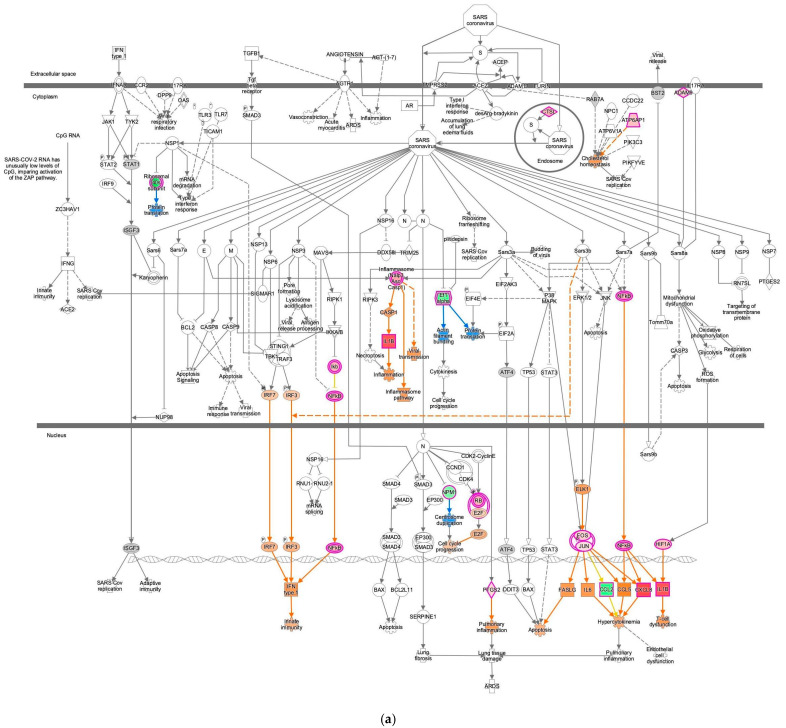 Figure 4
Figure 4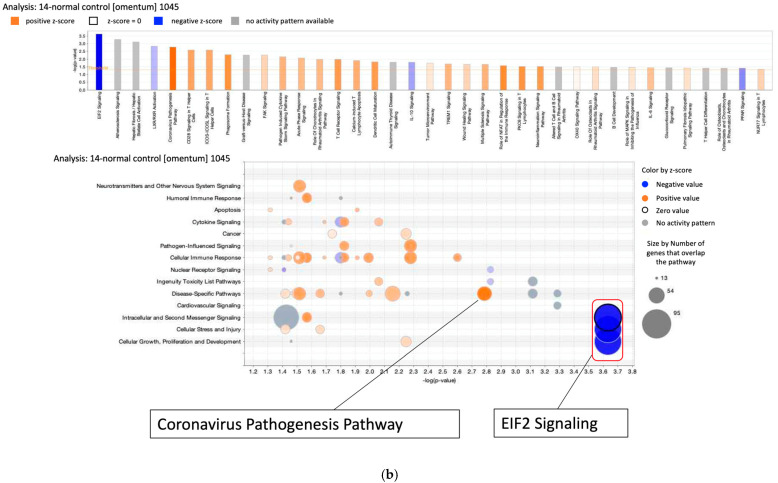 Figure 5
Figure 5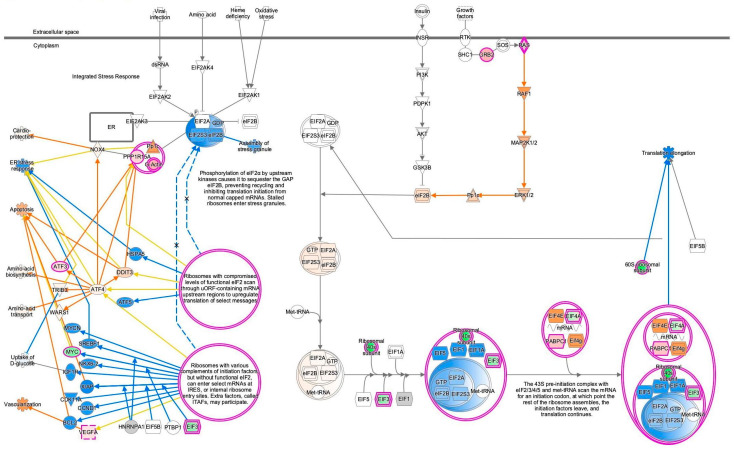 Figure 6
Figure 6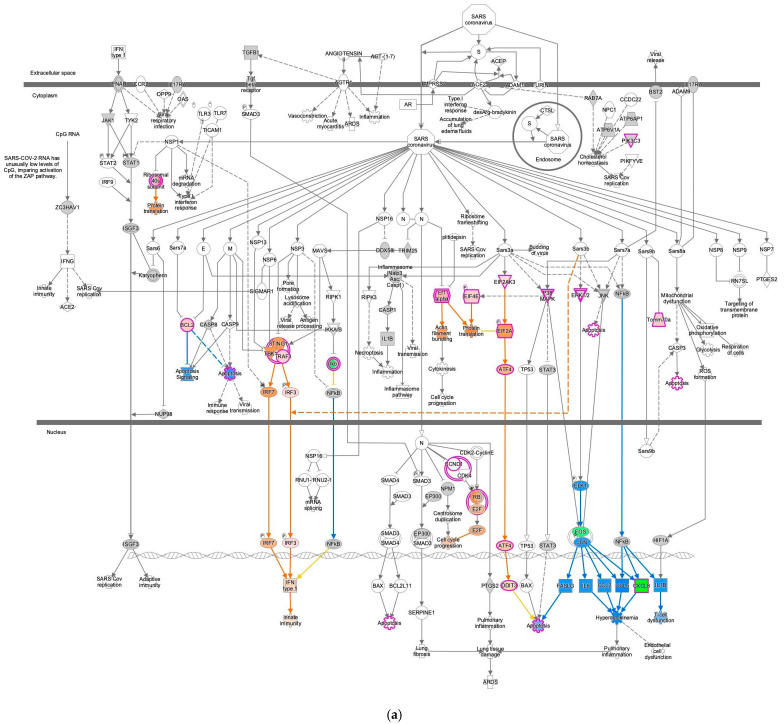 Figure 7
Figure 7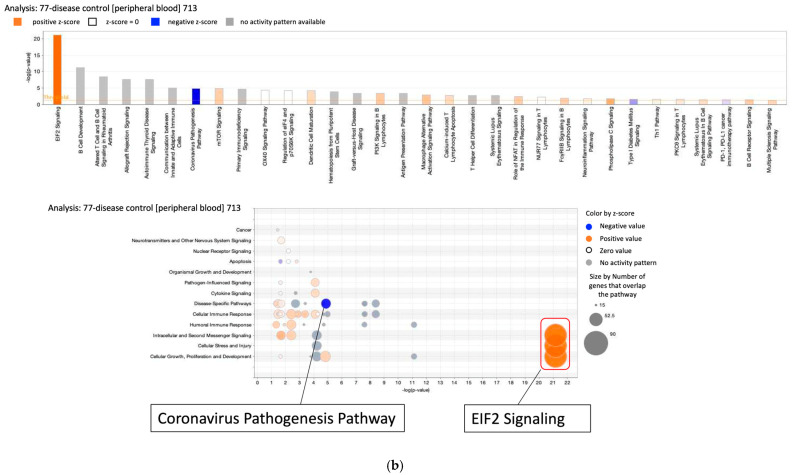 Figure 8
Figure 8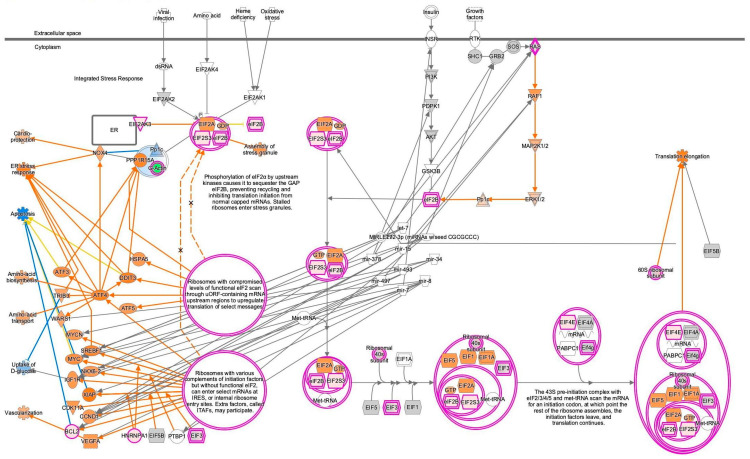 Figure 9
Figure 9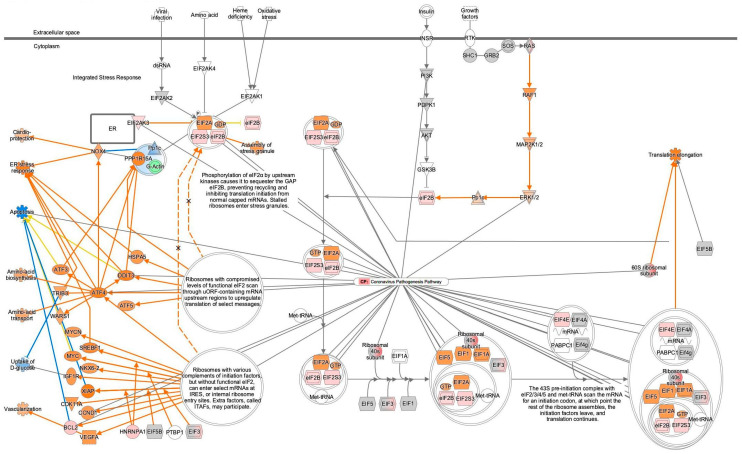 Figure 10
Figure 10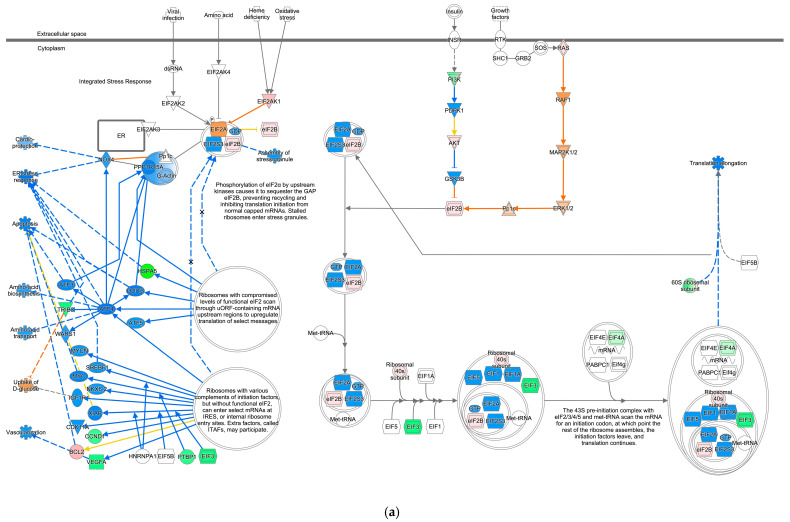 Figure 11
Figure 11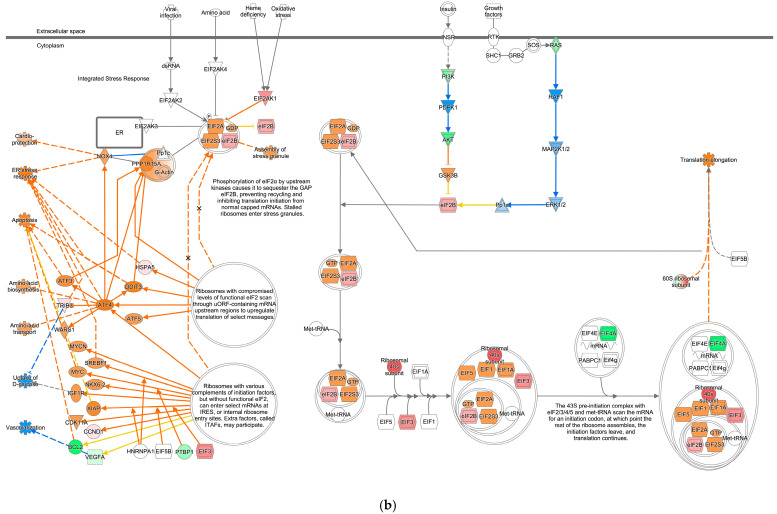 Figure 12
Figure 12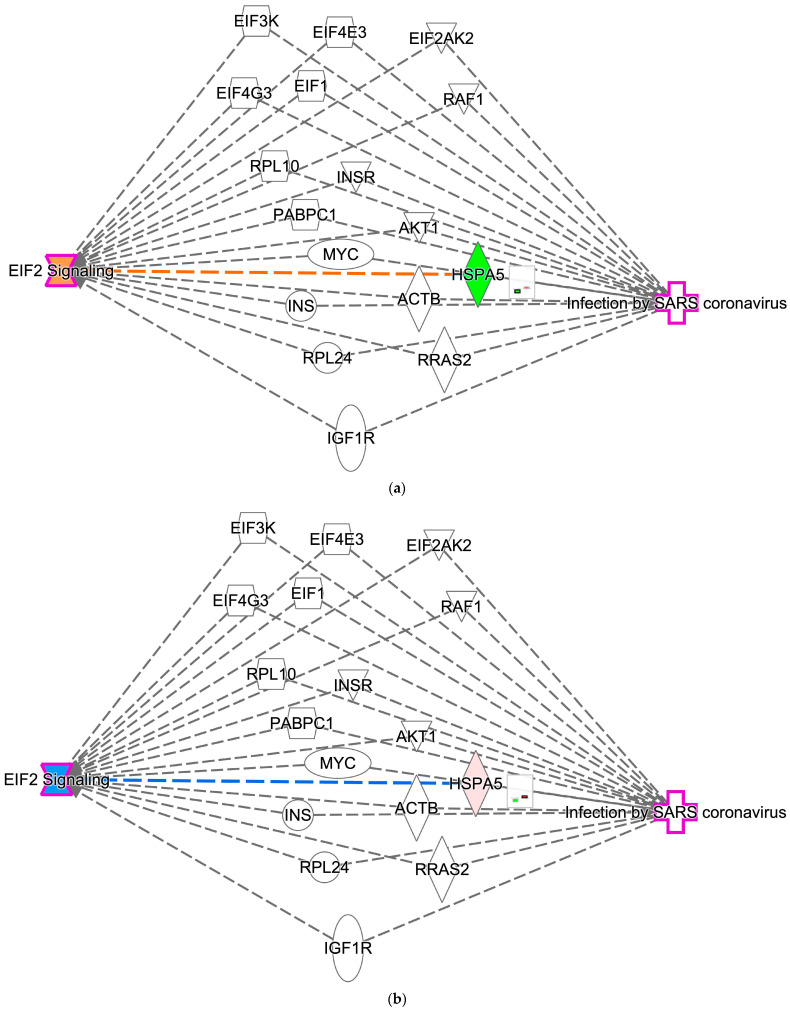 Figure 13
Figure 13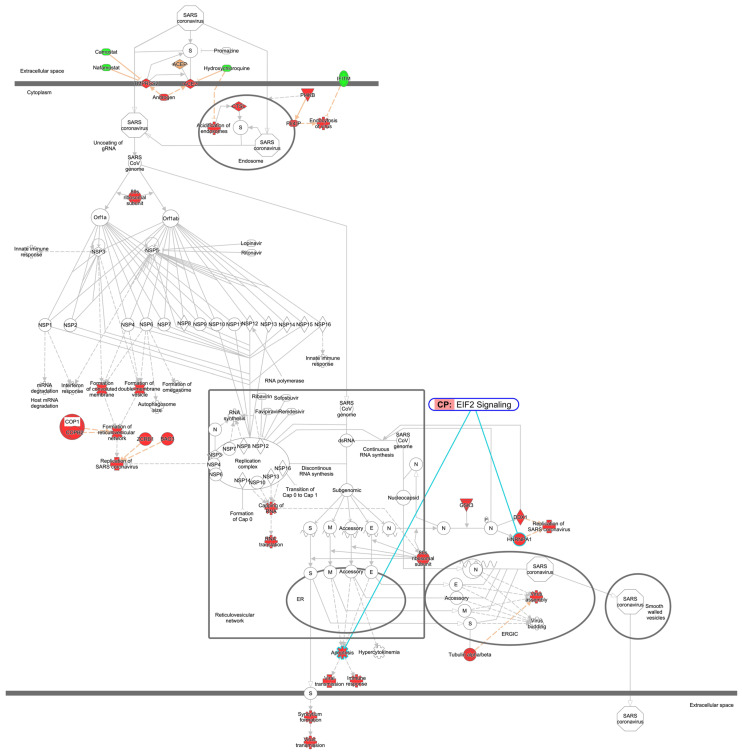 Figure 14
Figure 14- —Japan Agency for Medical Research and Development (AMED)
- —Japan Society for the Promotion of Science (JSPS)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMachine Learning in Bioinformatics · RNA regulation and disease · RNA and protein synthesis mechanisms
1. Introduction
Eukaryotic translation initiation factor 2 (EIF2) is a guanosine 5′-diphosphate (GDP)-binding protein with three subunits (alpha, beta, and gamma) that delivers initiator tRNA (Met-tRNAi) to 40S ribosomes in a guanosine 5′-triphosphate (GTP)-dependent manner [1]. EIF2 signaling contributes physiologically to the initiation of protein synthesis and the integrated stress response (ISR) [2]. The ISR involves a ternary