Runx2 in the Perichondrial Osteoblasts Enhances Terminal Differentiation of Chondrocytes Through Nell1 Induction
Xin Qin, Qing Jiang, Longfei Wu, Suemi Yabuta, Chiharu Sakane, Yuki Matsuo, Ziheng Zhang, Hisato Komori, Manyu Zhang, Kosei Ito, Toshihisa Komori

TL;DR
This study shows that Runx2 in perichondrial osteoblasts promotes chondrocyte maturation by increasing Nell1 expression.
Contribution
The novel finding is that Runx2 enhances terminal chondrocyte differentiation through Nell1 induction in perichondrial osteoblasts.
Findings
Runx2 in perichondrial osteoblasts promotes chondrocyte maturation via Nell1 induction.
Nell1 expression is upregulated by Runx2 overexpression and downregulated by Runx2 siRNA.
Runx2fl/−Cre mice show delayed chondrocyte differentiation and reduced mineralization.
Abstract
Runx2 plays essential roles in osteoblast differentiation and chondrocyte maturation. Runx2 in the perichondrium has been reported to inhibit chondrocyte maturation through Fgf18 induction. To further investigate the functions of Runx2 in the perichondrium, we generated Runx2fl/−Cre mice by crossing Runx2fl/+, Runx2+/−, and 2.3-kb Col1a1 Cre mice and compared them with Runx2fl/− mice at E15.5, when the endochondral bones were cartilaginous. Skeletal preparation of the upper limbs in Runx2fl/−Cre mice showed reduced mineralization of the humerus and scapula, and histological analysis of the femurs showed delays in the terminal differentiation of chondrocytes, as indicated by the absence of mineralization