Anti-Inflammatory Effects of Polyphenols in Brain Microvascular Endothelial Cells Stimulated with TNF-α
Joanna Czpakowska, Andrzej Glabinski, Piotr Szpakowski

TL;DR
This study shows that certain polyphenols reduce inflammation in brain endothelial cells, which could help protect the brain from inflammation-related damage.
Contribution
The study identifies specific polyphenols that effectively reduce inflammation and increase cell viability in brain endothelial cells.
Findings
Myricetin and chrysin reduced pro-inflammatory cytokine secretion in brain endothelial cells.
Chrysin and resveratrol downregulated VCAM-1 expression on brain endothelial cells.
Polyphenols increased cell viability and acted as antioxidants in brain endothelial cells.
Abstract
The blood–brain barrier (BBB) is a structure that regulates the exchange of substances between the peripheral circulation and the central nervous system (CNS), thereby protecting this environment. An increase in BBB permeability may lead to the influx of inflammatory cells, resulting in neuroinflammation and neurodegeneration. The integrity of the BBB is maintained due to the specific properties of brain endothelial cells. Considering the importance of brain endothelial cells in the BBB during inflammatory processes, these cells may be a target for anti-inflammatory agents. Polyphenols are substances exhibiting the ability to decrease inflammation; therefore, in our research, we aimed to examine their effectiveness in a brain endothelial cell culture stimulated with the pro-inflammatory cytokine TNF-α. The tested polyphenols were myricetin, chrysin, resveratrol, and curcumin. ELISA…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
