Molecular Mechanisms of NF-Y Transcription Factors in Horticultural Plant Development and Stress Responses: Recent Advances
Mengxia Zhang, Dan Chen, Chunjuan Dong

TL;DR
This review explores how NF-Y transcription factors regulate growth and stress responses in horticultural plants, offering insights for improving plant resilience and productivity.
Contribution
The paper provides a comprehensive overview of recent advances in understanding NF-Y molecular mechanisms in horticultural plants.
Findings
NF-Ys regulate plant development by integrating signals like GA and ABA and controlling downstream genes.
They enhance stress tolerance by managing ROS, ABA biosynthesis, and defense pathways.
Species-specific genomic and structural traits contribute to NF-Y specialization and function.
Abstract
Nuclear Factor Y (NF-Y) transcription factors are evolutionarily conserved regulators that bind the CCAAT box, playing central roles in horticultural plant growth and adaptation. This review summarizes recent progress on NF-Ys in horticultural plants, focusing on their molecular mechanisms in development and stress responses. For development, NF-Ys mediate phase transition, flowering regulation, embryogenesis, and organ development by integrating endogenous signals (gibberellic acid, GA; abscisic acid, ABA) and regulating downstream genes. For stress responses, they enhance tolerance to abiotic stresses (drought, salt, extreme temperatures) via regulating reactive oxygen species (ROS) scavenging, ABA biosynthesis, and stress networks, and mediate biotic stress resistance (e.g., pathogen infection) by activating defense pathways. This review also briefly covers species-specific genomic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
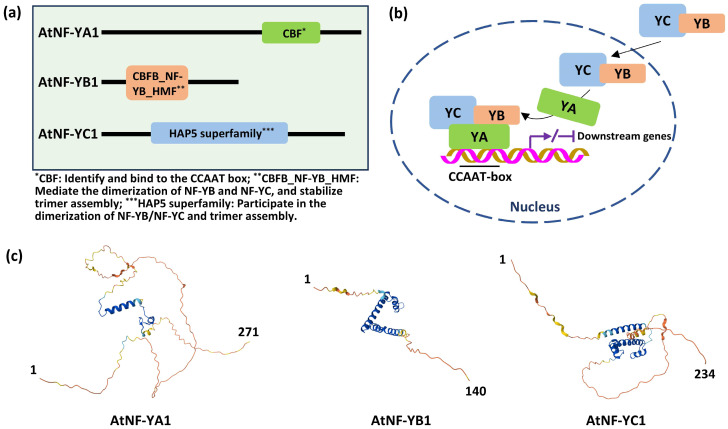 Figure 1
Figure 1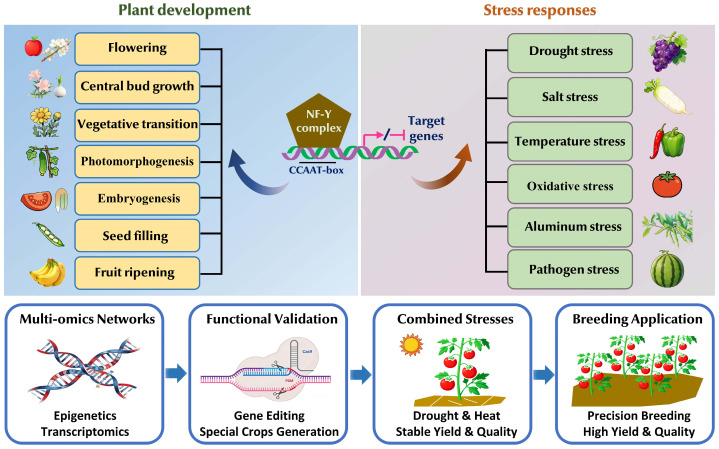 Figure 2
Figure 2- —National Key R&D Program of China
- —Research Startup Funding from Hainan Institute of Zhejiang University
- —National Natural Science Foundation of China
- —Science and Technology Innovation Project of the Chinese Academy of Agricultural Sciences
- —Key Laboratory of Biology and Genetic Improvement of Horticultural Crops (Vegetables), Ministry of Agriculture and Rural Affairs
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Gene Expression Analysis · Polysaccharides and Plant Cell Walls
1. Introduction
Horticultural plants, encompassing fruits, vegetables, ornamental flowers, and medicinal herbs, are vital components of global food security, economic sustainability, and ecological balance. They contribute significantly to human nutrition, aesthetic value, and agricultural diversification, while facing relentless challenges from abiotic stresses (e.g., drought, salt, extreme temperatures) and biotic stresses (e.g., pathogen infections) amid global climate change. Additionally, precise regulation of developmental processes—such as phase transition, flowering, seed maturation, and fruit ripening—is critical for optimizing crop yield, quality, and commercial value. Transcription factors (TFs) are key regulatory proteins that orchestrate gene expression networks, integrating environmental cues and endogenous signals to govern plant development and stress adaptation.
