The Role of Probiotics Limosilactobacillus reuteri, Ligilactobacillus salivarius, and Lactobacillus johnsonii in Inhibziting Pathogens, Maintaining Gut Health, and Improving Disease Outcomes
Li Li, Xiangqi Qiu, Shengyong Lu, Haitao Yu, Panpan Lu, Sumei Zeng, Aihua Deng, Min Zhu, E Xu, Jin Niu

TL;DR
This paper reviews how three gut bacteria help protect against pathogens, support gut health, and improve outcomes for various diseases.
Contribution
A comprehensive summary of the probiotic properties of three specific Lactobacillus species and their health benefits.
Findings
These bacteria enhance gut barrier function by upregulating tight-junction proteins and mucin secretion.
They reduce inflammation and provide protective effects against conditions like diarrhea, obesity, and colorectal cancer.
More research is needed to fully understand the mechanisms of these probiotic strains.
Abstract
As the critical component of the gastrointestinal tract, which lives in trillions of gut microorganisms, in a healthy state, the host interacts with the gut microbiota and is symbiotic. The species Limosilactobacillus reuteri, Ligilactobacillus salivarius, and Lactobacillus johnsonii are indigenous gut commensal bacteria that are mainly found in the digestive tracts. These three bacteria possess a variety of characteristics that reflect their ability to adapt to the gastrointestinal environment. Herein, we summarize the current progress of research on the probiotic properties of these strains in terms of their ability to protect against harmful pathogens, maintain intestinal health, and improve disease outcomes. These bacteria can impact the intestinal barrier function and enhance intestinal immunity through various mechanisms, such as upregulating the tight-junction protein expression…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
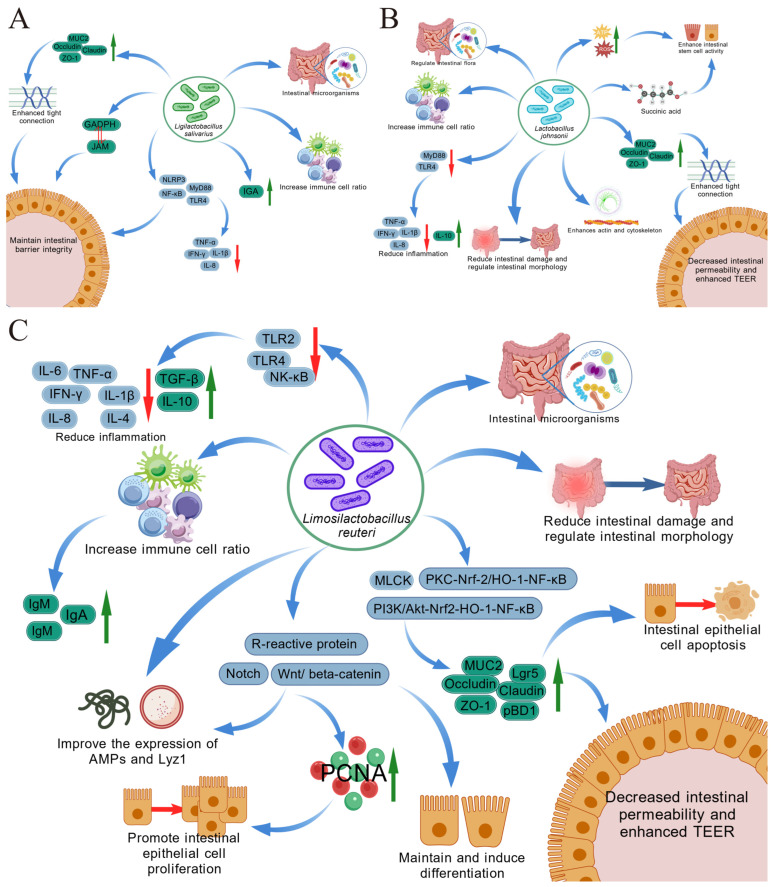 Figure 1
Figure 1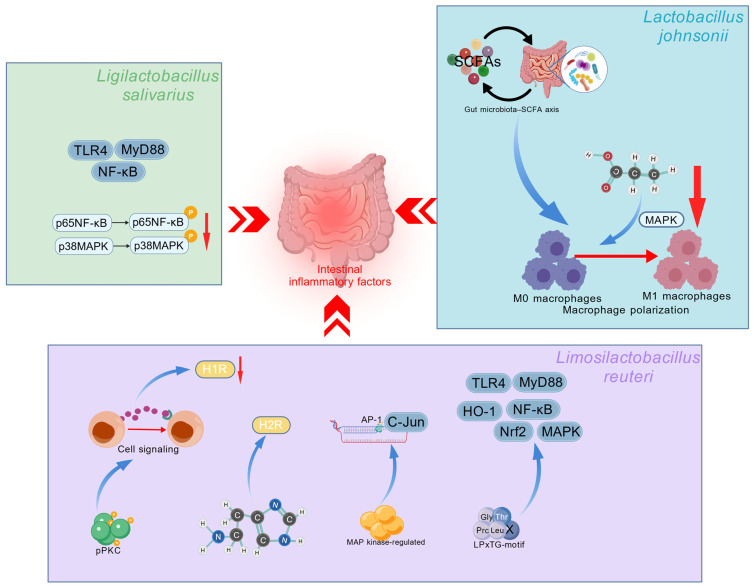 Figure 2
Figure 2- —Natural Science Foundation of China
- —Provincial Key Fund Project of Guizhou Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Gut microbiota and health · Microbial Metabolites in Food Biotechnology
1. Introduction
The gut microbiota is a crucial component of the gastrointestinal tract and plays a key role in maintaining host physiological functions. Many studies indicate that certain diseases in animals and humans, such as inflammatory bowel disease (IBD), obesity, metabolic syndrome, autism, and some cancers, are closely associated with impaired gut microbiota integrity [1,2]. Probiotics are defined as “live microorganisms which when intake in adequate amounts can benefit on host health” by the FAO/WHO [3]. Recently, probiotics