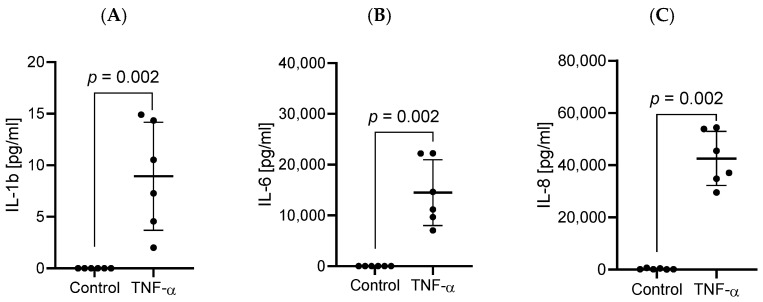 Figure 1
Figure 1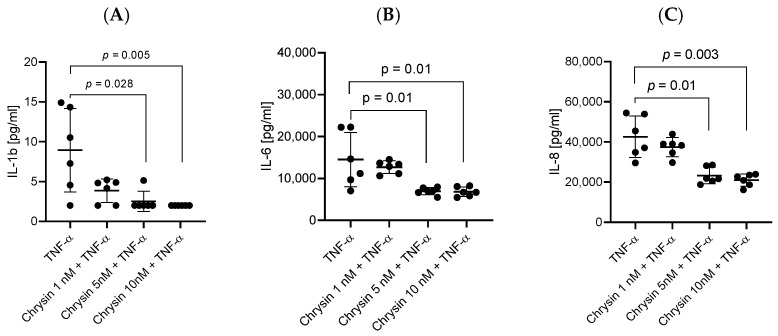 Figure 2
Figure 2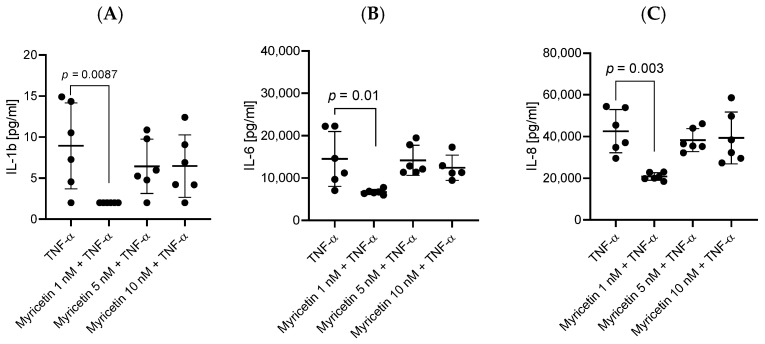 Figure 3
Figure 3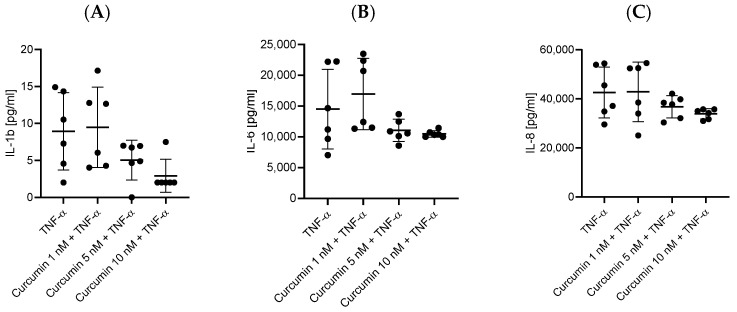 Figure 4
Figure 4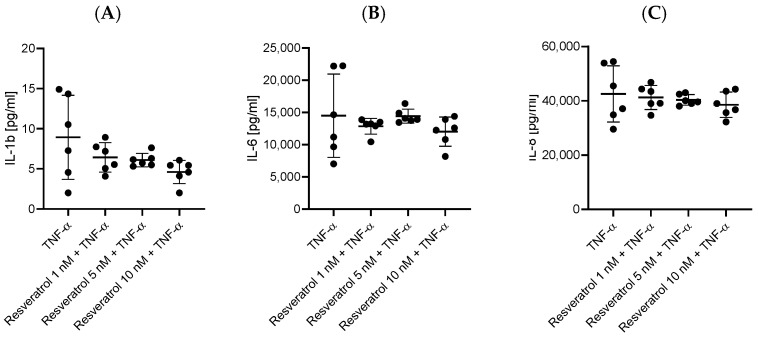 Figure 5
Figure 5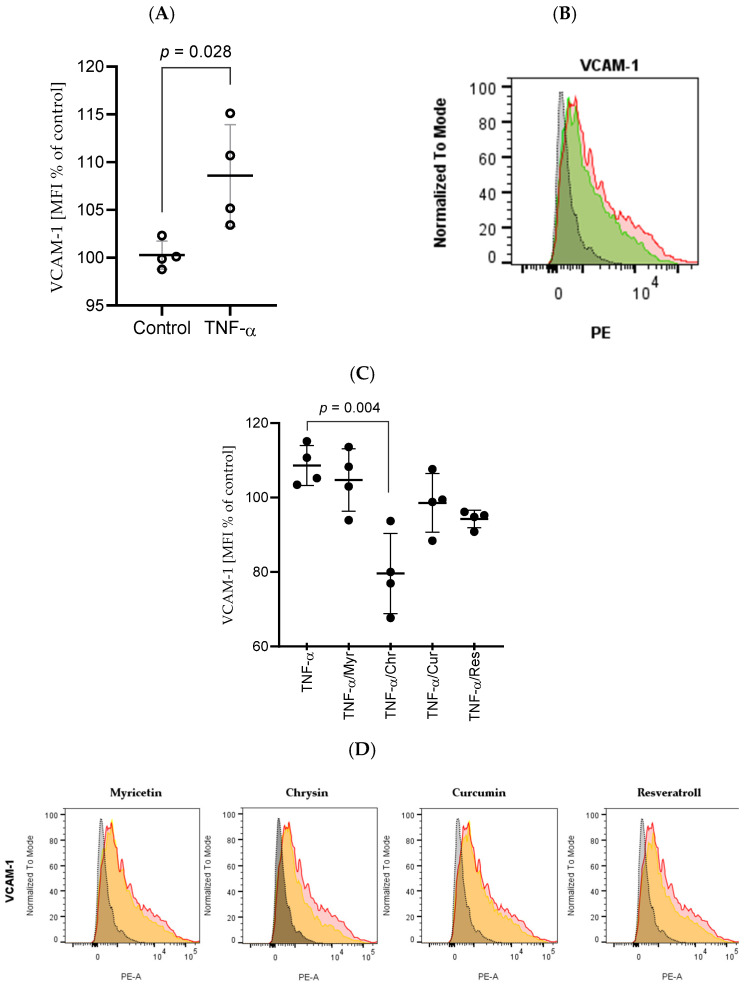 Figure 6
Figure 6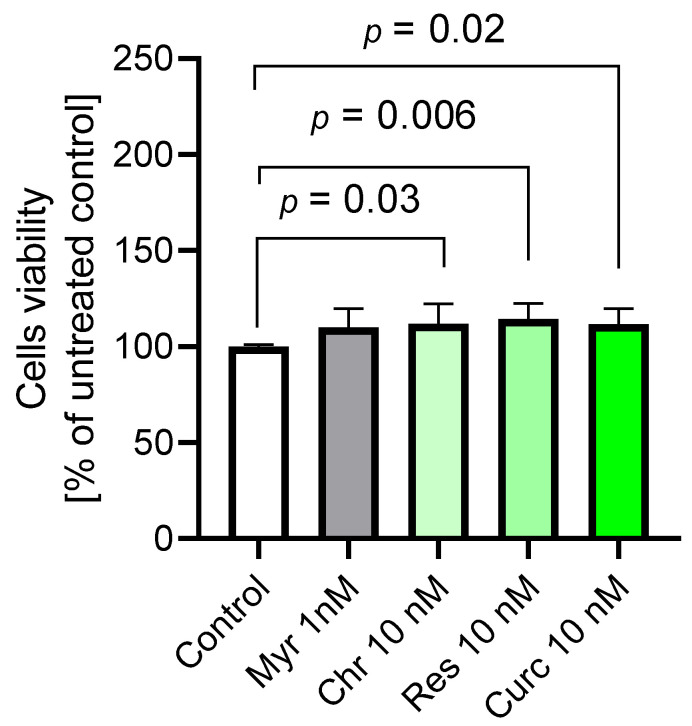 Figure 7
Figure 7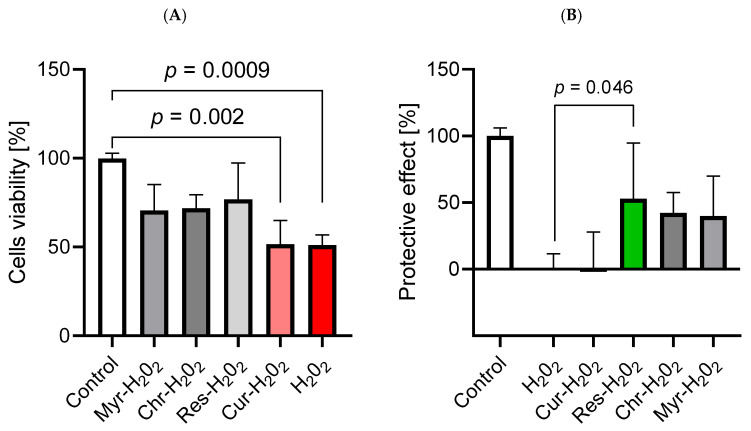 Figure 8
Figure 8- —Medical University of Lodz, Poland
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBarrier Structure and Function Studies · Flavonoids in Medical Research · Sirtuins and Resveratrol in Medicine
1. Introduction
Brain endothelial cells constitute the main component of the BBB, which plays a crucial role in the communication between the CNS and the circulatory system [1]. The proper functioning of brain endothelial cells, along with perivascular cells, astrocytes, microglia, and neuronal endings, which form the neurovascular unit (NVU), is essential in maintaining homeostasis in the CNS [2,3,4]. Brain endothelial cells contribute to the low permeability of the BBB due to their specific properties. The presence of tight junction (TJ) complexes on brain endothelial cells, formed by occludin, claudin-5, and junctional adhesion molecules, supported by VE-cadherin and E-cadherin, limits paracellular transport [5,6]. Transcellular passage is also restricted in brain endothelial cells [6,7]. Taken together, brain endothelial cells contribute to the activity of the BBB, which is able to prevent inflammation, inhibit toxins from entering the CNS, maintain ion balance, and correct signaling.