Among diverse TF families, NF-Y is a conserved heterotrimeric complex (NF-YA/NF-YB/NF-YC) that specifically binds to the CCAAT cis-element in target gene promoters to modulate transcriptional initiation [1,2,3]. Initially characterized in yeast and animals, NF-Ys have been extensively studied in model plants for their pleiotropic roles in development and stress tolerance [4,5,6]; with advances in high-throughput genome sequencing and functional genomics, NF-Y family members have since been systematically identified in an increasing number of horticultural plants, revealing key species-specific traits—including variations in subunit gene numbers, uneven chromosome distribution, and family expansion driven by gene duplication events [7,8,9]—as well as structural features: conserved core domains ensure trimer assembly and DNA-binding specificity, while variable N/C termini enable species-specific regulatory functions [10,11,12].
Horticultural plants possess a unique set of agronomically vital traits—such as fruit ripening, tuber/bulb organogenesis, ornamental flower development, and perennial growth cycles—that are not central to model plant studies yet are primary targets for yield and quality improvement. Despite substantial progress in dissecting NF-Y functions in individual horticultural species (e.g., tomato, apple, cucumber) [13,14,15,16], a comprehensive synthesis of their molecular mechanisms underlying these and other horticulture-specific processes as well as stress responses across diverse horticultural taxa remains lacking. Moreover, the crosstalk between NF-Y-mediated pathways and other regulatory networks (e.g., hormone signaling, miRNA regulation, epigenetic modifications) is not fully elucidated [17,18,19,20], and the translational potential of NF-Y factors for horticultural crop improvement is yet to be fully exploited. Currently, research on NF-Y in ornamental and medicinal plants remains fragmented. Many species have only undergone genome-wide identification of NF-Y genes, with functional characterization still lacking. This represents a significant knowledge gap and a promising direction for future research, particularly in exploring species-specific adaptation mechanisms and secondary metabolic regulation. In this review, we systematically summarize recent advances in the genomic organization, structural features, and molecular mechanisms of NF-Y TFs in horticultural plant development (phase transition, flowering, organogenesis) and stress responses (abiotic and biotic stresses) [1,13,14,15,21,22]. We also highlight knowledge gaps and propose future research directions, aiming to provide a holistic overview for researchers in this field and facilitate the application of NF-Y-related biotechnology in horticultural crop breeding.
This review systematically evaluates studies that advance our mechanistic understanding of NF-Ys in horticultural plants. While prioritizing recent findings from the past five years, it also incorporates foundational earlier work that remains critical to the field. We systematically searched databases including Web of Science, Scopus, PubMed, and Google Scholar using keywords such as “NF-Y”, “horticultural plants”, “stress response”, “development”, and relevant species names. The literature screening prioritized functional studies focusing on the roles of NF-Y in horticultural plant development and stress adaptation, ensuring the relevance and reliability of the included research.
2. Genomic Organization and Structural Features of NF-Y Families in Horticultural Plants
NF-Y transcription factors are conserved heterotrimeric complexes (NF-YA/NF-YB/NF-YC) that regulate horticultural plant growth, development, and stress responses [1,2,3]. With genome sequencing advances, their genomic organization and structural traits have been widely studied, revealing species-specific diversity.
2.1. Genomic Organization of NF-Y Families in Horticultural Plants
The genomic organization of the NF-Y family in horticultural plants exhibits distinct characteristics, primarily reflected in variations in subunit gene numbers, uneven chromosome distribution, and expansion driven by gene duplication events.
Regarding subunit gene numbers, significant interspecific differences exist, largely shaped by evolutionary processes such as whole-genome duplication (WGD) [3]. For instance, tomato (Solanum lycopersicum) harbors a large NF-Y family with 10 NF-YA, 29 NF-YB, and 20 NF-YC genes [15,16], while butterfly orchid (Phalaenopsis sp.) has a smaller family (4 NF-YA, 9 NF-YB, 11 NF-YC) [23]. Apple (Malus domestica), which has experienced multiple WGD events, contains 11 NF-YA, 22 NF-YB, and 10 NF-YC genes [13], highlighting duplication-mediated family expansion that supports adaptive functional differentiation.
NF-Y genes are unevenly distributed across chromosomes, with non-random patterns often linked to functional clustering. Cucumber (Cucumis sativus) NF-Y genes are present on all chromosomes except chromosome 2, with the highest density on chromosomes 3 and 6 (7 genes each) [14]; Pepper (Capsicum annuum) has 19 NF-YB genes on 7 of its 12 chromosomes, including tandem duplications on chromosome 7 [22]; and Japanese apricot (Prunus mume) NF-Y genes concentrate on chromosomes 01 and 03 (25.92% each) but are rare on chromosome 06 (3.70%) [24].