have become increasingly frequent in the diet due to their health benefits. Notably, Limosilactobacillus reuteri (formerly named Lactobacillus reuteri, L. reuteri), Ligilactobacillus salivarius (formerly named Lactobacillus salivarius, L. salivarius), and Lactobacillus johnsonii (L. johnsonii) are widely used in human and animal production because of their beneficial properties [4]. L. reuteri can produce antimicrobial molecules, such as organic acids, ethanol, and reuterin [5]. L. johnsonii has many metabolites, including short-chain fatty acids, bacteriocins, and hydrogen peroxide [6]. L. salivarius is a well-characterized bacteriocin producer and probiotic organism, many of which are producers of unmodified bacteriocins of sub-classes IIa, IIb, and IId [7]. L. salivarius also exhibits tolerance to acid and bile salts, thus allowing for a higher survival rate in the gastrointestinal tract (GIT) [8].
L. reuteri has multiple probiotic effects on humans and animals, including intestinal colonization, host immune system regulation, broad-spectrum antimicrobial compound production and secretion, and prevention of diarrhea and colitis. Furthermore, L. reuteri has been used to study the intestinal commensal bacteria ecology and evolution model in vertebrates [9]. L. johnsonii has been used as a probiotic to treat diseases, showing specific advantages in the treatment of a variety of diseases, and can guarantee the health of production and daily life [10]. L. salivarius enhances immune function, inhibits pathogen colonization, increases animal production, and can also be used to treat chronic diseases [11]. They play an important role as probiotics and have great application prospects in biomedicine and livestock production.
In the present review, we summarize and discuss the current progress of research on the probiotic properties of these strains in terms of their ability to protect against harmful pathogens and maintain intestinal health, and their function in disease improvement, which hopefully will provide a reference for the effective application of these strains in the clinical and nutritional fields.
2. Inhibits Pathogens and Ameliorates Damage
Pathogen infections are a serious and basic category of diarrhea, inflammation, and other organ damage caused by pathogen invasion. These pathogens that cause infections are mainly viruses and bacteria [12]. One of the most important characteristics of probiotics is that they can inhibit pathogenic microorganisms and bring beneficial effects to the host [13]. Herein, we summarize the effects of different strains of L. reuteri, L. salivarius, and L. johnsonii on the main pathogenic factors and their possible mechanisms (Table 1).