The improper functioning of the BBB can contribute to the emergence and progression of neurodegenerative and neuroinflammatory processes, leading to neurological dysfunctions such as multiple sclerosis (MS), Parkinson’s disease (PD), traumatic brain injury (TBI), and stroke, which highlights the importance of brain endothelial cells [2,8]. In the case of MS, the loss of BBB tightness, which is mainly based on TJs of brain endothelial cells, leads to the influx of immune cells. This process triggers inflammation and leakage of plasma proteins, such as fibrinogen, which results in their accumulation in demyelinating lesions [9,10,11]. Similar processes are observed in PD, where the entrance of α-synuclein precedes the secretion of inflammatory agents by NVU cells through the impaired BBB [12]. TBI can also be the cause of BBB damage, which can spread more intensively due to inflammatory responses after activation of microglia and astrocytes by leaking albumin [13]. The disruption of the BBB that takes place after a stroke is driven by oxidative stress. At the initial stage, reactive oxygen species (ROS) affect TJs and the cytoskeleton of brain endothelial cells. Next, metalloproteinases (MMPs) degrade TJs. As a consequence of BBB damage, cerebral edema emerges. Brain endothelial cells take part in ionic edema directly because their ion channels allow increased passage of Na^+^, Cl^−^, and water. To summarize, BBB integrity is based on the proper functioning of brain endothelial cells, especially of their TJs. Neuroinflammatory and neurodegenerative processes contribute to BBB dysfunction; therefore, brain endothelial cells constitute a promising target for future therapeutic approaches [10,14].
One of the candidates demonstrating anti-inflammatory properties is polyphenols. The group of phenolic compounds is divided into various sub-groups such as flavonoids, stilbenes, phenolic acids, and lignans. The division is based on similarities in their primary structures [15,16]. Polyphenols are widely available in fruits, herbs, and spices, which means that the human diet provides many sources of them. The role of these compounds in plants is not fully understood and may be implicated in defense against ultraviolet radiation and hostile microbial agents. In addition to these functions, polyphenols act as antioxidants by inhibiting ROS formation. Polyphenols can suppress enzymes that catalyze ROS synthesis, eliminate them directly, or enhance defense mechanisms. The reduction of ROS by polyphenols contributes to the constriction of tissue injuries, which can otherwise lead to inflammation. Additionally, the involvement of polyphenols in the inhibition of inflammatory signaling pathways and in the activity of inflammatory cells and enzymes engaged, for instance, in arachidonic acid metabolism, proves their anti-inflammatory effect [17,18,19].
2. Results
2.1. Stimulation of HBEC-5i Cells with TNF-α Resulted in the Increased Secretion of Pro-Inflammatory Cytokines
In response to the stimulation of HBEC-5i cells with TNF-α, the secretion of the pro-inflammatory cytokines IL-1ß (p = 0.0022), IL-6 (p = 0.002), and IL-8 (p = 0.002) was intensified in comparison to the control group with medium alone where these cytokines were not detected (Figure 1).
2.2. Treatment of HBEC-5i Cells with Polyphenols Reduced the Concentration of Secreted Pro-Inflammatory Cytokines
HBEC-5i cells stimulated simultaneously with TNF-α and one of the tested polyphenols (chrysin, myricetin, curcumin, and resveratrol), at 1 nM for myricetin and 5 or 10 nM for the others, showed a decrease in the levels of secreted pro-inflammatory cytokines compared to stimulation with only TNF-α. The most effective concentrations lowering the levels of IL-1ß, IL-6, and IL-8 turned out to be 10 nM for chrysin, curcumin, and resveratrol and 1 nM for myricetin. Statistically significant results in downregulating the expression of IL-1ß, IL-6, and IL-8 are assigned to chrysin (p = 0.005, p = 0.01, and p = 0.0033, respectively) (Figure 2) and myricetin (p = 0.0087, p = 0.01, p = 0.0033) (Figure 3). Notably, curcumin (Figure 4) and resveratrol (Figure 5) also reduced concentrations of secreted cytokines; however, their effects were not statistically significant.
2.3. Exposure of HBEC-5i Cells to TNF-α Led to the Upregulation of CD106 (VCAM-1) Expression
Flow cytometry was used to measure the expression of CD106 on the surface of HBEC-5i cells. Stimulation of HBEC-5i cells with TNF-α resulted in an increased level of CD106 in comparison to unstimulated cells (p = 0.028) (Figure 6).
2.4. Chrysin, Myricetin, Curcumin, and Resveratrol Caused Downregulation of CD106 Expression
Analysis of flow cytometry results from HBECs stimulated with TNF-α and polyphenols showed weaker CD106 upregulation in response to TNF-α in the presence of chrysin (p = 0.028) and resveratrol (p = 0.028). Stimulation of cells with curcumin and myricetin did not result in a statistically significant downregulation of CD106.