complex composed of EIF2, GTP, and charged Met-tRNAi [3] and is triggered by various stress conditions that alter cellular homeostasis [2]. Sensor kinases HRI (heme-regulated inhibitor, encoded by the EIF2 alpha kinase 1 (EIF2AK1) gene), PKR (double-stranded RNA-dependent protein kinase, encoded by the EIF2AK2 gene), and PERK (PKR-like endoplasmic reticulum kinase, encoded by the EIF2AK3 gene), in addition to GCN2 (general control non-repressible 2, encoded by the EIF2AK4 gene), contain both conserved kinase domains and divergent regulatory domains that enable them to respond to different stimuli [2].
The coronavirus pathogenesis pathway is activated upon coronaviral infection. The coronavirus pathogenesis pathway involves mitogen-activated protein kinase (MAPK) signaling, interferon (IFN) signaling, transforming growth factor (TGF)-beta signaling, and nuclear factor (NF)-kappa B signaling, leading to the inhibition of innate immunity and the dysfunction of T cells [4]. Although the coronavirus pathogenesis and coronavirus replication pathways include known signaling pathways, their precise mechanisms and regulation of activity have yet to be revealed. In this study, we focused on EIF2 signaling as a hub and compared the activation states of the EIF2 signaling and the coronavirus pathogenesis pathway using the Ingenuity Pathway