Gene duplication—encompassing WGD, segmental duplication, and tandem duplication—is a key driver of NF-Y family expansion. The large NF-Y family of banana (Musa acuminata), consisting of 14 NF-YA, 16 NF-YB, and 14 NF-YC subunits, is primarily driven by WGD [25]. For woodland strawberry (Fragaria vesca), segmental duplication underpins the distribution of NF-Y genes across its 7 chromosomes [21]. In pepper, tandem duplication leads to the clustering of NF-YB genes on chromosome 7 [22]. These duplication events facilitate functional differentiation of NF-Y genes, thereby enhancing plants’ adaptability to various stresses.
Detailed genomic organization data of NF-Y families and their key functional roles studied across various horticultural plants are summarized in Table 1.
2.2. Structural Features of NF-Y Subunits in Horticultural Plants
The function of the NF-Y trimer relies on conserved domains in each subunit. NF-YA contains a CCAAT-binding domain (CBF) that enables specific binding to target gene promoters [1,10], while its N/C termini interact with the NF-YB/NF-YC heterodimer [50]. NF-YB has a histone fold motif (YB_HMF) mediating dimerization with NF-YC, a structure resembling the histone H2A-H2B complex [10]. NF-YC harbors a Heme Activator Protein 5 (HAP5) superfamily domain that stabilizes NF-YB/NF-YC dimerization via hydrophobic interactions [11].
The trimer assembly follows a strict sequence: NF-YB and NF-YC first form a cytoplasmic heterodimer through their histone fold motifs, translocate to the nucleus, and then bind NF-YA to form a functional trimer [50]. Only the complete trimer binds DNA—NF-YA recognizes the CCAAT box, and the NF-YB/NF-YC heterodimer stabilizes this DNA interaction [10]. Notably, higher plants have retained more NF-Y subunit paralogs during evolution, and the diverse trimeric combinations derived from these paralogs endow NF-Y complexes with functional specificity—supporting their involvement in various plant developmental processes and responses to different stresses (e.g., drought, salt stresses and fungal infections) [2,3,31]. This combinatorial flexibility, built on the conserved assembly sequence, is considered a core mechanism that accounts for the multifaceted roles of NF-Y in plant adaptation and growth [51].
Core domains (e.g., histone fold motifs in NF-YB/NF-YC) are conserved across horticultural species like tomato, apple, and cucumber [13,14,15,16]. In contrast, variable N/C termini support species-specific functions: NF-YA in sweet orange (Citrus sinensis) has drought-responsive motifs [35], and NF-Y subunits in banana carry motifs related to fruit ripening [25]. The conserved core domains of NF-Y subunits underpin the universal assembly and DNA-binding specificity of the heterotrimeric complex across horticultural species. In contrast, the divergent N- and C-terminal regions provide the structural basis for functional diversification. This modular architecture—a conserved “engine” coupled with variable “regulatory modules”—enables the evolution of a large repertoire of NF-Y complexes through combinatorial associations of paralogous subunits. Consequently, the functional specificity of a given NF-Y in processes such as drought response or fruit ripening likely depends not only on its expression pattern but also on the precise subunit composition of the trimer, which may determine interactions with specific co-regulators or chromatin contexts.
Figure 1 takes three typical NF-Y subunits (AtNF-YA1/AtNF-YB1/AtNF-YC1) from Arabidopsis as examples to illustrate key structural details of NF-Y subunits—including their domain architectures, the NF-Y trimer-mediated CCAAT-box-dependent transcriptional regulation process, and the AlphaFold-predicted 3D structures of each subunit—all of which underpin the conserved domains and assembly mechanism of NF-Y subunits discussed above [52,53].
3. Molecular Mechanisms of NF-Ys in Horticultural Plant Development
Recent studies have elucidated diverse and important roles of NF-Y transcription factors in horticultural plant development. Their functions are particularly critical in two key, interconnected areas: the regulation of phase transition and flowering time (Section 3.1), and the coordination of early embryogenesis, organ morphogenesis, and seed maturation alongside storage compound accumulation (Section 3.2). The underlying molecular mechanisms for these processes are increasingly elucidated. This chapter synthesizes the latest advances in these two pivotal research areas, highlighting how NF-Y complexes integrate diverse signals to direct developmental fate and resource allocation.
3.1. Phase Transition and Flowering Control
NF-Y transcription factors function as central integrators within the gene regulatory networks that govern phase transition and flowering time in horticultural plants. They decode a combination of endogenous developmental signals, such as GA and ABA, and environmental cues, including photoperiod and temperature, to precisely regulate the expression of key flowering pathway genes like FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1). A recurring theme emerging from these studies is the role of NF-Y complexes as integrators of hormone signals—particularly GA and ABA—with environmental and developmental cues to fine-tune flowering time. This section synthesizes recent advances that elucidate how specific NF-Y heterotrimeric complexes—through direct promoter binding, interactions with other transcription factors, and integration with hormonal and epigenetic pathways—orchestrate the precise timing of flowering across diverse horticultural species.