2.5. The Application of Polyphenols Confirmed Their Antioxidative Properties and Resulted in the Improvement of Cerebral Microvascular Endothelial Cell Viability
The obtained results indicated that chrysin, resveratrol, and curcumin were involved in improving HBEC-5i cell viability (p = 0.03, p = 0.006, p = 0.02, respectively), which may be attributed to an increased mitochondrial metabolic rate or enhanced cellular proliferation. Myricetin did not show this effect. Tested compounds at the indicated concentrations did not reduce cell viability, but increased it, compared to the untreated control (medium), indicating a lack of intrinsic cytotoxicity (Figure 7). Simultaneously, these polyphenols acted as oxidants in groups of cells treated with H_2_O_2_. The viability of cells treated with H_2_O_2_ was significantly lower than that in the group with medium alone (p = 0.0009). Simultaneous treatment of cells with polyphenol and H_2_O_2_ resulted in increased viability. Only curcumin did not show the antioxidant effect (Figure 8A). The protective effect of polyphenols was the most visible in the case of resveratrol, which most effectively improved viability compared to the H_2_O_2_ group (p = 0.046) (Figure 8B).
3. Discussion
In the context of polyphenols’ application in the prevention and treatment of neurodegenerative and neuroinflammatory diseases, it is essential to analyze the molecular mechanisms underlying their anti-inflammatory, immunomodulatory, and antioxidant activities. Polyphenols modulate the immune system in many diverse ways. They can influence the production of proinflammatory cytokines and the activity of immune cells, and they can inhibit the expression of proinflammatory genes. Additionally, polyphenols affect many signaling pathways associated with immunity, such as nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and mitogen-activated protein kinase (MAPK). Polyphenols also contribute to ROS elimination by suppressing enzymes involved in their synthesis, such as NADPH oxidase (NOX), and by increasing the production of antioxidant enzymes, for instance, superoxide dismutase (SOD). This antioxidant activity reduces inflammation [20,21].
There are a few promising polyphenols worth attention for their anti-inflammatory activity. One of them is chrysin, a flavone from the flavonoid family [22]. Flavonoids constitute the most abundant group of polyphenols in our diet, with an average intake of 50–800 mg/day. In addition to their anti-inflammatory activity, they have antioxidant, neuroprotective, antiviral, cardioprotective, and anticarcinogenic properties [22,23]. Similar to other polyphenols, chrysin exhibits low bioavailability (in the case of oral consumption: 0.003–0.02%) and poor solubility. Additionally, it is metabolized rapidly and quickly eliminated. Therefore, to fully exploit the potential of chrysin and other polyphenols, strategies that overcome these restrictions must be developed [22,23,24].
The anti-inflammatory activity of chrysin is based on its modulation of key immunomodulatory mechanisms. Chrysin can decrease the levels of pro-inflammatory enzymes, including myeloperoxidase (MPO), cyclooxygenase-2 (COX-2), and inducible nitric oxide synthase (iNOS), by activating the peroxisome proliferator-activated receptor gamma (PPAR-γ). Inhibiting the NF-κB signaling pathway is another way of lowering the expression of iNOS and COX-2 by chrysin [25]. Additionally, suppressed by chrysin, NF-κB is not able to induce the production of pro-inflammatory cytokines such as IL-1β, IL-6, and IL-8 [26]. MAPK is another signaling pathway that can be influenced by chrysin. Among all kinases belonging to the MAPK family, which are extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38, this polyphenol selectively inhibits JNK by reducing its phosphorylation. As a result, the secretion of proinflammatory cytokines is decreased [27].
Another noteworthy member of the flavonoid family and the flavonol sub-group is myricetin. This polyphenol exhibits properties similar to those of other flavonoids, including anti-inflammatory and antioxidant activities. The distinguishing feature of myricetin is its high effectiveness at low doses in the elimination of free radicals [28,29]. The increased amount of hydroxyl groups in comparison to other flavonoids (three groups on ring B) in the myricetin chemical structure is thought to be responsible for its antioxidant properties [28,30]. Contrarily, some studies indicated that myricetin can also act in a pro-oxidative way by reducing oxygen to ROS and iron (III) to iron (II) [31].
Myricetin, similarly to chrysin, can inhibit the NF-κB pathway and downregulate iNOS and COX-2 [32,33]. In terms of the MAPK pathway, myricetin does not act selectively but inhibits all of its kinases (ERK, JNK, and p38) [32]. Myricetin, by inhibiting toll-like receptor 2 (TLR2), suppresses the activity of the mTOR signaling pathway, which contributes to lowered production of pro-inflammatory cytokines such as IL-6 and IL-1β [34]. In the context of CNS inflammatory processes, myricetin can help downregulate post-ischemic neuroinflammation. This polyphenol acts as an inhibitor of the nucleotide-binding oligomerization domain-like receptor protein 3 (NLRP3). After its activation, this receptor forms an inflammasome that triggers caspase-1, leading to the maturation of pro-inflammatory cytokines such as IL-1β. Due to myricetin activity, these processes are suppressed, and therefore, inflammation can be halted [35,36].
Besides flavonoids, polyphenols include other families, such as stilbenes. The best-known stilbenoid is resveratrol. This compound can be found mainly in grapes [37,38]. Similar to other mentioned polyphenols, resveratrol acts as an anti-inflammatory, antioxidative, anticarcinogenic, cardioprotective, and immunomodulatory agent. The presence of hydroxyl groups and a double-bonded ethylene bridge is the cause of resveratrol’s instability and sensitivity to pH, light, and increased temperature [39,40]. Additionally, resveratrol is quickly metabolized and has poor solubility and a low rate of bioavailability (<10 ng/mL after 25 mg intake) [41,42].