and Spp1 expression in the cartilage and osteoblast differentiation in the perichondrium, compared to those in Runx2fl/− mice. mRNA sequence analysis showed that the expression of Nell1,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
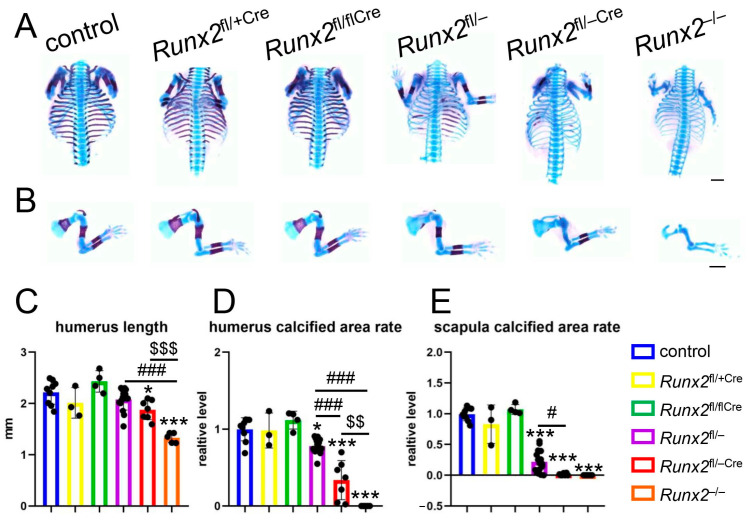 Figure 1
Figure 1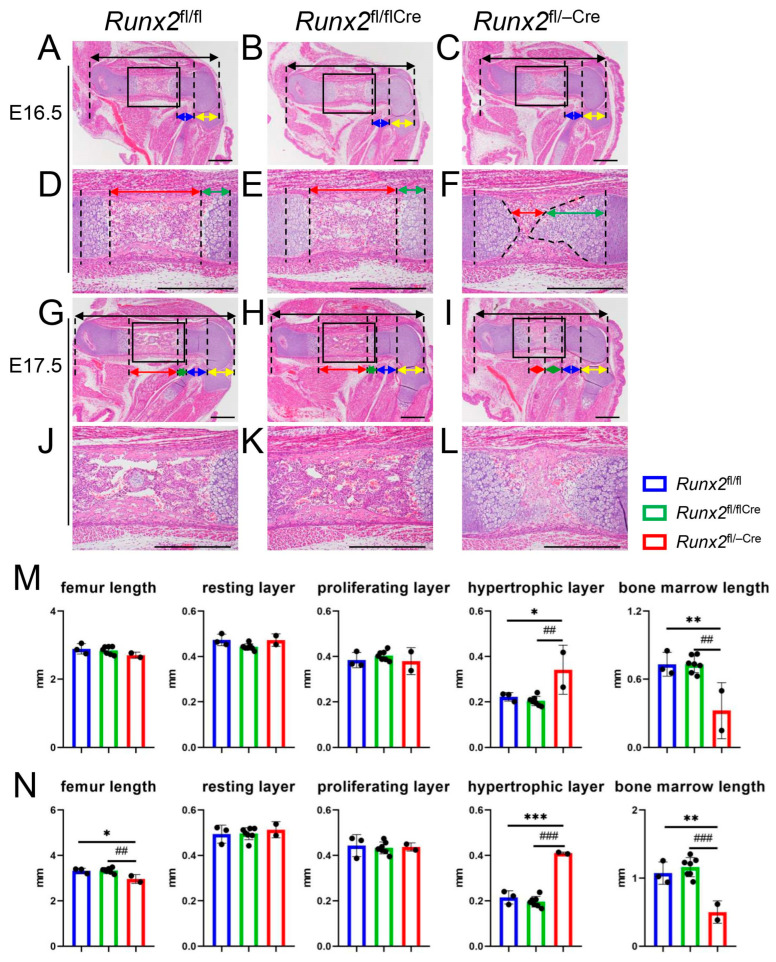 Figure 2
Figure 2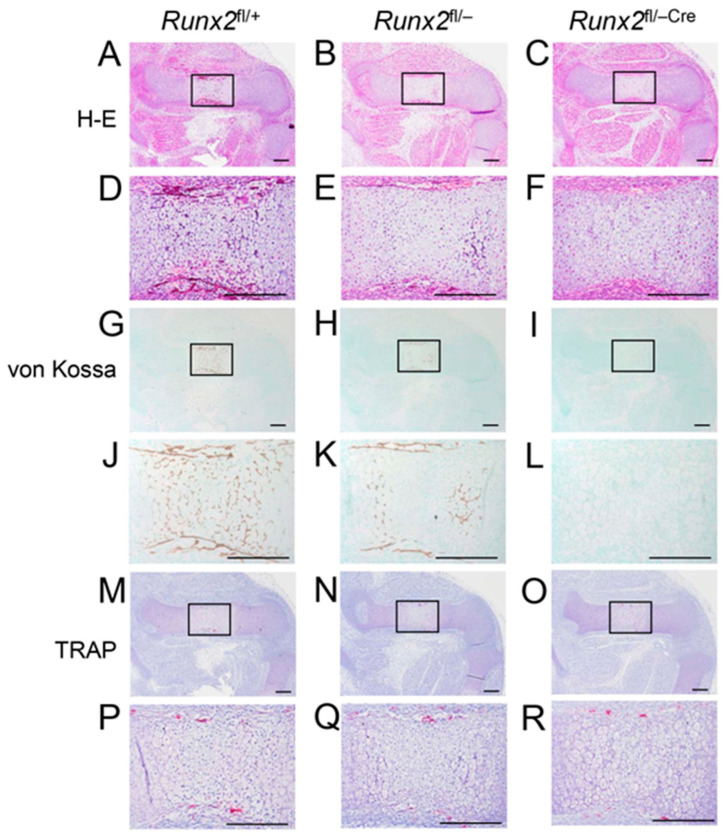 Figure 3
Figure 3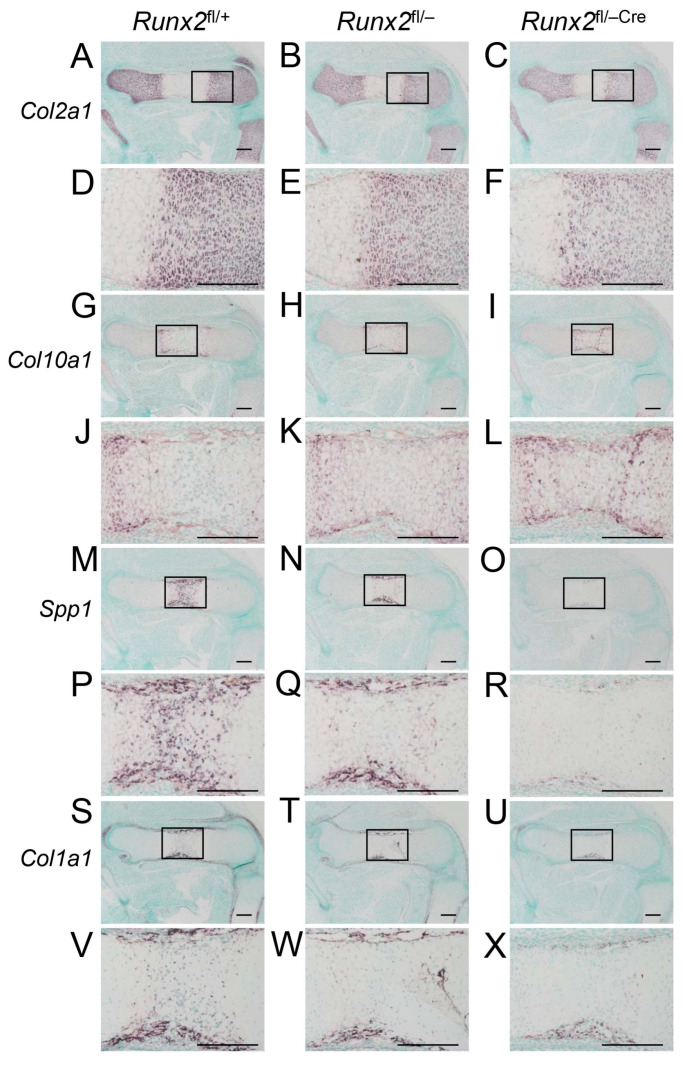 Figure 4
Figure 4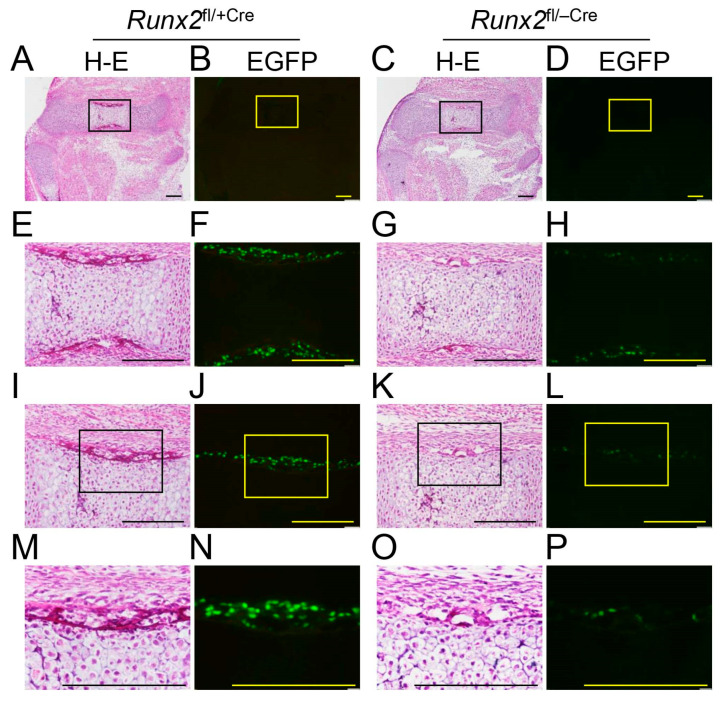 Figure 5
Figure 5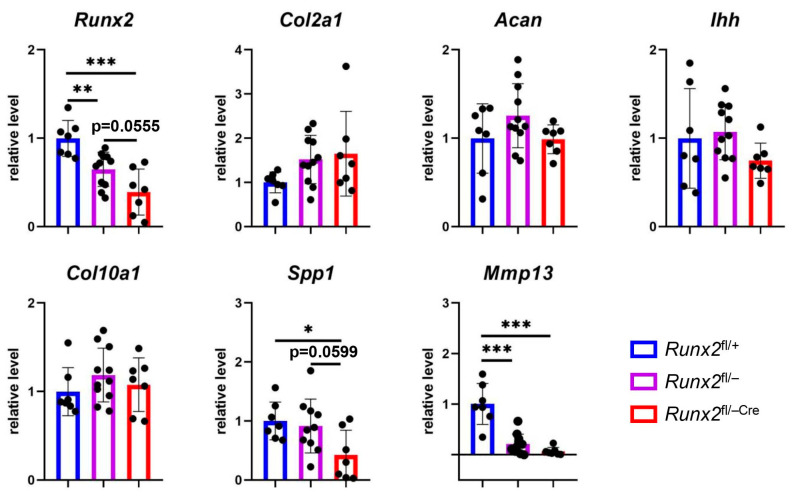 Figure 6
Figure 6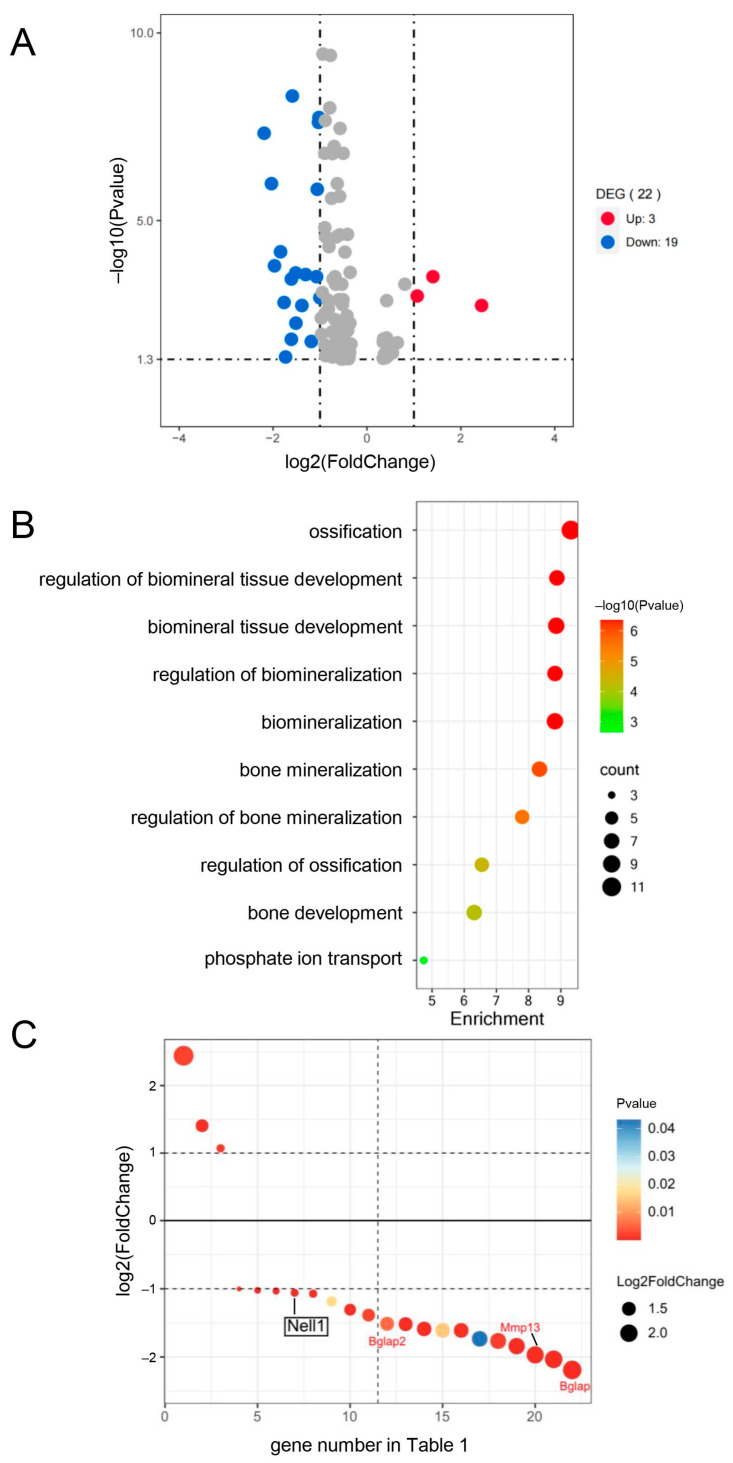 Figure 7
Figure 7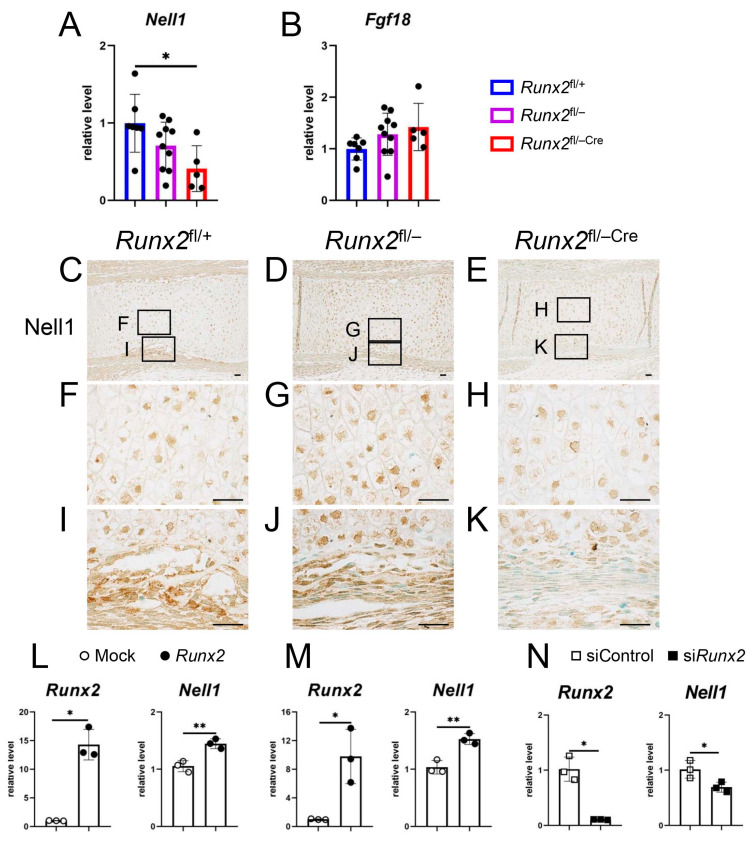 Figure 8
Figure 8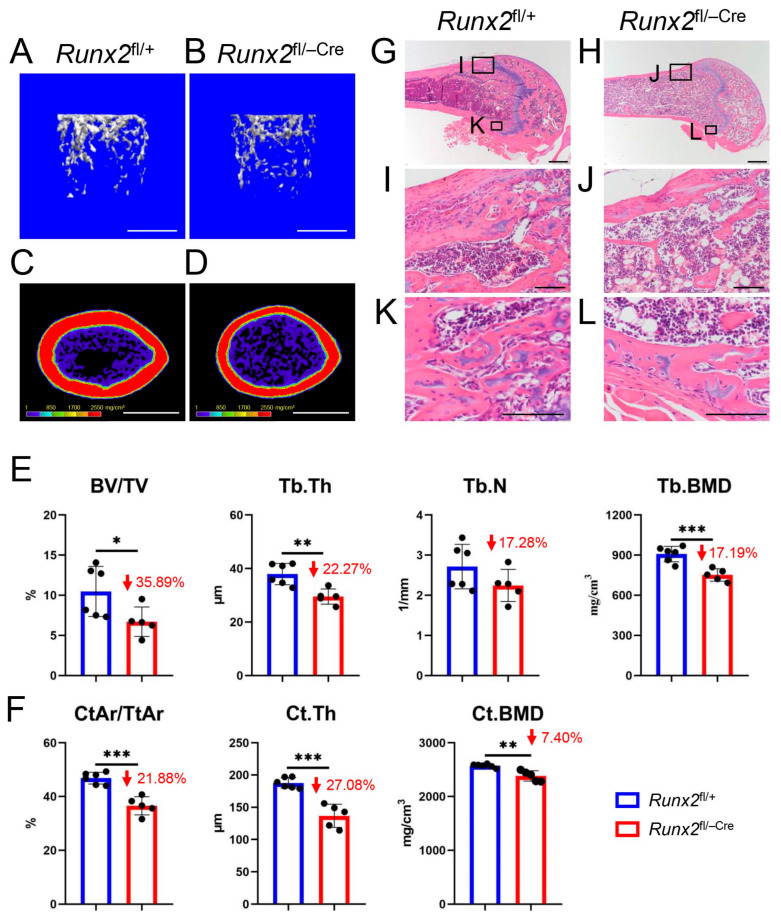 Figure 9