NF-Y transcription factors play pivotal roles in phase transition and flowering control of horticultural plants by mediating target gene expression and signal crosstalk. Li et al. (2025) identified 57 BcNF-Y members in non-heading Chinese cabbage (Brassica campestris), and BcNF-YA8, upregulated by ABA, promotes flowering via directly activating BcFT expression and inhibiting ascorbate accumulation [17]. Conversely, Zhang et al. (2024) reported that tomato SlNF-YA3b negatively regulates flowering by binding to the CCAAT cis-elements of SFT (FT homolog) promoter, with its knockout inducing early flowering [54]. Pan et al. (2023) demonstrated that lily (Lilium spp.) LoNFYA7 recruits Lilium oriental VERNALIZATION INSENSITIVE 3-LIKE 1 (LoVIL1)- Polycomb Repressive Complex 2 (PRC2) to enhance H3K27me3 at Lilium oriental CALLOSE SYNTHASE 3 (LoCALS3) locus, repressing callose synthesis and facilitating bud growth transition from dormancy [19]. In summary, NF-Ys exert diverse regulatory effects on phase transition and flowering through distinct molecular pathways. More details about NF-Y transcription factors that regulate phase transition and flowering in horticultural plants are summarized in Table 2.
The collected evidence positions NF-Ys as versatile, context-dependent regulators of flowering, capable of both promotion and repression (Table 2). This functional duality may be resolved by considering three key factors. First, subunit specificity: distinct NF-YA/B/C combinations likely confer unique target gene selectivity. Second, upstream signal integration: the same NF-Y can mediate opposite outcomes depending on whether it is activated by ABA, GA, or other signaling pathways. Third, experimental resolution: while some studies establish direct regulation through promoter binding and genetic interaction, others report correlative evidence. Thus, NF-Ys likely function as integrative nodes within flowering networks, with their output finely tuned by developmental stage, environmental cues, and specific protein partnerships.
3.2. Early Embryogenesis, Organ Morphogenesis, and Seed Maturation with Storage Compound Accumulation
Beyond flowering, NF-Y complexes are master regulators of organogenesis and maturation, largely through their capacity to coordinate hormone dynamics (e.g., auxin, ABA, GA, ethylene) and metabolic pathways. They direct the development of agriculturally vital organs and storage tissues by forming heterotrimeric complexes and modulating target gene expression. Specifically, NF-Y transcription factors regulate horticultural plant development—encompassing early embryogenesis, organ morphogenesis, seed maturation, and storage compound accumulation—primarily by forming heterotrimeric complexes and modulating target gene expression, with regulatory roles involving hormone signaling, chloroplast biogenesis, and metabolic pathways. In tomato, SlLEC1-LIKE4 (SlL1L4) harbors a unique N-terminal domain that integrates auxin and ABA signals to coordinate cell division, tissue patterning, and desiccation tolerance critical for early embryogenesis, with ZFN-mediated disruption leading to abnormal embryonic development, altered seed storage compound accumulation and delayed fruit ripening [67]. For cucumber, CsNF-YC2 and CsNF-YC9 directly bind the promoter of Cucumis sativus TRANSLOCON AT THE INNER MEMBRANE OF CHLOROPLASTS 21 (CsTIC21) to enhance its transcription, thereby promoting light-dependent chloroplast photomorphogenesis and leaf development [68]. In potato (Solanum tuberosum), the StNF-YA8-YB20-YC5 module activates genes related to GA synthesis and ABA catabolism, altering the ABA/GA balance to accelerate tuber dormancy release, a key process in stolon and tuber organogenesis [69]. For watermelon (Citrullus lanatus), ClNF-YB9 interacts with ClNF-YCs and recruits ClNF-YA7 to form a functional trimer, which is critical for embryo morphogenesis and seedling organ formation [33]. In tomato, SlNF-YA8 modulates cotyledon development during embryogenesis and seed maturation, with ZFN-mediated disruption leading to altered seedling establishment, 43% increased fruit weight, rounded fruit shape, and modified locule number that reflect its pleiotropic roles in linking early embryonic development to seed maturation and post-embryonic organ formation [58]. Together, these findings underscore the conserved and diverse roles of NF-Y transcription factors across the sequential processes of early embryogenesis, organ morphogenesis, and seed maturation, with detailed information on related genes and mechanisms summarized in Table 3.