From Table 1, we can see that these strains were isolated from different hosts, including humans, pigs, chickens, calves, and lambs. Based on the available information, we could find L. reuteri, L. salivarius, and L. johnsonii inhibit both viruses and bacteria; they all inhibit E. coli, Salmonella, H. pylori, and C. jejuni. What is special is that L. reuteri had the ability to protect P. gingivalis, E. faecalis, F. verticillioides, C. albicans, Shigella sonne, Shigella flexneri, Vibrio cholerae, PCV2, and Influenza A/PR8, LD50 infection. L. johnsonii inhibited WSSV, Endogenous pathogenic bacteria, RSV, and Clostridium perfringens, especially for L. salivarius, which can protect Aspergillus hydrophilus, Cronobacter sakazakii, Infectious, IBDV, Uremic toxins, Mycotoxin, and HIV-1 infection. The mechanisms by which L. reuteri, L. salivarius, and L. johnsonii inhibit pathogens are summarized in Table 1. We can observe that they fight against pathogen infection, mainly through producing antimicrobial molecules, regulating the intestinal microbiota, reducing the inflammatory response, and enhancing host defense.
3. Maintains Intestinal Health and Homeostasis
The gut is the largest immune organ and is involved mainly in digestion, absorption, immune homeostasis, endocrine regulation, and other physiological functions, and its health is a direct response to the characteristics of the organism [113]. Gut autochthonous commensal L. reuteri, L. salivarius, and L. johnsonii are utilized mainly as food or food supplements, and the gastrointestinal tract is an important part of their function. Understanding their effects on the intestinal tract allows us to visualize their functions.
3.1. Improves the Intestinal Barrier
The intestinal barrier is an important component of intestinal immunity, and L. salivarius, L. johnsonii, and L. reuteri can modulate intestinal immune responses by affecting the intestinal barrier. The intestinal barrier consists of four barriers, including mechanical, chemical, microbial, and immune barriers. Different mechanisms underlying the impacts of L. salivarius, L. johnsonii, and L. reuteri strains on the intestinal barrier is demonstrated in Figure 1.
Firstly, L. salivarius, L. johnsonii, and L. reuteri strains improve the intestinal barrier by impacting the tight-junction protein expression. For example, L. salivarius YL20 has been demonstrated to inhibit the invasion of Chlamydia sakazaki infection in the HT-29 and Caco-2 monolayer cell model, increasing ZO-1 and occludin expression. This strain also enhances intestinal barrier function by increasing the goblet cell count, MUC-2 levels, and ZO-1 expression in mouse intestinal organoids [87]. L. salivarius LI01 can effectively restore the intestinal barrier biomarkers Claudin-1 and MUC2 expression [115]. L. salivarius CPU-01 upregulated the colonic ZO-1 and Occludin expressions [116]. L. salivarius CML352 significantly increased Muc-2 to improve the intestinal barrier [117]. L. salivarius SNK-6 has been shown to upregulate the jejunum intestinal barrier-related mRNAs ZO-1, CLDN1, and MUC2 expressions [118]. About L. johnsonii, the results showed that L. johnsonii strain MG enhanced the intestinal barrier integrity through the interaction between GAPDH and the junctional adhesion molecule-2 (JAM-2), which showed that GAPDH interacts and docks with JAM-2 by the two peptides, 11GRIGRLAF18 at the N-terminus, and 323SFTCQMVRTLLKFATL338 at the C-terminus [119,120]. L. johnsonii MG pretreatment enhanced intestinal barrier function, protected against Enterococcus faecium-induced damage, and increased ZO-1 expression of Caco-2 cells [121]. Treatment with L. johnsonii N5 increased the expression levels of ZO-1, closure protein, and cytoprotective HSP70 under physiological conditions, and it alleviated colitis [122]. L. johnsonii L531 alleviated Salmonella typhimurium-induced damage through activating tight-junction protein expression and inhibiting the TLR4/NF-κB/NLRP3 signaling pathway [123]. L. reuteri strains LR1, DSM 17938, and 1563F had a protective effect against ETEC-induced damage to the mucosa, mainly through increased expression of ZO-1 [124,125,126]. L. reuteri 2892 upregulated the expression of tightly connected molecules ZO-1 and claudin-4 and inhibited the expression of metalloproteinase (MMP-2) and MMP-9 [127]. Supplementation with L. reuteri 81 significantly increased the gene expression of the ileum tight-junction protein ZO-1 [128]. Administration of L. reuteri 4569 improved the expression of barrier-protective tight-junction protein (TJ) and cell-protective heat shock protein (HSP) 70 and HSP25 [129]. L. reuteri B1/1 was even able to increase those of the tight-junction-related genes CLDN1 and OCLN [130]. In this sense, several studies have found that different strains of L. salivarius, L. johnsonii, and L. reuteri improved tight-junction protein, thereby enhancing intestinal barrier function.