Resveratrol is another polyphenol reported to have inhibitory effects on the NF-κB signaling pathway. The mechanism of resveratrol’s action on this pathway is based on influencing different targets. One of them may be the activation of sirtuin-1 (Sirt-1), which is a deacetylase that suppresses the TLR-4/NF-κB/STAT pathway. Additionally, resveratrol can prevent NF-κB activation mediated by the inhibitor of κBβ (IκBβ) and by IL-1β. In the case of the MAPK family, resveratrol can inhibit all of its kinases [43]. Lowering the expression of COX-2 and iNOS by resveratrol is another common pathway for polyphenols for decreasing inflammation [44]. Resveratrol can also downregulate inflammation by the inhibition of activator protein-1 (AP-1), which regulates gene expression related to inflammatory processes [43]. Resveratrol was also proven to lower the amount of pro-inflammatory cytokines such as TNF-α, IFN-γ, IL-2, IL-9, IL-12, and IL-17 [45].
Curcuminoids serve as another example of the polyphenol family. The most prevalent member of this group is curcumin, exhibiting anti-inflammatory, immunomodulatory, and antioxidant properties [46]. Curcumin has low bioavailability and absorbs poorly after oral intake; therefore, despite its broad range of potential applications, it is difficult to exploit its full potential. One of the ways of increasing curcumin’s bioavailability by 2000% is simultaneous consumption of piperine [47,48].
Curcumin’s anti-inflammatory effect can be seen, for instance, through its ability to lower monocyte chemoattractant/chemotactic protein-1 (MCP-1) expression. This chemokine intensifies the inflammatory response, and various studies have indicated that curcumin inhibits the synthesis of MCP-1. Curcumin can exert this effect, similarly to other polyphenols, by influencing several signaling pathways, mainly MAPK (especially JNK) and NF-κB but also protein kinase C (PKC) and MMP, which are considered targets [49]. Curcumin was also reported to exhibit anti-inflammatory properties by influencing the Janus kinase/signal transducer and activator of transcription (JAK/STAT) signaling pathway [50]. The inhibition of NF-κB by curcumin was additionally proven to lower the activity of COX-2, iNOS, and lipoxygenase (LOX) [49]. Another consequence of curcumin acting on NF-κB is suppressing the activation of the NLRP3 inflammasome [50,51].
To assess the effectiveness of the mentioned polyphenols in reducing inflammation, in our study, HBEC-5i cells were stimulated with TNF-α. This pro-inflammatory agent, under in vivo conditions, acts specifically on the brain endothelial cells by activating the TLR4/NFκB signaling pathway and phosphatidylinositol-3 kinase (PI3K), which results in lowered expression of TJs proteins (ZO-1, claudin-5, and occludin). As a consequence, the BBB is disrupted, which may be the initial step of neuroinflammation and neurodegeneration [5,10]. TNF-α also contributes to the disruption of the BBB by activating Rho-kinase and stimulating the production of substance P (SP) with its receptor neurokinin 1 (NK1R). RhoA protein, after binding with the Rho-kinase, causes phosphorylation of the myosin light chain (MLC), resulting in the disruption of TJs. Additionally, the level of nitric oxide (NO) acting as an antioxidant is lowered. SP, by acting on NK1R, induces the activation of the majority of cytokines and therefore takes part in the inflammatory processes [25].
TNF-α activates many signaling pathways in the brain endothelial cells, resulting in an increase in inflammation. This mechanism of action leads to the secretion of pro-inflammatory cytokines such as IL-6, IL-8, and IL-1β, which may serve as indicators of the ongoing inflammation.
One of the primary roles of IL-6 is to cause the differentiation and activation of B cells, which allows them to synthesize antibodies. IL-6 also fulfils the role of activating T lymphocytes and causes an increase in their number [52]. This cytokine appears rapidly after infection or injury. The intensified production of IL-6 leads to chronic inflammation, which is the cause of many pathological conditions [53,54]. The secretion of IL-6 after stimulation of the brain endothelial cells with TNF-α was observed in models of neuroinflammation, including the study conducted by Rochfort et al. and our research [55]. The elevated IL-6 level in HBEC-5i cells cultured in medium alone, compared with the control group, indicates the correct induction of the inflammatory response. The polyphenols assessed for anti-inflammatory properties demonstrated effectiveness. The application of chrysin and myricetin was confirmed, with statistical significance, to attenuate inflammation by lowering IL-6 levels. Our findings regarding chrysin align with the results of a study by Ma et al. performed on human coronary artery endothelial cells. The cells stimulated with TNF-α secreted an increased level of IL-6, which was downregulated by chrysin. The proposed chrysin’s mechanism of action relies on modulation of nuclear factor of activated T cells 2 (NFAT2) [56]. Similarly, the study on human umbilical vein endothelial cells (HUVECs), induced with lipopolysaccharide (LPS), conducted by Lin et al., revealed that chrysin decreases the IL-6 concentration [57]. In the case of myricetin, its effectiveness was also confirmed by Chen et al. in a study on A549 cells. The upregulated IL-6 level after TNF-α stimulation was reduced by myricetin. Myricetin, by activating Sirt-1, leads to the reduction of NF-κB p65 and p53 acetylation, which results in the inhibition of IL-6 secretion [58]. In the case of resveratrol and curcumin, their anti-inflammatory effect was visible, but it cannot be confirmed statistically that these polyphenols are effective agents against inflammation in the HBEC-5i cell environment. Despite our findings, a study conducted by Kolahdouz-Mohammadi et al. reports that resveratrol effectively downregulates IL-6 expression in endometrial stromal cells from patients with endometriosis [59]. Regarding curcumin’s ability to reduce IL-6 concentration, it was confirmed by Cho et al. in a study on HaCaT cells induced with TNF-α. Curcumin inhibited the activation of MAPKs and NF-κB, leading to the downregulation of IL-6 expression [60].