Beyond their ancestral role in embryogenesis, NF-Ys have been co-opted for the regulation of horticulturally vital processes, such as fruit ripening, tuber development, and seed storage compound accumulation. This functional expansion in horticultural species often involves conserved regulators like LEC1 (NF-YB9) but targets distinct downstream genes affecting texture, color, flavor, and nutrition [33,34,70]. The mechanistic studies, particularly in tomato and potato, reveal that NF-Ys interface with hormone (ethylene, ABA) and sugar signaling networks to control these specialized developmental programs. However, the extent to which the underlying regulatory logic is conserved between, for example, fruit ripening and seed maturation, or is newly evolved in specific lineages, remains an open and compelling question.
4. Molecular Mechanisms of NF-Ys in Horticultural Plant Stress Responses
Horticultural plants are frequently exposed to diverse abiotic stresses (e.g., drought, salt, extreme temperatures) and biotic stresses (e.g., pathogen infections) during growth and development, which severely affect their yield and quality. As key transcriptional regulators, NF-Y factors play pivotal roles in mediating stress-responsive signaling pathways by binding to CCAAT elements in target promoters and coordinating with other regulatory factors to modulate gene expression. This section systematically summarizes recent advances in the molecular mechanisms underlying NF-Y-mediated stress tolerance in horticultural plants, with a focus on major abiotic and biotic stress types.
4.1. Drought Stress
NF-Y transcription factors function as integrative hubs that coordinate multiple hormone signaling pathways to mediate drought adaptation. This integrative function is exemplified by their central role in amplifying ABA signaling—a common hub in drought response—while simultaneously interacting with other hormone pathways such as jasmonate (JA) to orchestrate a comprehensive adaptive output. The drought response mediated by NF-Ys predominantly converges on the amplification of ABA signaling and the enhancement of cellular antioxidant capacity. Drought stress severely restricts the growth, yield, and quality of horticultural plants, with NF- Y transcription factors constituting key components of these pathways. In apple (Malus hupehensis), the drought- and ABA-induced NF-YA subunit MhNF-YA3-like enhances drought tolerance by directly binding the promoter of the ABA biosynthesis gene Malus hupehensis Arabidopsis ALDEHYDE OXIDASE 3 (MhAAO3) to activate its expression, and its interaction with Malus hupehensis MULTICOPY SUPPRESSOR OF IRA1 4-LIKE (MhMSI4-like) further amplifies this effect—overexpression reduces leaf wilting and ROS accumulation, while silencing increases susceptibility [76]. In chrysanthemum (Chrysanthemum morifolium), the negatively acting CmNF-YB8 is downregulated by drought; silencing it upregulates Chrysanthemum morifolium CBL-INTERACTING PROTEIN KINASE 6 (CmCIPK6, involved in stomatal adjustment) and Chrysanthemum morifolium SHINE 3 (CmSHN3, involved in cuticle biosynthesis) to reduce water loss, whereas overexpression exacerbates drought damage [77]. In citrus (Citrus spp.), drought-induced CiNF-YA1 forms a complex with CiNF-YB2/CiNF-YC2 to activate CiFT (mediating drought-induced flowering), and while overexpression increases drought susceptibility via ROS accumulation, silencing enhances tolerance through improved photosynthesis [78]. These studies demonstrate the diverse roles of NF-Ys in horticultural plant drought responses, with the involved mechanisms—hormone biosynthesis, stomatal adjustment, cuticle development, and flowering crosstalk—being only a subset of their regulatory modes. More NF-Y-mediated drought response mechanisms in horticultural plants are summarized in Table 4.
A conserved theme emerging from studies across species is the central role of NF-Ys in amplifying the ABA signaling pathway to orchestrate drought responses, including stomatal closure and expression of protective genes (e.g., LEAs, HSPs) [18,86,87]. Functional validation for this role is strong in cases like potato StNF-YA7 and sweet orange CsNF-YA5 [18,35,84,85]. However, the landscape of evidence is mixed, with several entries supported primarily by expression correlation. Notably, species-specific adaptations exist alongside this ABA core; for instance, CmNF-YB8 in chrysanthemum regulates cuticle morphology, a mechanism less emphasized in other species [77]. This indicates that while a central ABA-connected module is widely utilized, NF-Y networks also contribute to lineage-specific adaptive traits.
4.2. Salt Stress
In response to salt stress, which severely impacts horticultural plant growth, NF-Y transcription factors have been widely shown to regulate gene networks essential for salt tolerance. In grapevine (Vitis amurensis), VaNF-YA6 enhances salt tolerance when overexpressed in grapevine leaves and Arabidopsis by upregulating stress-responsive genes (e.g., VvSOS2, VvSOS3) and improving antioxidant enzyme activities [81]. In tomato, SlNF-YC1 interacts with SlMYB1 to strengthen the transcriptional activation of the glutamate decarboxylase gene SlGAD1, promoting γ-aminobutyric acid (GABA) accumulation and ROS scavenging, thereby enhancing saline-alkali tolerance [88]. Overexpression of CsNF-YC6 from tea plant (Camellia sinensis) in Arabidopsis (CsNF-YC6-OE transgenic lines) resulted in increased seed germination and root length under ABA and GA treatments, as well as improved salt stress tolerance by regulating ABA signaling-related genes and increasing proline content [27]. These findings underscore the diverse mechanisms of NF-Ys in mediating salt stress responses, including gene regulation, metabolite accumulation, and antioxidant defense. For a comprehensive summary of NF-Ys involved in salt stress responses in horticultural plants, refer to Table 5.