Analysis (IPA) computational network pathway analysis tool and gene expression databases. We investigated whether the EIF2 signaling pathway and the coronavirus pathogenesis pathway are inversely activated in public gene expression datasets, defined their integration with miRNA interactions, and conducted a cross-tissue comparison of EIF2 signaling and the coronavirus pathogenesis pathway.
2. Results
2.1. EIF2 Signaling in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)-Infected Induced Pluripotent Stem (iPS) Cells
During our previous study on the coronavirus pathogenesis pathway [4], we found that EIF2 signaling was inactivated in SARS-CoV-2-infected induced pluripotent stem (iPS) cells. In the computational analysis of SARS-CoV-2-infected iPS cells, the gene expression of EIF3 (complex) was decreased, and EIF1, EIF1A, and EIF5 were predicted to be inhibited (Figure 1). Translation elongation was predicted to be inhibited in SARS-CoV-2-infected iPS cells, as shown in Figure 1. EIF2A, EIF2S3, and EIF2B (complex) (eIF2B) were predicted to be activated in the EIF2 signaling of SARS-CoV-2-infected iPS cells. The source dataset of the computational analysis is specified in Table 1. The Gene Expression Omnibus (GEO) accession number of the source dataset is GSE156754 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE156754, accessed on 17 December 2025) [5].