Figure 9- —Japanese Ministry of Education, Culture, Sports, Science and Technology
- —Suzhou Science and Technology Program Project
- —Natural Science Foundation of Jiangsu Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOsteoarthritis Treatment and Mechanisms · Bone Metabolism and Diseases · Connective Tissue Growth Factor Research
1. Introduction
During endochondral ossification, mesenchymal cells condense and differentiate into chondrocytes, forming cartilaginous skeletons. Chondrocytes proliferate and mature into prehypertrophic, hypertrophic, and terminal hypertrophic chondrocytes. Osteoblast differentiation occurs in the perichondrium surrounding the prehypertrophic, hypertrophic, and terminal hypertrophic chondrocyte layers, forming bone collars. Furthermore, the matrix of the terminal hypertrophic chondrocyte layer is mineralized [1,2]. Runx2 is upregulated in prehypertrophic chondrocytes and induces chondrocyte maturation. Upregulated Runx2 expression is maintained in hypertrophic and terminal hypertrophic chondrocytes. Runx2 induces Ihh expression in prehypertrophic chondrocytes, Ihh induces Pthlh expression in the periarticular growth plate, and Pthlh inhibits chondrocyte maturation by inhibiting Runx2 expression in prehypertrophic chondrocytes through Pth1r, forming a negative feedback loop [3,4,5]. Ihh also induces Runx2 expression and osteoblast differentiation in the perichondrium [5,6,7]. Therefore, chondrocyte maturation induces osteoblast differentiation in the perichondrium.