NF-Ys enhance salt tolerance through two primary, often interconnected, strategies: ionic homeostasis (e.g., regulating SOS pathway genes as seen in grapevine VaNF-YA6 [81]) and osmotic adjustment (e.g., promoting compatible solute accumulation like GABA in tomato via SINF-YC1 [88]). The relative emphasis on these strategies may reflect ecological adaptation. A critical examination reveals that the evidence for direct transcriptional control of ion transporters by NF-Ys is less prevalent than for their role in osmotic stress signaling. Furthermore, the frequent induction of NF-YA genes by salt stress, often via downregulation of miR169, presents a conserved regulatory layer, yet the functional significance of specific miR169-targeted NF-YA paralogs in salt tolerance warrants deeper investigation across species [32,89].
4.3. Temperature Stress
4.3.1. Heat Stress
Heat stress significantly impairs horticultural plant growth, with NF-Y transcription factors orchestrating stress responses via complex regulatory networks. In cucumber, CsNFYA1 interacts with Cucumis sativus MULTIPROTEIN BRIDGING FACTOR 1c (CsMBF1c) to enhance heat tolerance by stabilizing the photosynthetic system and promoting transcriptional activation of heat-related genes [90]. In cabbage (Brassica oleracea), Brassica oleracea DNA POLYMERASE II SUBUNIT B3-1 (BoDPB3-1)/NF-YC10 acts as a coactivator of DEHYDRATION-RESPONSIVE ELEMENT BINDING PROTEIN 2A (DREB2A), and its induced expression under heat stress contributes to thermotolerance, while suppressed expression increases heat sensitivity [91]. In tomato, Sly-NF-YA9/A10 function as negative regulators of heat tolerance; miR169-mediated downregulation of these NF-YAs relieves repression of heat stress-responsive genes, with heat shock factors (HSFs) transcriptionally regulating MIR169 to form a regulatory loop [20]. These findings highlight NF-Ys’ diverse roles in mediating heat stress responses through protein interactions and miRNA regulation. For a comprehensive summary of NF-Y transcription factors and their specific functions in horticultural plants under heat stress, refer to Table 6.
4.3.2. Cold Stress
Cold stress poses a severe threat to horticultural plants, and NF-Y transcription factors mediate cold tolerance through various regulatory mechanisms. In strawberry (Fragaria ananassa), CBF/NF-Y (YZ9)-overexpressed fruits promote coloring under cold treatment, while HSF20 (YZ1)-overexpressed ones are sensitive to cold/heat, regulating fruit quality traits [73]. In melon (Cucumis melo), 25 CmNF-Ys (6 NF-YAs, 11 NF-YBs, 8 NF-YCs) are identified, with 12 induced by cold stress, highlighting their key role in cold tolerance [36]. In pepper, CaNF-YC1 interacts with the TIFY family member protein CaTIFY7 to form a module, activating Capsicum annuum C-REPEAT BINDING FACTOR 1a/b (CaCBF1a/b) expression and enhancing cold tolerance by regulating antioxidant enzymes and stress-responsive genes [29]. These findings reveal NF-Ys’ multifaceted roles in cold signaling and tolerance. For a comprehensive summary of NF-Y transcription factors involved in cold stress responses in horticultural plants, refer to Table 7.
NF-Ys demonstrate remarkably diverse, even opposing, functions in temperature stress responses. In heat stress, they can be positive effectors (e.g., cucumber CsNFYA1 [90]) or negative regulators released by miR169 (e.g., tomato Sly-NF-YA9/A10 [20]). In cold stress, they often act as positive regulators within CBF-dependent or -independent pathways. This complexity likely arises from their position as hubs that integrate temperature signals with other environmental and developmental cues. A key insight is that their role cannot be generalized as simply “positive” or “negative”; instead, it is conditional, depending on the specific subunit, interacting partners (e.g., DREB2A, MBF1c), and the physiological process being modulated (e.g., photosynthesis protection vs. growth arrest).