EIF2B (complex) consists of EIF2B1, EIF2B2, EIF2B3, EIF2B4, and EIF2B5 and is a member of 43S TRANSLATION PREINITIATION (complex), 48S RIBOSOME (complex), EIF2 (complex), and MET/TRNA/EIF2 (complex). EIF2B (complex) is involved in the cachexia signaling pathway, cardiac hypertrophy signaling, cardiac hypertrophy signaling (enhanced), estrogen receptor signaling, eukaryotic translation initiation, the IFN-gamma-activated inhibitor of translation (GAIT) signaling pathway, insulin receptor signaling, the insulin secretion signaling pathway, the regulation of EIF4 and p70S6K signaling, and the stress granule signaling pathway, in addition to EIF2 signaling in the IPA canonical pathway knowledge base. Key molecules in EIF2 signaling are summarized in Table 2.
2.2. Activation State of EIF2 Signaling
EIF2 signaling and the expected activation state of EIF2 signaling in IPA are shown in Figure 2. Amino acid activates EIF2AK4, leading to the assembly of stress granules, while RAS signaling activates eIF2B, leading to the initiation of translation by the 43S preinitiation complex with EIF2/3/4/5 and Met-tRNA (Figure 2). The expected activation state of EIF2 signaling contained several types of nodes, including increased/decreased measurement and predicted activation/inhibition in computational pathway analysis.