Secondly, L. salivarius, L. johnsonii, and L. reuteri strains can also improve the intestinal barrier through regulating the intestinal flora. It has been reported that L. salivarius CPU-01 and LS160 regulated the intestinal flora, maintained the intestinal structure, and enhanced intestinal barrier function [116,131]. L. salivarius LI01, CML352, and SNK-6 significantly altered the Firmicutes/Bacteroides ratio to protect the intestinal barrier structure [117,118,132]. L. Salivarius PS21603 reduced Escherichia, and increased Bifidobacterium to improve the intestinal barrier in piglets [133]. A consortium of L. salivarius 7247 and L. curvatureus 2029 strains prevented intestinal barrier dysfunction caused by Campylobacter jejuni, maintained transepithelial resistance of intestinal cell monolayers, and prevented permeability of intestinal epithelial cells [134]. L. johnsonii 6084 mitigated a decrease in gut microorganism diversity and abundance, restored the Mycobacterium anisopliae and the Aspergillus phylum abundance in LPS-treated mice, and adjusted the balance of gut microorganisms as a means of enhancing intestinal barrier function [93]. Reports indicate that L. reuteri NK33 alleviates gut microbiota imbalance by reducing the number of Proteus bacteria and increasing the number of Clostridium bacteria [135]. L. reuteri DSM17938 regulated the gut microbiota, increasing bacterial diversity to protect the intestinal barrier [136]. Synthetic bacteria composed of L. reuteri and inulin protected the integrity of the intestinal barrier, altered the composition of the intestinal microbiota, and increased the abundance of Bifidobacterium butyricum [137]. L. reuteri KUB-AC5 could lead to the enrichment of potentially beneficial lactic acid bacteria and inhibit Proteobacteria, including nonbeneficial bacteria, to protect the intestinal barrier [138]. L. reuteri ZJF036 increased the Firmicutes and Clostridia and was also found to decrease the Firmicutes to Bacteroides ratio. The Lactobacillus increased, and the Turicibacter and Blautia decreased to protect the intestinal barrier [139]. L. reuteri FYNDL13 could promote the formation of butyric acid, upregulate the transcription of antimicrobial peptide-encoding genes, and prevent hyperimmune reactions around and in the intestine. In addition, it increased the beneficial bacteria abundance (including Bifidobacteria, Akkermania, Cyanobacteria, and Spirochaeta oscillosum) and limited the relative abundance of harmful bacteria (Bacteroides and Subelia) to protect the intestinal barrier [140]. L. reuteri CCFM1175 effectively reduced CAP-induced damage to the ileum and colon by increasing the UCG_014 and Ackermannia in Ruminococcaceae [141]. Thus, L. salivarius, L. johnsonii, and L. reuteri strains improved the intestinal barrier by inhibiting harmful bacteria and increasing beneficial bacteria in the gut.
Thirdly, L. salivarius, L. johnsonii, and L. reuteri strains improve the intestinal barrier by regulating the intestinal immune function. L. salivarius 160 upregulated the intestinal IL-6 and TLR2 expression to maintain the intestinal barrier integrity [131]. The addition of L. salivarius CML352 decreased the My-D88, IFN-γ, and TLR-4 to improve the intestinal barrier [110,117]. L. johnsonii BS15 improved intestinal immunity and enhanced intestinal mucosal immunity by increasing the level of sIgA [142,143]. L. reuteri LR1 increased the contents of sIgA, porcine β-defensin 2, and protein 1-5 transcripts of ileum secretory immunoglobulin A to improve intestinal barrier function of weaned pig ileum mucosa [144]. L. reuteri f041 could promote intestinal sIgA production and antimicrobial peptide-related gene expression and enhance the mucosal barrier function [145]. In LPS-stimulated mice, L. reuteri ZJ617 and ZJ615 modulated intestinal immune responses and metabolism [146]. L. reuteri 22 inhibited the Notch signaling pathway, increased the expression of mucin 2, and improved intestinal mucosal immunity [147]. Bovine L. reuteri RGW1 increased the levels of TGF-β and IL-10, and its cell-free supernatant (RCS) decreased the serum TNF-α levels [148]. L. reuteri D8 enhanced the intestinal mucosal barrier through increasing goblet cells and antimicrobial peptides (AMPs) related gene expression, including Muc2, Lyz1, and pBD1 [149]. L. reuteri ATCC 55,730 administration induced CD4-positive T lymphocytes in the ileal epithelium of a human intestinal tract [150]. The BBC3-derived ev (lrev) of L. reuteri inhibited the Th1- and Th17-mediated inflammatory response by inhibiting NF-κB activity. The activation of macrophages enhanced immunoregulatory cell-mediated splenic lymphocyte immunosuppression and maintained intestinal immune homeostasis [151]. L. reuteri TPC32 could enhance intestinal biochemical and physical barrier functions by increasing sIgA expression [152]. In a mouse model of alcoholic leaky gut, L. reuteri 3632 modulated gut and immune homeostases by activating aryl hydrocarbon receptors (AhR) [153]. L. salivarius, L. johnsonii, and L. reuteri strains played important roles in maintaining intestinal immune homeostasis, especially L. reuteri, which has been reported more frequently.