IL-8 is another tested cytokine, secreted in response to tissue injury and infection. It exhibits pro-inflammatory properties by recruiting lymphocytes, especially neutrophils, to the target area [61,62]. Additionally, IL-8 activates neutrophils, allowing them to discharge their granule contents [63]. In our research, it was observed that the stimulation of HBEC-5i cells with TNF-α induced strong secretion of IL-8 in comparison to the control group. This response of brain endothelial cells to TNF-α was also confirmed in the study conducted by O’Carroll et al. [64]. The anti-inflammatory activity of polyphenols was similar to that previously described. Chrysin and myricetin significantly decreased the concentration of the pro-inflammatory cytokine IL-8, whereas resveratrol and curcumin showed only a tendency to lower inflammation, without statistical confirmation. The activity of chrysin in reducing the expression of IL-8 was also confirmed in a study by Lee et al. conducted on HEK 293 cells stimulated with TNF-α. The results indicated that chrysin suppresses IL-8 promoter activation and gene expression. Additionally, chrysin prevents the translocation of NF-κB p65 [65]. Regarding myricetin, Chen et al. obtained similar results in a previously mentioned study on A549 cells. The elevated IL-8 level induced by TNF-α was reduced by myricetin [58]. Contrary to our findings regarding resveratrol, Oh et al. demonstrated that it can decrease IL-8 production in LPS-induced THP-1 cells. It was concluded that resveratrol achieves this effect by inhibiting MAPK phosphorylation and NF-κB activation [66]. In the context of curcumin’s effectiveness, Wang et al. demonstrated in their study on HUVECs that this polyphenol decreases the expression of IL-8 by modulating the ERK and p38 MAPK signaling pathways [67].
We demonstrated in our study that stimulation of HBEC-5i cells with TNF-α results in the secretion of another pro-inflammatory cytokine, which is IL-1β. This cytokine initially exists in an inactive form, pro-IL-1β. In its activation to the mature form, the NLRP3 inflammasome is involved. IL-1β is released by activated macrophages under inflammatory conditions and is engaged in the attraction of leukocytes to the affected site [68]. Additionally, it is involved in the activation and differentiation of T lymphocytes, which produce IL-17 cytokine [69]. Similar to the previous results, chrysin and myricetin effectively suppressed the production of IL-1β cytokine, proving their anti-inflammatory properties statistically. Similar results regarding chrysin were reported by Ha et al. in a study on microglia activated with LPS. Chrysin was shown to decrease IL-1β secretion. Additionally, it was confirmed that this polyphenol suppresses the activation of JNK and NF-κB [27]. The ability of myricetin to lower the expression of IL-1β was confirmed in a study conducted by Park et al. on RAW 264.7 murine macrophages. The cells treated with endocrine-disrupting chemical Di(2-ethylhexyl) phthalate (DEHP) showed an increased level of IL-1β, which was reduced by myricetin, acting as an inhibitor of PKC and ERK [70]. Resveratrol and curcumin use were associated with the reduction of IL-1β; however, these changes were not statistically significant. Despite our findings concerning resveratrol, the study by Tian et al. on human rheumatoid arthritis fibroblast-like synoviocytes induced with TNF-α revealed that it can reduce the amount of IL-1β by inhibiting the PI3K-Akt signaling pathway [71]. Regarding curcumin, the study confirming its ability to decrease IL-1β levels was conducted by Yin et al. in mouse bone marrow–derived macrophages induced with LPS. The proposed curcumin’s mechanism of action is based on inhibition of NLRP3 inflammasome activation [72].
The effectiveness of tested polyphenols in reducing the amount of the pro-inflammatory cytokines IL-6, IL-8, and IL-1β can be attributed to their ability to inhibit several signaling pathways responsible for the increase of inflammation. Chrysin, myricetin, resveratrol, and curcumin have been shown to suppress the activity of the NF-κB and MAPK pathways, which are the primary inflammatory regulators responsible for the secretion of the mentioned cytokines. The limited effectiveness of resveratrol and curcumin in the inhibition of cytokine production by HBEC-5i cells may be attributed to their especially low stability. These data lay the foundations for further research focusing on the enhancement of their durability and delivery methods. Chrysin and myricetin proved their efficacy; however, myricetin emerged as the optimal candidate for downregulating inflammation in the HBEC-5i cell environment since it is most effective at the lowest concentration.