2.3. Activity Plots of the Coronavirus Pathogenesis Pathway
To reveal the activation states of the coronavirus pathogenesis pathway under various changes in gene expression, activity plots of the coronavirus pathogenesis pathway were analyzed using the IPA database to identify data with the highest or lowest activation z-score. Activity plots of the coronavirus pathogenesis pathway are shown in Figure 3. A total of 22,959 analyses were identified in coronavirus pathogenesis pathway activity plots. The 22,959 analyses were defined using a canonical pathway search with the term “Coronavirus Pathogenesis Pathway” and by selecting the activity plot option under the optional license of IPA computational pathway analysis as of 17 October 2022. At that time, the number of gene expression data analyses relating to the coronavirus pathogenesis pathway increased. The datasets (analyses) in the IPA database under the optional license, which included the coronavirus pathogenesis pathway as an associated canonical pathway as of 17 October 2022, were computationally selected. Detailed information on datasets that have absolute z-scores greater than 3 is available in Supplementary Table S1. M2 macrophages in the omentum had the highest activation z-score, indicating that the gene expression pattern of the molecules in the dataset is similar to the pattern expected based on the available literature (the coronavirus pathogenesis pathway activity plot as of October 2022 (Figure 3)). The gene expression of M2 macrophages in the dataset was compared to that of B cells, dendritic cells, fibroblasts, mast cells, mesothelial cells, stromal cells, and T cells (Figure 3). An analysis of B cells in peripheral blood showed the lowest z-score in coronavirus pathogenesis pathway activity plots as of October 2022 (Figure 3). In the dataset, gene expression in B cells was compared to that in monocytes, neutrophils, platelets, T cells, and unassigned cells, indicating that the gene expression pattern of B cells is less similar to the pattern expected based on the available data on the coronavirus pathogenesis pathway compared to other cells in peripheral blood (Figure 3).
2.4. EIF2 Signaling Inhibition of the M2 Macrophage in Omentum with the Most Activated Coronavirus Pathogenesis Pathway
The coronavirus pathogenesis pathway was activated in M2 macrophages in the omentum (Figure 4). The result for 14-normal control (omentum) 1045 was overlaid onto the coronavirus pathogenesis pathway, which showed activation of the coronavirus pathogenesis pathway in computational pathway analysis (Figure 4). The same analysis result was overlaid onto the EIF2 signaling pathway, which showed inactivation of the EIF2 signaling (Figure 4b and Figure 5). In the 14-normal control (omentum) 1045 analysis, the M2 macrophages were compared to other cell types, such as plasma B cells, dendritic cells, fibroblasts, mast cells, mesothelial cells, stromal cells, and T cells, in the male omentum tissue of the normal control in the IPA database. In the IPA analysis of miRNA interactions with EIF2 signaling, nine microRNAs (miRNAs) (let-7, miR-1292-3p (miRNAs w/seed CGCGCCC), miR-15, miR-34, miR-378, miR-493, miR-497, miR-7, and miR-8) and one group (MIRLET7) were identified as having direct relationships with EIF2 signaling (Table 3). The computational analysis was conducted purely using knowledge-based prediction as part of IPA; this hypothesis-generating prediction is the limitation of this study that will require validation in future investigations.
2.5. Enrichment Analysis of the Predicted Target Genes of microRNAs (miRNAs)
To further investigate the roles of miRNAs associated with EIF2 signaling, enrichment analysis was performed on the predicted target genes of the miRNAs. The predicted target genes of the nine miRNAs and one group indicated in Table 3 that have direct relationships with EIF2 signaling are shown in Table 4 and Supplementary Table S2.
The target genes were uploaded to the Database for Annotation, Visualization, and Integrated Discovery (DAVID), which resulted in the analysis of 528 Homo sapiens genes. Functional annotation clustering of the enrichment analysis showed that chronic myeloid leukemia, hepatitis B, cellular senescence, human T-cell leukemia virus 1 infection, and viral carcinogenesis were included in the top cluster with the highest enrichment scores (Table 5, Supplementary Table S3).
2.6. EIF2 Signaling Activation in B Cells in Peripheral Blood with the Least Activated Coronavirus Pathogenesis Pathway
Computational pathway analysis suggested reduced coronavirus pathogenesis pathway activity in peripheral blood B cells (Figure 6). The result for the 77-disease control (peripheral blood) 713 analysis was overlaid onto the coronavirus pathogenesis pathway, which showed inactivation of the coronavirus pathogenesis pathway (Figure 6). The same analysis result was overlaid onto EIF2 signaling, and this showed increased EIF2 signaling activity in computational pathway analysis (Figure 6b and Figure 7). The disease state of the 77-disease control (peripheral blood) 713 analysis was normal. The gene expression data were obtained from adult female peripheral blood B cells, and other cells (monocytes, neutrophils, platelets, T cells, and unassigned cells) were used for comparison in the analysis performed using the IPA database. The data source is available in ArrayExpress with Accession number E-MTAB-9221 (https://www.ebi.ac.uk/biostudies/arrayexpress/studies/E-MTAB-9221, accessed on 18 November 2025) [6]. Detailed information of the 77-disease control (peripheral blood) 713 analysis is also presented in Supplementary Table S1.
A total of 36 nodes, including 8 molecules (ATF4, BCL2, CCND1, DDIT3, EIF2A, EIF2AK3, EIF4E, and ERK1/2), 1 complex (the ribosomal 40s subunit), and 1 function (apoptosis) in coronavirus pathogenesis, were overlapped with EIF2 signaling (Figure 8, Table 6).
2.7. EIF2 Signaling in Diffuse-Type and Intestinal-Type Gastric Cancer
To investigate whether the inverse relationship between the activation states of the coronavirus pathogenesis pathway and EIF2 signaling holds in other disease states, we analyzed diffuse-type and intestinal-type gastric cancer, in which we had previously observed the differential dysregulation of the coronavirus pathogenesis pathway.