Furthermore, there are still other mechanisms by which L. salivarius, L. johnsonii, and L. reuteri strains improve the intestinal barrier. L. salivarius SMXD51 enhanced intestinal barrier function by increasing transepithelial electrical resistance (TEER) and strengthening the F-actin cytoskeleton [84]. L. salivarius 7247 and the Limosilactobacillus fermentum 3872 consortium also protected intestinal barrier functions through increasing TEER and inhibiting paracellular permeability in monolayers of human and animal enterocytes [154]. L. reuteri NPL-88 could increase the TEER [155], L. reuteri I5007 increased the protein expression of intestinal epithelial protein-1 in newborn piglets and maintained the IPEC-J2 cells’ TEER, and its supernatant inhibited an increase in TNF-α and IL-6 expression and a decrease in TJ protein expression induced by LPS [156]. L. reuteri 100-23 stimulates the development of regulatory T cells [157]. The specific cell-free fermentation supernatant (CFS) of L. reuteri G7 promoted the growth and proliferation of the intestinal epithelial cell line Caco-2 and enhanced the intestinal barrier [158]. In rats and mice, L. salivarius LI01 reduces serum endotoxin and bacterial translocation, improves intestinal ultrastructure damage, and maintains the intestinal barrier [159]. L. reuteri ZJ617 protected against LPS-induced intestinal barrier dysfunction by enhancing antioxidant activity through the mTOR signaling pathway [160]. Administration of L. johnsonii YH1136 ameliorates high-altitude hypoxia-induced intestinal injury by regulating Staphylococcus and Corynebacterium cooperated with miR-196a-1-3p and miR-3060-3p, respectively [60]. Extracellular vesicles derived from L. johnsonii (LJ-EVs) could effectively prevent colitis symptoms because LJ-EVs could be directly absorbed by intestinal epithelial cells, activate the Nrf2/HO-1 antioxidant signaling pathway, reduce endotoxin damage to cells, and thus maintain intestinal barrier homeostasis [161]. Whether there are other mechanisms of action remains to be studied.
Overall, L. salivarius, L. johnsonii, and L. reuteri can ameliorate the intestinal barrier, enhance intestinal immune function, and protect the organism from external influences by increasing intestinal epithelial tight-junction protein expression and mucin production, adjusting and balancing the level of the intestinal microflora, blocking pro-inflammatory cytokines, and so on.
3.2. Alleviate Intestinal Inflammatory Response
Systemic inflammation is a natural, protective biological response of the host immune system that fights off harmful foreign pathogens (including bacteria, viruses, and toxins) and helps the body restore health.