The perichondrium was shown to inhibit chondrocyte maturation. In the organ culture of chicken tibiotarsi, in which the perichondrium was removed, chondrocyte maturation was accelerated, indicating that the perichondrium inhibits chondrocyte maturation [8]. Furthermore, it has been reported that Runx2 in the perichondrium inhibits chondrocyte maturation by inducing Fgf18 expression [9]. In Fgf18^−/−^ mice, chondrocyte proliferation in the resting and proliferating layers of the growth plate was increased, and the proliferating, prehypertrophic, and hypertrophic chondrocyte layers were elongated; however, the differentiation of hypertrophic chondrocytes into terminal hypertrophic chondrocytes was inhibited, as shown by the reduction in Spp1-expressing terminal hypertrophic chondrocytes, resulting in the retardation of endochondral ossification [10,11]. Although the enlarged Col10a1-positive hypertrophic chondrocyte layer in Fgf18^−/−^ mice was considered to be caused by enhanced chondrocyte maturation [10,11], it was likely due to the enhanced proliferation and inhibited terminal differentiation of chondrocytes. Thus, Fgf18 inhibits chondrocyte proliferation but is not likely to inhibit chondrocyte maturation and enhances terminal differentiation of chondrocytes. Twist1, which is expressed in the perichondrium, interacts with Runx2 and inhibits Runx2 function, and Twist1 mutant mice have shown a reduced Col10a1-positive hypertrophic chondrocyte layer due to decelerated chondrocyte maturation [9]. However, the phenotypes showed that the reduced Col10a1-positive hypertrophic chondrocyte layer was caused by enhanced chondrocyte maturation and endochondral ossification. Thus, the misinterpretation of the phenotypes of Fgf18^−/−^ and Twist1 mutant mice makes it difficult to evaluate Runx2 function in the perichondrium during chondrocyte maturation.
Nel-like molecule-1 (Nell1), a neuroepidermal growth factor-like secreted protein, has been identified as an upregulated gene in coronal synostosis [12]. Overexpression of Nell1 causes craniosynostosis, and Nell1 mutant mice exhibit skeletal defects in the skull, vertebral column, and rib cage [13,14]. Nell1 is expressed in osteoblasts, chondrocytes, and the perichondrium and induces the differentiation of osteoblasts and chondrocytes [13,15,16,17,18,19]. Chondrocyte maturation was inhibited in Nell1 mutants and chondrocyte-specific Nell1 deficient mice, and Nell1 induced chondrocyte maturation [16,17,18]. Moreover, Runx2 directly regulates the Nell1 promoter, cranial defects in Runx2^+/−^ mice were partially rescued by crossing with Nell1 overexpression transgenic mice or adding recombinant Nell1 in the organ culture, and Nell1 expression is regulated by Runx2 in chondrocytes [16,19,20].
We previously reported that Runx2 is required for the expression of major bone matrix protein genes in osteoblasts by generating osteoblast-specific Runx2-deficient mice using 2.3 kb Col1a1-enhanced green fluorescent protein (EGFP)-Cre mice (Runx2^fl/flCre^ mice) [21]. To investigate Runx2 functions in the perichondrium, we generated Runx2^fl/−Cre^ mice and analyzed them at E15.5, when the endochondral bones were cartilaginous. We found that deletion of Runx2 in perichondrial osteoblasts decelerated the terminal differentiation of chondrocytes. Therefore, contrary to a previous report [9], Runx2 in the perichondrium enhances chondrocyte maturation. We investigated the mechanism of the regulation of chondrocyte maturation by Runx2 in the perichondrium and found that Nell1 expression in the perichondrium of Runx2^fl/−Cre^ mice was reduced compared with that of Runx2^fl/−^ mice at E15.5. Overexpression of Runx2 induced Nell1 expression and knockdown of Runx2 reduced it, indicating that Runx2 in perichondrial osteoblasts enhances the terminal differentiation of chondrocytes, at least in part, through the induction of Nell1 expression.
2. Results
2.1. Mineralization of Ribs, Vertebrae, Clavicles, Humeri, and Scapulae in Runx2fl/−Cre Mice Was Less than That in Runx2fl/− Mice at E15.5
We generated Runx2^fl/−Cre^ mice by crossing Runx2^fl/+^, 2.3 kb Col1a1 EGFP-Cre, and Runx2^+/−^ mice. We compared the skeletal preparation of control (Runx2^fl/+^ and Runx2^fl/fl^), Runx2^fl/+Cre^, Runx2^fl/flCre^, Runx2^fl/−^ (or Runx2^+/−^), Runx2^fl/−Cre^, and Runx2^−/−^ mice. The mineralization of the ribs, vertebrae, clavicles, humeri, and scapulae was similar among the control, Runx2^fl/+Cre^, and Runx2^fl/flCre^ mice (Figure 1A,B,D,E). Histological analysis of the femurs also showed that the lengths of the femurs and the resting, proliferating, and hypertrophic chondrocyte layers were similar between Runx2^fl/fl^ and Runx2^fl/flCre^ mice (Supplementary Figure S1). Compared with the control, Runx2^fl/+Cre^, and Runx2^fl/flCre^ mice, the mineralization of the ribs, vertebrae, clavicles, humeri, and scapulae was reduced in Runx2^fl/−^ mice, further reduced in Runx2^fl/−Cre^ mice, and absent in Runx2^−/−^ mice (Figure 1A,B,D,E). The length of the humeri in Runx2^fl/−Cre^ mice but not Runx2^fl/−^ mice was shorter than that in control mice and further reduced in Runx2^−/−^ mice (Figure 1C).