We also focused in our research on measuring the expression of vascular cell adhesion molecule-1 (VCAM-1) present on the surface of brain endothelial cells after stimulation with TNF-α. VCAM-1 belongs to the group of cell adhesion molecules, present mainly on endothelial cells, which under inflammatory conditions are responsible for adhesion of leukocytes to the cell surface and allow for their transendothelial migration. The process of leukocyte adhesion to VCAM-1 is possible due to the presence of α4β1 integrin on the leukocyte surface, which directly binds to VCAM-1, leading to the activation of signaling pathways and resulting in TJs deterioration and remodeling of actin [73,74]. Similarly to the results obtained in the study conducted by O’Carroll et al., we observed intensified expression of VCAM-1 after the treatment of HBEC-5i cells with TNF-α [64]. Also, the research conducted by Wojkowska and Szpakowski on the murine brain endothelial cells revealed that their induction with TNF-α results in the increase of VCAM-1 presence [74]. The expression of VCAM-1 after stimulation with TNF-α is possible because this cytokine induces signaling pathways in which NF-κB and activator protein 1 (AP1), belonging to the MAPK cascade, are activated and stimulate the expression of the VCAM-1 gene [73,75,76]. The increased level of VCAM-1 favors inflammation; therefore, it is essential to identify the anti-inflammatory agents able to lower its expression. Our research shows that the tested polyphenols turned out to be the optimal candidates. Chrysin and resveratrol decreased the amount of VCAM-1 on the surface of HBEC-5i cells compared with the TNF-α-only group, validating their effectiveness in eradicating inflammation. Our results regarding chrysin’s effectiveness in attenuating VCAM-1 expression in brain endothelial cells are consistent with the study by Lee et al. on murine brain endothelial cells. This study demonstrated that chrysin reduces the expression of VCAM-1 by suppressing NF-κB translocation and the MAPK pathway [77]. The study conducted by Zhao et al. also confirms chrysin’s mechanism of action [78]. The findings from our study focusing on resveratrol’s ability to downregulate VCAM-1 expression align with the study by Zhang et al., where endothelial progenitor cells were treated with TNF-α and resveratrol. Similarly to our results, TNF-α upregulated the expression of VCAM-1, while application of resveratrol led to its reduction by NF-κB inhibition [79]. Myricetin and curcumin also reduced the expression of VCAM-1; however, their activity was not statistically significant when compared with more effective resveratrol and chrysin. However, in the literature, studies can be found confirming the effectiveness of these polyphenols on different types of cells. Bai et al. report that myricetin downregulates VCAM-1 expression in HUVECs induced with oxidized low-density lipoprotein [80]. In the case of curcumin, Kim et al. confirm in the study on human endometriotic stromal cells induced with TNF-α that this polyphenol reduces the expression of VCAM-1 [81]. Because of the involvement of NF-κB and MAPK in VCAM-1 overexpression and the proven ability of the tested polyphenols to inhibit activities of these signaling pathways, it can be concluded that these pathways constitute the primary targets of chrysin, myricetin, curcumin, and resveratrol.
Polyphenols are known for their antioxidant properties, which we also confirmed in our study on HBEC-5i cells. Their ability to eliminate ROS is mainly based on the abundant presence of phenolic hydroxyl groups acting as active hydrogen donors. ROS, after receiving the hydrogen atom, becomes a stable molecule. Polyphenols, after losing a hydrogen atom, are safe for cells and less toxic than ROS. Additionally, polyphenols increase the activity of antioxidant enzymes [82]. In our study, only curcumin lacked antioxidant activity. This observation can be explained by the fact that curcumin is highly unstable. Under conditions of 37 °C and pH 7.2, the t_1/2_ for curcumin is approximately 10 min. Additionally, curcumin can contribute to the generation of ROS [48]. This polyphenol can covalently modify Cys496 and Sec497 residues in thioredoxin reductase (TrxR), which leads to the loss of its reduction activity of thioredoxins (Trxs) and transformation into NADPH oxidase, which produces ROS [83]. The selected polyphenols in our study also showed potency in increasing the viability of HBEC-5i cells. The mechanism of action behind this phenomenon is based on the polyphenols’ influence on mitochondria. The tested polyphenols activate Sirt-1, leading to the deacetylation of PGC-1α. In consequence, the transcription of nuclear and mitochondrial genes takes place, resulting in the increased proliferation of mitochondria and therefore improved cell metabolism and survival [84,85,86,87].
4. Materials and Methods
4.1. Cerebral Microvascular Endothelial Cell Culture
The cerebral microvascular endothelial cell line (HBEC-5i) was isolated from human cerebral cortex and purchased from ATCC (Manassas, VA, USA; CRL-3245™). The cells were cultured in 75 cm^2^ culture flasks coated with 0.1% gelatin (Sigma–Aldrich, St. Louis, MO, USA, 1 mL per 10 cm^2^. Cells were grown in culture medium DMEM-F12 (Biowest, Nuaillé, France) supplemented with 1% of Antibiotic Antimycotic Solution (penicillin, streptomycin, amphotericin) (Sigma–Aldrich), 40 μg/mL endothelial growth supplement (ECGS) (Corning^®^, Corning, NY, USA), and 10% of heat-inactivated fetal bovine serum (FBS) (Biowest, Nuaillé, France) in 5% CO_2_ atmosphere, 37 °C, and increased humidity. The culture medium was renewed every 2–3 days. When the cell culture reached 90% confluency, the cells were washed with Dulbecco’s PBS (Biowest, Nuaillé, France) and then collected with the use of Trypsin-EDTA (ethylenediaminetetraacetic acid) solution (Sigma–Aldrich) and 10% FBS in PBS, centrifuged (125× g, 6 min, 20 °C), and seeded on 24-well culture plates coated with gelatin (70 × 10^3^/well), and cultured for 24 h, enabling cells to bind to the growth surface.
4.2. Stimulation of HBEC-5i Cells with TNF-α and Polyphenols
HBEC-5i cells were stimulated with 5 ng/mL rhTNF-α (R&D Systems, Minneapolis, MN, USA) to induce the pro-inflammatory response and also with polyphenols: chrysin, myricetin, curcumin, and resveratrol (all reagents from Sigma–Aldrich), at concentrations of 1 nM for myricetin and 10 nM for the rest, to confirm their anti-inflammatory properties and establish the most effective concentration. The control group constituted cells cultured in medium alone. The cells after the stimulation were incubated for 48 h in 24-well culture plates. After this time, the culture medium was collected and centrifuged (5000× g, 10 min, 20 °C). The obtained supernatants were placed at −80 °C. Simultaneously with the medium collection, the remaining cells were harvested after 48 h and prepared for flow cytometry analysis.
4.3. Analysis of HBEC-5i Cell Response with the ELISA Test
The concentrations of cytokines secreted by HBEC-5i cells were measured using ELISA kits (DuoSet, R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s protocols. The collected culture media were analyzed in terms of the presence of the pro-inflammatory cytokines IL-1ß, IL-6, and IL-8. The supernatants used for the measurement of IL-6 and IL-8 concentrations were diluted 50 times prior to the ELISA test. The concentration of IL-1ß was measured without dilution of culture media.