4.4. Other Abiotic and Biotic Stresses
NF-Y transcription factors are also implicated in a range of additional abiotic and biotic stresses in horticultural plants. In tomato, under oxidative stress, the CCAAT-binding factor SlNFYA10 negatively regulates ascorbate biosynthesis by targeting the GDP- mannose-3′,5′-epimerase gene SlGME1 and the GDP-L-galactose phosphorylase gene SlGGP1 in the D-mannose/L-galactose pathway, thereby increasing sensitivity to oxidative stress [92]. In pigeon pea (Cajanus cajan), under aluminum (Al) stress, CcNFYB3 enhances citrate efflux by activating Cajanus cajan MULTIDRUG AND TOXIC COMPOUND EXTRUSION TRANSPORTER 35 (CcMATE35), while the Cajanus cajan long noncoding RNA targeting citrate synthase (CcLTCS) upregulates the Cajanus cajan citrate synthase gene CcCS to promote citrate synthesis; these two modules synergistically improve aluminum tolerance [93]. In watermelon, against stresses from fungal (Fusarium oxysporum) and bacterial (Pseudomonas syringae) pathogens, multiple ClNF-Y genes (e.g., ClNF-YB8, ClNF-YA3) mediate resistance via hormone-related signaling [31]. In common bean (Phaseolus vulgaris), for stress caused by the powdery mildew fungus Erysiphe diffusa, NF-YA3 is identified as a candidate gene associated with oligogenic resistance [94]. These findings broaden the functional scope of NF-Ys in both abiotic and biotic stress adaptation. For a comprehensive summary of NF-Y transcription factors involved in additional stress responses in horticultural plants, refer to Table 8.
NF-Ys are recurrently implicated in biotic stress responses, primarily through association with SA- and JA-mediated defense gene activation. However, a critical gap exists between correlation and mechanistic understanding. For most reported NF-Ys, it is unclear whether they directly bind defense gene promoters or exert effects indirectly by modulating hormone signaling components. The observation of both positive (e.g., CINF-YB8) and negative (e.g., CINF-YA2) regulators within the same species suggests a nuanced, subunit-specific regulatory network that is far from decoded [31]. Moreover, the current literature is strikingly focused on fungal and bacterial pathogens, leaving the role of NF-Ys in antiviral defense or resistance to insect herbivory—critical for horticulture—virtually unexplored.
5. Conclusions and Future Directions
This review systematically organizes the research progress of NF-Y transcription factors in horticultural plants, with a core focus on their molecular mechanisms in regulating plant developmental processes (e.g., phase transition, flowering, organogenesis) and responding to abiotic/biotic stresses. These core regulatory functions (encompassing representative developmental events and stress types in horticultural species) and the critical future research priorities aimed at addressing existing knowledge gaps are visually synthesized in Figure 2.
As conserved heterotrimeric complexes, NF-Ys integrate endogenous developmental signals and exogenous environmental cues to modulate the expression of target genes, acting as key regulators for optimizing horticultural plant growth, yield quality, and stress adaptability. While existing studies have clarified the functional roles of NF-Ys in multiple species and biological processes, there remain unaddressed knowledge gaps—such as incomplete regulatory networks and limited translational application—that hinder a comprehensive understanding of their potential, necessitating further targeted research to provide more solid theoretical support for horticultural crop genetic improvement.
Based on the above research status, future studies should first prioritize deciphering the crosstalk between NF-Ys and other regulatory layers in governing development and stress responses. Current research has preliminarily indicated interactions between NF-Ys and miRNAs, epigenetic modifications, or other transcription factors, but the global regulatory networks—including how these components synergize to fine-tune NF-Y function across different developmental stages and stress types—remain unclear. Employing integrative multi-omics approaches (e.g., combining transcriptomics with epigenomics or protein-protein interactomics) to systematically map these networks will help clarify the upstream regulators and downstream effectors of NF-Ys, deepening the understanding of their precise regulatory mechanisms.
Second, investigating NF-Y-mediated mechanisms under combined stress conditions is essential to bridge the gap between laboratory research and field applications. Most current studies focus on single stresses (e.g., drought, hot, cadmium stress), yet horticultural plants in natural environments frequently face concurrent stresses (e.g., drought and temperature stress) [96,97]. Future work should simulate field-relevant combined stress scenarios to explore how NF-Ys coordinate responses—such as whether they mediate cross-tolerance via shared signaling pathways (e.g., ABA signaling) or activate stress-specific cascades. This research will provide direct theoretical support for breeding climate-resilient horticultural cultivars capable of withstanding complex environmental challenges.
Third, efforts should be made to accelerate the functional validation of uncharacterized NF-Y genes, especially in understudied specialty horticultural species—such as ornamental flowers (e.g., orchid) and medicinal herbs (e.g., ginseng)—for which only genome-wide gene identification has been reported to date with functional characterization still lagging behind, given that many NF-Y genes across horticultural taxa have been identified via genome-wide analysis but lack in-depth functional characterization, and emerging regulatory modes of NF-Ys (e.g., involvement in secondary metabolism or beneficial plant-microbe interactions) also warrant exploration. Applying high-throughput functional screening techniques (e.g., virus-induced gene silencing, CRISPR/Cas9-based gene editing) to these uncharacterized candidates will not only expand the functional repertoire of NF-Y transcription factors but also enrich genetic resources for improving species-specific traits (e.g., ornamental value, medicinal component content) of specialty horticultural plants.