2.2. The Process of Endochondral Ossification Was Retarded in Runx2fl/−Cre Mice Compared with That in Runx2fl/fl and Runx2fl/flCre Mice
Femurs were histologically analyzed at E16.5 and E17.5 (Figure 2). The femur lengths were comparable in Runx2^fl/fl^, Runx2^fl/flCre^, and Runx2^fl/−Cre^ mice at E16.5 (Figure 2A–C,M), whereas those in Runx2^fl/−Cre^ mice were shorter than those in Runx2^fl/fl^ and Runx2^fl/flCre^ mice at E17.5 (Figure 2G–I,N). The lengths of the resting and proliferating chondrocyte layers were comparable in Runx2^fl/fl^, Runx2^fl/flCre^, and Runx2^fl/−Cre^ mice at E16.5 and E17.5, whereas those of the hypertrophic chondrocyte layer in Runx2^fl/−Cre^ mice were longer than those in Runx2^fl/fl^ and Runx2^fl/flCre^ mice (Figure 2A–I,M,N). Furthermore, the length of the bone marrow was severely reduced in Runx2^fl/−Cre^ mice compared to that in Runx2^fl/fl^ and Runx2^fl/flCre^ mice at E16.5 and E17.5 (Figure 2D–L,M,N), indicating that endochondral ossification was retarded in Runx2^fl/−Cre^ mice compared to Runx2^fl/fl^ and Runx2^fl/flCre^ mice.
2.3. Terminal Differentiation of Chondrocytes Was Retarded in Runx2fl/−Cre Mice Compared with Runx2fl/− Mice
Chondrocyte maturation and bone collar formation were examined in the femurs at E15.5 (Figure 3). Chondrocyte hypertrophy, but no vascular invasion, occurred at the diaphysis in Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice (Figure 3A–F). However, mineralization was observed in the diaphysis and bone collar in Runx2^fl/+^ and Runx2^fl/−^ mice but not in Runx2^fl/−Cre^ mice (Figure 3G–L), suggesting that terminal differentiation of chondrocytes and bone collar formation were retarded in Runx2^fl/−Cre^ mice. TRAP-positive cells were observed in the perichondrium of Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice (Figure 3M–R).
In situ hybridization was performed using Col2a1, Col10a1, Spp1, and Col1a1 probes (Figure 4). Col2a1 is expressed in resting and proliferating chondrocytes, Col10a1 is expressed in hypertrophic chondrocytes, Spp1 is expressed in terminal hypertrophic chondrocytes and immature osteoblasts, and Col1a1 is weakly expressed in preosteoblasts and strongly expressed in mature osteoblasts [2]. Col2a1 was similarly detected in resting and proliferating chondrocytes among Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice (Figure 4A–F). Col10a1 expression was detected in the metaphyses of Runx2^fl/+^ and Runx2^fl/−^ mice, whereas it was detected in the diaphyses of Runx2^fl/−Cre^ mice (Figure 4G–L). Spp1 was detected in the terminal hypertrophic chondrocytes in the diaphysis and osteoblasts in the bone collar in Runx2^fl/+^ and Runx2^fl/−^ mice, whereas it was weakly detected only in the bone collar in Runx2^fl/−Cre^ mice (Figure 4M–R). Col1a1 was strongly detected in the bone collar of Runx2^fl/+^ and Runx2^fl/−^ mice but weakly detected in Runx2^fl/−Cre^ mice (Figure 4S–X). In accordance with these data, the number of EGFP-Cre-positive cells in the perichondrium of the femurs was much lower, and the intensity of EGFP-Cre was weaker in Runx2^fl/−Cre^ mice than in Runx2^fl/+Cre^ mice at E15.5 (Figure 5). These findings indicate that the terminal differentiation of chondrocytes and osteoblast differentiation in the bone collar are retarded in Runx2^fl/−Cre^ mice compared to those in Runx2^fl/−^ mice.