EIF2 signaling was overlaid onto gene expression in diffuse-type and intestinal-type gastric cancer (Figure 9). Most of the nodes of the EIF2 signaling pathway in diffuse-type gastric cancer were predicted to be inhibited (Figure 9a), whereas most of the nodes in intestinal-type gastric cancer were predicted to be activated (Figure 9b) in the EIF2 signaling pathway. Translation elongation was predicted to be inhibited or activated in diffuse-type or intestinal-type gastric cancer, respectively, in computational pathway analysis. Uptake of D-glucose was predicted to be activated or inhibited in diffuse-type or intestinal-type gastric cancer, respectively. Vascularization was predicted to be inhibited in both diffuse-type and intestinal-type gastric cancer in computational pathway analysis.
2.8. Interaction Between EIF2 Signaling and Infection by SARS-CoV
The interaction between EIF2 signaling and SARS-CoV infection was analyzed using IPA. The gene expression overlay for diffuse-type gastric cancer in relation to the interaction of EIF2 signaling and infection by SARS-CoV revealed EIF2 signaling activation and a decrease in heat shock protein family A (Hsp70) member 5 (HSPA5) in computational analysis (Figure 10a). The gene expression overlay for intestinal-type gastric cancer in relation to the interaction of EIF2 signaling and infection by SARS-CoV revealed EIF2 signaling inhibition and an increase in HSPA5 in computational analysis (Figure 10b).
The nodes that interacted with EIF2 signaling and infection by SARS-CoV included actin beta (ACTB), AKT serine/threonine kinase 1 (AKT1), eukaryotic translation initiation factor 1 (EIF1), eukaryotic translation initiation factor 2 alpha kinase 2 (EIF2AK2), eukaryotic translation initiation factor 3 subunit K (EIF3K), eukaryotic translation initiation factor 4E family member 3 (EIF4E3), eukaryotic translation initiation factor 4 gamma 3 (EIF4G3), heat shock protein family A (Hsp70) member 5 (HSPA5), insulin-like growth factor 1 receptor (IGF1R), insulin (INS), insulin receptor (INSR), MYC proto-oncogene, bHLH transcription factor (MYC), poly(A) binding protein cytoplasmic 1 (PABPC1), Raf-1 proto-oncogene, serine/threonine kinase (RAF1), ribosomal protein L10 (RPL10), ribosomal protein L24 (RPL24), and RAS-related 2 (RRAS2). Among the nodes linking EIF2 signaling and infection by SARS coronavirus, HSPA5 was identified as the gene in which gene expression was altered in diffuse-type and intestinal-type gastric cancer.
2.9. EIF2 Signaling in Coronavirus Replication Pathway
The EIF2 signaling and the coronavirus replication pathway are associated through heterogenous nuclear ribonucleoprotein A1 (HNRNPA1) and apoptosis (Figure 11). HNRNPA1 is a member of a family of ubiquitously expressed heterogeneous nuclear ribonucleoproteins, which are RNA-binding proteins that associate with pre-mRNAs in the nucleus and influence pre-mRNA processing, as well as other aspects of mRNA metabolism and transport.
3. Discussion
Molecular network analysis using the IPA network analysis tool revealed that the coronavirus pathogenesis pathway and EIF2 signaling pathway have inverse activation states. EIF2 signaling is involved in the process of protein translation. Major factors in EIF2 signaling include EIF2A, EIF2 subunit gamma (EIF2S3), eIF2B (EIF2B (complex)), and EIF2AK2. The level of phosphorylation of EIF2A is elevated in hypoxia via the induction of ER stress and PERK [7]. EIF2 signaling may be involved in various physiological conditions, including tumor progression, viral infection, amino acid starvation, heme deprivation, proteasome inhibition, and UV irradiation [7,8].
EIF2AK2 (or protein kinase double-stranded RNA-dependent; PKR) was initially identified as a kinase that phosphorylates EIF2A in response to viral infection [8]. EIF2AK2 plays important roles in eukaryotic response to viral infection, the regulation of cell growth and differentiation, apoptosis, and cancer [9]. EIF2 signaling and virus infection are closely correlated. Recent studies have demonstrated that viral infection causes an alteration in EIF2 signaling pathways [10,11,12,13,14,15]. SARS-CoV-2 infection induces beta cell transdifferentiation via the EIF2 pathway, which is mediated by PKR phosphorylation [12]. EIF2 was suggested as a potential target for reversing beta cell transdifferentiation in diabetes following SARS-CoV-2 infection [12]. Human coronavirus (HCoV) infection downregulates the EIF2 signaling pathway [10], which aligns with the findings of our study. SARS-CoV-2 nonstructural protein (NSP) 6 can trigger EIF2AK3-EIF2A pathway-mediated autophagy and inhibit interferon production [11]. Upon detection of cytoplasmic double-stranded RNA (dsRNA), mammalian cells initiate pathways including the EIF2AK2/PKR pathway, the oligo(A) synthetase–ribonuclease L (OAS-RNase L) pathway, and the RIG-I-like receptor–mitochondrial antiviral-signaling pathway [13]. Upon binding to viral dsRNA, EIF2AK2/PKR forms homodimers, which induce the autophosphorylation of Thr446 (phospho-PKR) within the activation segment of its catalytic domain and activate EIF2AK2/PKR catalytic activity [13]. EIF2AK2/PKR is a key player in the innate immune response to RNA virus infection that upregulates antiviral gene expression, including the production of interferons [16]. Coronaviruses counteract EIF2AK2/PKR-mediated signaling in order to prevent the translational shut-off due to EIF2A phosphorylation [16].