Fourth, translating NF-Y-related basic research into practical breeding tools needs to be strengthened. While some NF-Y genes have been validated as promising targets for trait improvement (e.g., enhancing stress tolerance, optimizing flowering time), challenges like off-target effects or pleiotropy (e.g., improved stress resistance trading off with yield) limit their application. Future translational research should focus on precision genetic modification—such as using tissue-specific or stress-inducible promoters to control NF-Y expression—and gene stacking to improve multiple traits simultaneously. Combining laboratory validation with field trials will ensure the stability and practicality of NF-Y-modified cultivars, fully exploiting the potential of NF-Ys for addressing key challenges in horticultural production.
Finally, a critical assessment of the translational challenges is essential to bridge NF-Y research and practical breeding. While the agronomic potential of NF-Ys is clear, several key bottlenecks must be addressed. Genetic redundancy within expanded NF-Y families, especially in polyploid or perennial crops, may buffer the effect of single-gene modifications, necessitating strategies like multiplex gene editing. Pleiotropic effects and trade-offs—where improved stress tolerance might compromise yield or development—require solutions such as the use of tissue-specific or stress-inducible promoters for precise spatiotemporal control. Ultimately, application must be evidence-based and context-specific, moving beyond proof-of-concept in model systems to demonstrate robust, field-validated performance in target horticultural species without adverse agronomic penalties.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhang H. Liu S.J. Ren T.M. Niu M.X. Liu X. Liu C. Wang H.L. Yin W.L. Xia X.L. Crucial abiotic stress regulatory network of NF-Y transcription factor in plants Int. J. Mol. Sci.202324442610.3390/ijms 2405442636901852 PMC 10002336 · doi ↗ · pubmed ↗
- 2Kishor P.B.K. Ganie S.A. Wani S.H. Guddimalli R. Karumanchi A.R. Edupuganti S. Naravula J. Kumar V. Polavarapu R. Suravajhala P. Nuclear factor-Y (NF-Y): Developmental and stress-responsive roles in the plant lineage J. Plant Growth Regul.2023422711273510.1007/s 00344-022-10739-6 · doi ↗
- 3Wu H. Hou X. Zhang C. Nuclear factor-Y transcription factors in crops: Biological roles, regulation, and breeding applications Plant Commun.2025610153010.1016/j.xplc.2025.10153040994001 PMC 12785170 · doi ↗ · pubmed ↗
- 4Siefers N. Dang K.K. Kumimoto R.W. Bynum W.E.IV Tayrose G. Holt B.F.III Tissue-specific expression patterns of Arabidopsis NF-Y transcription factors suggest potential for extensive combinatorial complexity Plant Physiol.200914962564110.1104/pp.108.13059119019982 PMC 2633833 · doi ↗ · pubmed ↗
- 5Hwang K. Susila H. Nasim Z. Jung J.Y. Ahn J.H. Arabidopsis ABF 3 and ABF 4 transcription factors act with the NF-YC complex to regulate SOC 1 expression and mediate drought-accelerated flowering Mol. Plant 20191248950510.1016/j.molp.2019.01.00230639313 · doi ↗ · pubmed ↗
- 6Zhao Y.J. Zhang Y.Y. Li T.J. Ni C.Y. Bai X.Y. Lin R.Z. Xiao K. Ta NF-YA 7-5B, a gene encoding nuclear factor Y (NF-Y) subunit A in Triticum aestivum, confers plant tolerance to PEG-inducing dehydration simulating drought through modulating osmotic stress-associated physiological processes Plant Physiol. Biochem.2022188819610.1016/j.plaphy.2022.07.03635988390 · doi ↗ · pubmed ↗
- 7Liu R. Wu M. Liu H.L. Gao Y.M. Chen J. Yan H.W. Xiang Y. Genome-wide identification and expression analysis of the NF-Y transcription factor family in Populus Physiol. Plant.202017130932710.1111/ppl.1308432134494 · doi ↗ · pubmed ↗
- 8Zheng X.R. Zhang H. Zhang L.M. Xu F.S. Shi L. Wang S.L. Hong J. Ding G.D. Identification and comprehensive analysis of the Nuclear Factor-Y family genes reveal their multiple roles in response to nutrient deficiencies in Brassica napus Int. J. Mol. Sci.2021221035410.3390/ijms 22191035434638695 PMC 8508618 · doi ↗ · pubmed ↗