L. salivarius, L. johnsonii, and L. reuteri strains reduce intestinal inflammation mainly through regulating inflammatory cytokine expression and secretion. For instance, L. salivarius ZLP-4b could improve the intestinal morphology of mice, increase the contents of the anti-inflammatory cytokines IL-4 and IL-10, and decrease the content of the pro-inflammatory factor IL-17A [162]. L. salivarius UCC118 intervention attenuated the secretion of IL-8 and the pro-inflammatory response induced by Salmonella typhimurium and stimulated the secretion of IL-10 and TNF-α by dendritic cells (DCs) to mediate the response of intestinal pathogens and play an immunomodulatory role [32]. Both L. salivarius CCFM 1266 and L. salivary UCC118TM alleviated colon inflammation by increasing M2 macrophage polarization and anti-inflammatory IL-10 production [163]. L. salivarius CNCM I-4866 reduced the expression of markers of colon injury and inflammation. It could also exert anti-inflammatory effects by reducing the amount of IL-8 produced by TNF-α-stimulated cells and regulating cytokine profiles in peripheral blood mononuclear cells (PBMCs) [17]. Combined treatment with L. salivarius Li01 and Bifidobacterium longum TC01 more effectively reduced the TNF-α, MCP-1, and M-CSF levels, thereby inhibiting systemic inflammation in rats [115]. The combination of RJGP16 and L. salivarius B1 treatment significantly increased the levels of IL-6 and porcine β-defencin (pBD)-2 in the duodenum and ileum [164]. L. johnsonii L531 preincubation induced the expression of pro-inflammatory cytokines [123]. L. johnsonii N5 inhibited the intestinal TNF-α and IL-6 production, and increased the intestinal Peyer’s patch MHCII and CD103 dendritic cell populations and the number of regulatory T cells, and thus reduced the production of the Th17 population and IL-17a production during colitis to increase the expression of IL-10 to ameliorate colonic inflammation [122]. L. reuteri plays a very important role in regulating inflammatory cytokines in vivo and in vitro. L. reuteri LM1071 displays potential anti-inflammatory capacity, which is achieved by inhibiting the production of inflammatory mediators such as NO, arachidic acids like PGE1 and PGE2, pro-inflammatory cytokines, and COX proteins [165]. Furthermore, it can increase the expression of inflammation-related genes such as IL-11, BMP4, LEFTY2, and EET metabolites production. L. reuteri MT180537 upregulated the anti-inflammatory cytokines and resulted in the control of E. faecalis-MW051601-induced overexpression of pro-inflammatory cytokines [166]. L. reuteri ATG-F3 (F3) and F4 strains exhibited anti-inflammatory effects on RAW264.7 mouse macrophages, and mice orally administered with the F4 strain showed increased ileum IL-10 production [167]. L. reuteri F-9-35 reduced the colon tissue pro-inflammatory gene expression [168]. The soluble factor of L. reuteri CRL1098 significantly decreased the production of NO, COX-2, Hsp70, TNF-α, and IL-6 in LPS-stimulated macrophages [169]. In children with active distal ulcerative colitis, rectal instillation of L. reuteri ATCC 55,730 effectively improved mucosal inflammation through increasing IL-10 levels and decreasing IL-1β, TNF-α, and IL-8 levels in the mucosa [170]. Specific strains of L. reuteri L3 and L8 were identified in control mice and obese mice, respectively. L. reuteri L8 induced the production of IL-6, IL-12, and TNF-α, while L. reuteri L3 induced IL-10 production [171]. L. reuteri 5454 efficiently triggered IL-22 secretion and regulatory T-cell induction in dendritic cells [172]. L. ruminis and L. reuteri LY2-2, through regulating pro-inflammatory cytokines production, alleviated DSS-induced colitis [173]. These results illustrated that the L. salivarius, L. johnsonii, and L. reuteri strains regulated the intestinal inflammatory response by decreasing anti-inflammatory factors and reducing pro-inflammatory factors.
The signaling pathways involved in the L. salivarius, L. johnsonii, and L. reuteri strains regulate inflammatory cytokines (Figure 2). Research has shown that L. salivarius reduced inflammation-related factors by decreasing the p38 MAPK and p65 NF-κB phosphorylation of IPEC-J2 cells [16]. L. salivarius WZ1 inhibited the jejunum inflammatory damage induced by ETEC K88 via the TLR4/NF-κB/MyD88 pathways [15]. L. johnsonii inhibited pro-inflammatory cytokines secretion mainly by gut microbiota-derived short-chain fatty acids, suppressing the M1 macrophages polarization [15]. In particular, the increased concentration of propionic acid in the gut inhibited the MAPK signaling pathway activation of macrophages, thereby reducing the polarization of M1 macrophages [174,175]. L. reuteri DSM 8533 inhibited TNF-α and IL-1β production by regulating the ERK-JNK-related MAPK signaling cascade through LPxTG-motif surface protein [176]. L. reuteri GroEL and LrPGN seemed to inhibit inflammation by the activation of a noncanonical TLR4 pathway [85,177]. L. reuteri 17938 exerted its anti-inflammatory effect through the TLR2 pathway [178]. L. reuteri 6475 attenuated pPKC-mediated mammalian cell signaling to inhibit the pro-inflammatory response mediated by H1R in the gut [179]. In lipopolysaccharide-activated monocytes and primary monocytes derived from children with Crohn’s disease, the probiotic L. reuteri strain ATCC PTA 6475 inhibited TNF transcription via suppressing MAP-regulated c-Jun and activating transcription factor AP-1 [180]. L. reuteri LR1 could activate the MLCK signaling pathway of IPEC-J2 cells to inhibit ETEC k88 challenge [181]. L. reuteri SH 23-derived LPxTG-motif surface protein had the function to alleviate inflammatory diseases through the NF-κB pathway [182]. L. reuteri that carries the amino acid decarboxylase gene converted L-histidine into histamine in the intestinal lumen, activating H2R, which ultimately inhibits acute inflammation in the mouse colon [183]. L. reuteri RE225 reduces mice inflammation by inhibiting the TLR4/MyD88/NF-κB and Nrf2/HO-1pathways [184]. These findings improve our understanding and knowledge of how L. salivarius, L. johnsonii, and L. reuteri reduce intestinal inflammation and provide new insights for human disease and animal treatment target identification.