Viruses enter cells via cellular receptors and use the protein translation machinery of the infected host cells to replicate viral proteins [17,18]. Viral infection triggers cellular ISR, in which the regulation of the EIF2 pathway plays a vital role as an antiviral immune system [17]. It is likely that viral replication induces mitochondrial gene transcription and production, and mitochondrial dsRNAs, a potential major source of endogenous dsRNAs in mammalian cells, activate PKR and subsequently phosphorylate EIF2A, resulting in the inhibition of host protein translation [17,19]. The systemic inverse relationship between the coronavirus pathogenesis pathway and EIF2 signaling observed in the current study would explain why EIF2 signaling inhibition is triggered by the host antiviral immune system. It is also possible that the virus suppresses the host protein translation to promote the translation of viral proteins.
Upregulated genes in diabetes gastroenteropathy include the SARS-CoV-2 viral entry genes and genes involved in viral replication [14]. The mouse hepatitis coronavirus induces the phosphorylation of EIF2A, which leads to the formation of stress granules and processing bodies and, subsequently, host mRNA translational shut-off [20]. EIF2AK3/PERK is an ER-localized transmembrane unfolded protein response (UPR) sensor with a luminal amino-terminal domain that binds HSPA5 [15]. The UPR reduces ER load via the EIF2AK3-dependent arrest of cap-dependent translation initiation and by increasing ER-associated catabolic activities. Here, reticulophagy, a catabolic process in which ER cargo is diverted to the lysosome for degradation, plays an essential role in maintaining ER homeostasis [15]. Structural glycoprotein expression in Zaire ebolavirus is decreased during viral infection to increase the virus’s fitness via reticulophagy [21]. The expression of glycoprotein induces ER stress, which activates the UPR to stall protein translation, thereby reducing the protein load in the ER and activating the gene expression of proteins involved in protein folding [21]. EIF2AK3/PERK in the ER transmembrane activates the transcription factor ATF4 during UPR in ER stress [21].
M2 macrophages are involved in anti-inflammatory responses and allergic diseases [22,23]. The characteristics of M2 macrophages may resemble the activated state of the coronavirus pathogenesis pathway. Computational pathway analysis predicted that EIF2 signaling was activated, while the coronavirus pathogenesis pathway was inactivated in B cells. Some immune modulation may be involved, and this requires future investigation. Among the nodes linking EIF2 signaling and infection by SARS coronavirus, HSPA5 was altered in diffuse-type and intestinal-type gastric cancer. Since Hsp70 is involved in stress response, and caloric restriction increased the expression of Hsp70 in the gut [24], disease onset and malignancy might be associated with translational regulation.
There is also the possibility of knowledge base bias or circularity, with the selection of coronaviruses already known to manipulate EIF2. It is possible that EIF2 nodes are downstream of stress or viral proteins. In future investigations, orthogonal validation approaches (e.g., gene set enrichment analysis or independent dataset reanalysis) may be needed to ensure the inverse pattern is not an artifact of pathway definition.
EIF2 signaling plays a crucial role in translation initiation and beta cell transdifferentiation following severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection [12]. Our findings indicate that EIF2 signaling is inactivated when the coronavirus pathogenesis pathway is activated. The difference between coronavirus and SARS-CoV-2 might cause this discrepancy. In the current study, we identified multiple miRNAs (e.g., miR-378, let-7 family members, miR-34, miR-1292-3p, etc.) that interact with the EIF2 signaling pathway. An miRNA analysis in COVID-19 patients revealed that let-7 was significantly downregulated in the plasma of acute-phase COVID-19 patients [25]. Another study also demonstrated that the let-7 family was downregulated in the plasma of both mild and severe COVID-19 patients compared to healthy controls [26]. Another study demonstrated that let-7 was upregulated in the serum and urine of moderate COVID-19 cases, but not in mild or severe COVID-19 cases, compared to healthy controls [27]. These previous experimental findings indicate that some regulation in the let-7 family of miRNAs is involved in coronavirus pathogenesis. Indeed, a small-molecule C1632, which blocks the interaction between LIN28 and pri/pre-let-7, thus promoting the maturation of let-7, was reported to inhibit SARS-CoV-2 replication and virus-induced inflammation via upregulating let-7 [28]. The target genes of miRNAs identified were involved in viral signaling.
A limitation of this study is that the interactions were analyzed in silico; thus, experimental confirmation is required. Comprehensive quantitative analyses are needed to capture the overall relationship between EIF2 signaling and coronavirus pathogenesis pathways. This could involve the following: (i) a scatter plot of EIF2 signaling vs. the coronavirus pathogenesis pathway across all analyses, including correlation coefficients and the proportion of analyses with opposite activation directions in the activity plot; (ii) measurement of EIF2 phosphorylation and translation rates in primary M2 macrophages or B cells during coronavirus infection; (iii) perturbation of selected miRNAs, such as the let-7 family, and assessment of EIF2 pathway readouts; and (iv) validation of the EIF2–coronavirus pathway interplay in gastric organoid models. The EIF2 signaling pathway is a potential target of viral infection, as viral infection and EIF2 signaling interact closely.
4. Materials and Methods
4.1. Canonical Pathway Analysis
The coronavirus pathogenesis pathway and EIF2 signaling were analyzed using the QIAGEN IPA network pathway tool [4,29] (https://digitalinsights.qiagen.com) (QIAGEN Digital Insights, Aarhus C, Denmark, and Redwood City, CA, USA) (accessed on 22 September 2025). As of 2021, 106 analyses and 106 datasets from more than 100,000 pieces of data were found to be related to SARS-CoV in the IPA database as previously described [4]. The details of activation z-scores were described in a previous study [4]. We focused on the relationship between the coronavirus pathogenesis pathway and EIF2 signaling in the current study.