4.4. Assessment of CD106 (VCAM-1) Expression with Flow Cytometry
After 48 h of incubation, HBEC-5i cells were collected on ice using cell dissociation solution (Sigma–Aldrich, St. Louis, MO, USA). Next, the cells, including antibody isotype controls, were washed with cold 1% FBS in PBS and stained with anti-CD106 PE mouse IgG1 κ (Biolegend, San Diego, CA, USA). The cells were incubated with the fluorescently labelled antibodies for 30 min, 4 °C, in the dark, then washed twice with cold 1% FBS in PBS, fixed with formalin (20 min, on ice), again washed twice with cold 1% FBS in PBS, and frozen in FBS with 10% dimethyl sulfoxide (DMSO; Sigma–Aldrich, St. Louis, MO, USA). Flow cytometry was performed on an LSR II flow cytometer (Becton Dickinson, San Jose, CA, USA).
4.5. Measurement of HBEC-5i Cell Viability and Antioxidative Properties of Polyphenols
HBEC-5i cells were seeded into two 96-well plates at a density of 12.5 × 10^3^/well under 5% CO_2_, 37 °C conditions. After 24 h, the cells on both plates were stimulated with 1 nM mirycetin, 10 nM chrysin, 10 nM resveratrol, and 10 nM curcumin, excluding the control group. After 24 h of stimulation with polyphenols, cells from one of the plates were treated with 250 µL H_2_O_2_; the other plate was incubated without changes. Then, 48 h after stimulation with polyphenols, the viability and antioxidative properties of the polyphenols were measured using EZMTT assay (Merck Millipore, Darmstadt, Germany). After incubation, the medium was removed and replaced with 100 µL of fresh medium containing EZMTT reagent diluted 200 times per well. The plates were incubated for 3 h at 37 °C, and the absorbance was measured at λ = 450 nm.
4.6. Statistical Analysis
For comparisons between two groups, the non-parametric Mann–Whitney U test was conducted. Comparisons between multiple groups were performed using non-parametric ANOVA. The non-parametric analysis of the data was performed due to the low sample number. When Kruskal–Wallis tests were considered significant (p < 0.05), post-hoc Dunn’s tests were utilized for precise comparisons between groups. Statistical tests were performed using Statistica 13.00. Flow cytometry data analysis was performed using FlowJo 10 software.
5. Conclusions
Brain endothelial cells present themselves as a promising target for anti-inflammatory agents such as polyphenols. In our study, myricetin, chrysin, resveratrol, and curcumin were tested for decreasing the inflammatory response of HBEC-5i cells after stimulation with pro-inflammatory TNF-α. Myricetin and chrysin showed statistically confirmed effectiveness in lowering the concentration of secreted IL-6, IL-8, and IL-1β. Additionally, myricetin was the most effective at the lowest concentration. Resveratrol and curcumin presented a tendency in decreasing inflammation; however, these results were not statistically significant. Chrysin and resveratrol reduced the expression of VCAM-1, which is a molecule common for endothelial cells, responsible for intensifying inflammation. The selected polyphenols also increased the viability of HBEC-5i cells and showed antioxidant properties. The effects of polyphenols presented here suggest that they are optimal candidates for combating inflammation in the BBB environment. Despite promising results in the in vitro study, they might not be as effective in vivo. Due to their poor bioavailability, solubility, and high rate of metabolism, it is essential to develop strategies allowing for the delivery of polyphenols to the target, where their anti-inflammatory properties can be utilized.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang Y. Wang N. Cai B. Wang G. Li J. Piao X. In Vitro Model of the Blood-Brain Barrier Established by Co-Culture of Primary Cerebral Microvascular Endothelial and Astrocyte Cells Neural Regen. Res.201510201110.4103/1673-5374.17232026889191 PMC 4730827 · doi ↗ · pubmed ↗
- 2Li X. Simo L. Zhao Q. Kim E.G. Ding Y. Geng X. Endothelial Cells and the Blood–Brain Barrier: Critical Determinants of Ineffective Reperfusion in Stroke Eur. J. Neurosci.202561 e 1666310.1111/ejn.1666339935212 PMC 11814926 · doi ↗ · pubmed ↗
- 3Grammas P. Martinez J. Miller B. Cerebral Microvascular Endothelium and the Pathogenesis of Neurodegenerative Diseases Expert. Rev. Mol. Med.201113 e 1910.1017/S 146239941100191821676288 · doi ↗ · pubmed ↗
- 4Mc Connell H.L. Mishra A. Cells of the Blood–Brain Barrier: An Overview of the Neurovascular Unit in Health and Disease Methods Mol. Biol.202224923243573303610.1007/978-1-0716-2289-6_1PMC 9987262 · doi ↗ · pubmed ↗
- 5Xiang Y. Gu Q. Liu D. Brain Endothelial Cells in Blood–Brain Barrier Regulation and Neurological Therapy Int. J. Mol. Sci.202526584310.3390/ijms 2612584340565303 PMC 12193000 · doi ↗ · pubmed ↗
- 6Chow B.W. Gu C. The Molecular Constituents of the Blood–Brain Barrier Trends Neurosci.20153859860810.1016/j.tins.2015.08.00326442694 PMC 4597316 · doi ↗ · pubmed ↗
- 7Ashby J.W. Mack J.J. Endothelial Control of Cerebral Blood Flow Am. J. Pathol.20211911906191610.1016/j.ajpath.2021.02.02333713686 · doi ↗ · pubmed ↗
- 8Patabendige A. Janigro D. The Role of the Blood–Brain Barrier during Neurological Disease and Infection Biochem. Soc. Trans.20235161362610.1042/BST 2022083036929707 PMC 10212550 · doi ↗ · pubmed ↗