4. Relieving the Progression of Diseases
In animal models and clinical trials, it has been found that L. salivarius has been used to prevent and treat multiple human chronic diseases, including asthma, cancer, atopic dermatitis, and halitosis, and has also been used for infection prevention or treatment [11]. For example, L. salivarius LI01 and L. salivarius LI02 have been demonstrated to prevent acute liver failure in rats [185]. L. salivarius WB21 inhibited the quantity of oral periodontopathic bacteria, such as Porphyromonas gingivalis, P. intermedia, Tannerella forsythensis, and Fusobacterium nucleatum [186]. L. salivarius LI01 plays a protective role against thioacetamide (TAA)-induced acute liver injury and hyperammonemia in mice [187]. A 14-day oral administration of L. salivarius K12 was conducted on hospitalized COVID-19 patients and confirmed the hypothesis that oral microbiota directly participates in the lung microbiota establishment, significantly reducing the mortality rate associated with COVID-19 infection [188]. Nadja Larsen et al. found that L. salivarius Ls-33 modified the fecal microbiota in obese adolescents in a way not related to metabolic syndrome [189].
The patient underwent surgical resection of <1 cm, removing all visible lesions in the past 21 days, and received encapsulated freeze-dried L. johnsonii LA1 (2 × 10^9^ CFU) twice daily, resulting in a reduction of endoscopic recurrence from 64% to 49% [190]. L. johnsonii N6.2 reduced the oxidative response protein expression (i.e., Gpx1, GR, and Cat) in the intestinal mucosa and inhibited the onset of type1 diabetes in rats [191]. Xin et al. reported that L. johnsonii BS15 inhibited non-alcoholic fatty liver disease (NAFLD)-associated insulin resistance in mice, and reduced the gene expression of acetyl-CoA carboxylase 1 (ACC1), fatty acid synthase (FASN), and peroxisome proliferator-activated receptor gamma (PPARγ); it also increased the expression of fasting-induced adipokines in the liver of obese mice [192]. L. johnsonii BS15 alleviated abnormal mitochondrial by decreasing uncoupling protein 2 and increasing cytochrome C levels in diabetes [193]. Yin et al. found that heat-killed L. johnsonii (HKLJ) upregulated the intestinal lysozyme expression in alcohol-related liver disease, and enhanced intestinal bacteria-mediated immunoregulatory substances production [194]. This activation leads to the NOD2-IL-23-IL-22 innate immune axis activation, and elevated IL-22 upregulates the synthesis of antimicrobial peptides to maintain intestinal homeostasis. Additionally, HKLJ also activates the liver signal transducer and activator of the transcription 3 (STAT3) pathway and promotes liver damage repair [195].
A review summarized that L. reuteri was used as a probiotic for the treatment of functional abdominal pain, diarrhea, constipation, H. pylori infection, IBD, diverticulitis, colorectal cancer, and liver disease [196]. L. reuteri also contributes to alleviating depression, which has been confirmed in studies involving both mice and humans [48,197]. Samiraninezhad et al. found that a probiotic nanomedicine containing L. reuteri treatment with recurrent aphthous stomatitis for one week significantly increased the lesion size and reduced the severity of pain compared to the control group [198]. In the first use of L. reuteri selenium nanoparticles for the treatment of ulcerative colitis in mice, the prepared SeNPs effectively alleviated symptoms such as diarrhea, weight loss, bloody stools, and colon shortening [199]. The decrease in the number of L. reuteri in the human body has been positively correlated with the increase in the incidence of inflammatory diseases over the same period. Direct supplementation of L.reuteri or the regulation of its content through prebiotics is a promising method for the prevention and/or treatment of diseases.