4.2. Activity Plot Analysis
The activity plot of the coronavirus pathogenesis pathway activation z-score in the IPA database identified more than 20,000 analyses as of October 2022, which is many more than previously described [4]. Among the analyses of the coronavirus pathogenesis pathway in the IPA database, the M2 macrophage in the omentum had the highest z-score, and B cells in peripheral blood had the lowest z-score, as of October 2022.
4.3. miRNA Interaction Analysis
The upstream or downstream miRNA with direct interactions with the nodes in the canonical pathways was investigated. The confidence level of the miRNA was set as experimentally observed or high (predicted) in the QIAGEN IPA setting.
4.4. Enrichment Analysis
The enrichment analysis of the predicted target genes of the miRNAs was performed using the Database for Annotation, Visualization, and Integrated Discovery (DAVID) (https://davidbioinformatics.nih.gov/home.jsp, accessed on 15 December 2025). The predicted target genes of the miRNAs were extracted from the IPA database search. The predicted target genes of the miRNAs were then uploaded to DAVID, which resulted in the enrichment analysis of 528 Homo sapiens genes (9 genes were identified as unknown). Functional annotation clustering was performed using DAVID.
4.5. IPA Network Analysis
The analysis results for iPS cells infected with SARS-CoV-2 are available in the IPA database. The source dataset is available in the GEO public database with the GEO accession number GSE156754 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE156754, accessed on 17 December 2025) [5]. Data on intestinal-type and diffuse-type gastric cancer from The Cancer Genome Atlas (TCGA) of the cBioPortal for Cancer Genomics database (https://www.cbioportal.org, accessed on 30 September 2025) at the National Cancer Institute (NCI) Genomic Data Commons (GDC) data portal (https://portal.gdc.cancer.gov, accessed on 30 September 2025) [30,31,32,33] were uploaded and analyzed using IPA, as previously described [34,35].
4.6. Statistical Analysis
The RNA sequencing data on diffuse-type and intestinal-type gastric cancer were analyzed using IPA, as previously described [36]. The activation z-score in each network or pathway was calculated using IPA to show the level of activation [29].
5. Conclusions
The molecular network analysis revealed that the coronavirus pathogenesis pathway and EIF2 signaling frequently exhibit inverse activation states in the examined datasets. Interestingly, when the coronavirus pathogenesis pathway is activated, EIF2 signaling is inactivated, and when EIF2 signaling is activated, the coronavirus pathogenesis pathway is inactivated according to computational pathway analysis. The findings of the study may provide insight into treatments for coronavirus infection.
While the computational prediction is a valid approach for a hypothesis-generating study, experimental validation is critical to overcome inherent limitations. Future investigations—for example, validation of key protein levels, such as phosphorylated EIF2A or PKR, or testing the role of identified miRNAs in coronavirus-infected cell models—are needed to validate the computational predictions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Suragani R.N. Ghosh S. Ehtesham N.Z. Ramaiah K.V. Expression and purification of the subunits of human translational initiation factor 2 (e IF 2): Phosphorylation of e IF 2α and βProtein Expr. Purif.20064722523310.1016/j.pep.2005.10.00316289913 · doi ↗ · pubmed ↗
- 2Costa-Mattioli M. Walter P. The integrated stress response: From mechanism to disease Science 2020368 eaat 531410.1126/science.aat 531432327570 PMC 8997189 · doi ↗ · pubmed ↗
- 3Milne G.L. Yin H. Hardy K.D. Davies S.S. Roberts L.J. Isoprostane Generation and Function Chem. Rev.20111115973599610.1021/cr 200160 h 21848345 PMC 3192249 · doi ↗ · pubmed ↗
- 4Tanabe S. Quader S. Ono R. Tanaka H.Y. Yamamoto A. Kojima M. Perkins E.J. Cabral H. Artificial Intelligence Approach in Machine Learning-Based Modeling and Networking of the Coronavirus Pathogenesis Pathway Curr. Issues Mol. Biol.20254746610.3390/cimb 4706046640699865 PMC 12191508 · doi ↗ · pubmed ↗
- 5Perez-Bermejo J.A. Kang S. Rockwood S.J. Simoneau C.R. Joy D.A. Silva A.C. Ramadoss G.N. Flanigan W.R. Fozouni P. Li H. SARS-Co V-2 infection of human i PSC–derived cardiac cells reflects cytopathic features in hearts of patients with COVID-19Sci. Transl. Med.202113 eabf 7872.10.1126/scitranslmed.abf 787233723017 PMC 8128284 · doi ↗ · pubmed ↗
- 6Silvin A. Chapuis N. Dunsmore G. Goubet A.-G. Dubuisson A. Derosa L. Almire C. Hénon C. Kosmider O. Droin N. Elevated Calprotectin and Abnormal Myeloid Cell Subsets Discriminate Severe from Mild COVID-19Cell 202018214011418.e 1810.1016/j.cell.2020.08.00232810439 PMC 7405878 · doi ↗ · pubmed ↗
- 7Rozpedek W. Pytel D. Mucha B. Leszczynska H. Diehl J. Majsterek I. The Role of the PERK/e IF 2α/ATF 4/CHOP Signaling Pathway in Tumor Progression During Endoplasmic Reticulum Stress Curr. Mol. Med.20161653354410.2174/156652401666616052314393727211800 PMC 5008685 · doi ↗ · pubmed ↗
- 8Donnelly N. Gorman A.M. Gupta S. Samali A. The e IF 2α kinases: Their structures and functions Cell. Mol. Life Sci.2013703493351110.1007/s 00018-012-1252-623354059 PMC 11113696 · doi ↗ · pubmed ↗