Taken together, L. salivarius, L. johnsonii, and L. reuteri strains play a significant and important role in promoting human health. They could provide certain protective effects against various health issues in humans, such as diarrhea, constipation, colorectal cancer, and liver diseases. Additionally, they have been found to have the potential to suppress the onset of diabetes. However, their role in human clinical research is still limited (Table 2). Therefore, developing these three probiotics as therapeutic agents for human diseases has broad research prospects.
5. Summary and Outlook
L. salivarius, L. johnsonii, and L. reuteri strains can regulate the intestinal barrier and enhance intestinal immune function through various mechanisms, such as increasing the expression of tight-junction proteins and mucin secretion in intestinal epithelial cells, adjusting and balancing intestinal microbiota, and blocking pro-inflammatory cytokine production. They have been shown to reduce intestinal inflammation in multiple animal models and provide protective effects against various health issues in humans, such as diarrhea, constipation, colorectal cancer, and liver diseases.
However, the detailed mechanism of some strains remains unclear, and new and powerful strains still need to be isolated and identified. The probiotic properties depend on their genome, and new microbiological strategies, such as mutational breeding of microbial strains, can better enhance the probiotic properties of strains lacking genetic manipulation systems. In addition, fecal metagenomic sequencing, such as exfoliome sequencing (Foli-seq), was used to profile fecal exfoliated eukaryotic messenger RNAs (feRNAs) originating from the upper and lower gastrointestinal regions, and metabolomics studies have revealed the impact of the gut microbiota on health and disease. Microorganisms have unique and efficient biotransformation capabilities and can produce a variety of metabolites. With the development of metabolomics, more metabolites of L. salivarius, L. johnsonii, and L. reuteri will be identified and produced by synthetic biology technology in the future, and both probiotics and their products have broad application prospects in the fields of biology, medicine, and food and animal feed production.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1de Araújo Boleti A.P. de Oliveira Cardoso P.H. Frihling B.E.F. de Moraes L.F.R.N. Nunes E.A.C. Mukoyama L.T.H. Biembengute M.E.F. de Melo V.C.B. Morales M.F. de Castro A.P. Immune dysregulation and gut microbiota: Connection to health and disease development Neural Regen. Res.2025 ahead of print 10.4103/NRR.NRR-D-25-0024441017678 · doi ↗ · pubmed ↗
- 2De Luca R. ArrèV. Nardone S. Incerpi S. Giannelli G. Trivedi P. Anastasiadou E. Negro R. Gastrointestinal microbiota and inflammasomes interplay in health and disease: A gut feeling Gut 20267516117510.1136/gutjnl-2025-334938 PMC 1270332840441864 · doi ↗ · pubmed ↗
- 3Gibson G.R. Hutkins R. Sanders M.E. Prescott S.L. Reimer R.A. Salminen S.J. Scott K. Stanton C. Swanson K.S. Cani P.D. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics Nat. Rev. Gastroenterol. Hepatol.20171449150210.1038/nrgastro.2017.7528611480 · doi ↗ · pubmed ↗
- 4Kumar S. Chauhan N. Chauhan T. Balaga S. Tyagi N. Samanta A.K. Evaluation of the techno-functional properties of lactobacilli strains originated from Bos indicus and Bubalus bubalis calves for probiotic potential Int. Microbiol.2025281649166810.1007/s 10123-025-00641-y 39982624 · doi ↗ · pubmed ↗
- 5Mu Q. Tavella V.J. Luo X.M. Role of Lactobacillus reuteri in human health and diseases Front. Microbiol.2018975710.3389/fmicb.2018.0075729725324 PMC 5917019 · doi ↗ · pubmed ↗
- 6Zhao M. Li Y. Zhang Y. Li G. Genomic analysis and functional properties of Lactobacillus johnsonii GJ 231 isolated from healthy beagles Front. Microbiol.202415143703610.3389/fmicb.2024.143703639355429 PMC 11442259 · doi ↗ · pubmed ↗
- 7Messaoudi S. Manai M. Kergourlay G. Prévost H. Connil N. Chobert J.-M. Dousset X. Lactobacillus salivarius: Bacteriocin and probiotic activity Food Microbiol.20133629630410.1016/j.fm.2013.05.01024010610 · doi ↗ · pubmed ↗
- 8Yang Y. Song X. Wang G. Xia Y. Xiong Z. Ai L. Understanding Ligilactobacillus salivarius from probiotic properties to omics technology: A review Foods 20241389510.3390/foods 1306089538540885 PMC 10969406 · doi ↗ · pubmed ↗