2.4. Expression of the Marker Genes for Terminal Hypertrophic Chondrocytes Was Reduced in Runx2fl/−Cre Mice
We examined the expression of chondrocyte marker genes using real-time reverse transcription (RT) polymerase chain reaction (PCR) using E15.5 femurs and tibiae (Figure 6). Runx2 expression in Runx2^fl/−Cre^ mice was lower than that in Runx2^fl/+^ mice and marginally lower than that in Runx2^fl/−^ mice. The expression of Col2a1 and Acan, which are expressed in resting and proliferating chondrocytes, was similar in Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice. The expression levels of Ihh, which is expressed in prehypertrophic chondrocytes, and Col10a1 were comparable among Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice. Spp1 expression in Runx2^fl/−Cre^ mice was significantly and marginally lower than that in Runx2^fl/+^ and Runx2^fl/−^ mice, respectively. The expression of Mmp13, which is mainly expressed in terminal hypertrophic chondrocytes [2], was significantly lower in Runx2^fl/−^ and Runx2^fl/−Cre^ mice than in Runx2^fl/+^ mice, with a greater reduction in Runx2^fl/−Cre^ mice.
2.5. RNA Sequence Analysis Showed That Nell1 Expression Is Downregulated in Runx2fl/−Cre Mice Compared with Runx2fl/− Mice More than Two-Fold
To investigate the reason for retarded terminal chondrocyte differentiation in Runx2^fl/−Cre^ mice, we compared the mRNA sequences extracted from the femurs and tibiae of Runx2^fl/−Cre^ and Runx2^fl/−^ mice at E15.5. One hundred and four genes were significantly changed (p-value < 0.05), three of which were upregulated, and nineteen were downregulated by more than two-fold (Figure 7A). Ossification and related GO terms in the biological process were enriched in the 22 genes, and 4 genes, including Nell1, Bglap2, Mmp13, and Bglap, were included in ossification (Figure 7B,C, Table 1). Although Spp1 expression in Runx2^fl/−Cre^ mice was six times lower than that in Runx2^fl/−^ mice, it was not selected because the p-value was 0.06. EGFP-Cre was expressed only in the perichondrium of the femur and tibia at E15.5 (Figure 5). Therefore, deletion of Runx2 in the perichondrium should decelerate the terminal differentiation of chondrocytes in Runx2^fl/−Cre^ mice at E15.5. Thus, genes responsible for deceleration should be expressed in the perichondrium and encode the secreted proteins. Nell1 is a secreted protein that has been reported to be expressed in the perichondrium and chondrocytes and to induce chondrocyte maturation [15,16,18]. Moreover, Nell1 expression has been reported to be regulated by Runx2 [16,19,20].
2.6. Nell1 Was Downregulated in the Perichondrium of Runx2fl/−Cre Mice, Overexpression of Runx2 Induced Nell1 Expression, and Runx2 siRNA Reduced Nell1 Expression
Real-time RT-PCR using RNA from femurs and tibiae at E15.5 showed that the expression of Nell1 in Runx2^fl/−Cre^ mice was significantly lower than that in Runx2^fl/+^ mice, although the reduction was not significant compared to that in Runx2^fl/−^ mice (Figure 8A). Although Fgf18 in the perichondrium had been shown to regulate chondrocyte maturation [9], Fgf18 expression was comparable among Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice (Figure 8B). The expression of Nell1 was also examined by immunohistochemistry using an Nell1 antibody. Nell1 was detected similarly in chondrocytes among Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice, but Nell1 expression in the perichondrium in Runx2^fl/−Cre^ mice was reduced compared to that in Runx2^fl/+^ and Runx2^fl/−^ mice (Figure 8C–K). In accordance with previous reports [16,19,20], Runx2 overexpression in primary osteoblasts induced Nell1 expression (Figure 8L,M). Moreover, Runx2 siRNA reduced Nell1 expression in primary osteoblasts (Figure 8N).
2.7. Both Trabecular and Cortical Bone Were Severely Reduced in Runx2fl/−Cre Femurs at 11 Weeks of Age
Micro-CT analysis showed that the trabecular parameters, including bone volume/total volume (BV/TV), trabecular thickness (Tb.Th), trabecular number (Tb.N), and trabecular bone mineral density (Tb.BMD), and the cortical parameters, including cortical area/total area (CtAr/TtAr), cortical thickness (Ct.Th), and cortical BMD (Ct.BMD), were severely reduced in Runx2^fl/−Cre^ femurs compared to those in Runx2^fl/+^ femurs (Figure 9A–F). The reduction compared to that in wild-type mice was more severe in Runx2^fl/−Cre^ mice than in Runx2^+/−^ mice [22]. Histological analysis also showed a reduction in trabecular and cortical bones in Runx2^fl/−Cre^ mice compared to that in Runx2^fl/+^ mice (Figure 9G–L). These findings indicate the requirement of Runx2 in committed osteoblasts in adult mice.
3. Discussion
Deletion of Runx2 by Col1a1 Cre resulted in retarded terminal differentiation of the chondrocytes. Nell1 expression was reduced in the perichondrium of Runx2^fl/−Cre^ mice, and overexpression and knockdown of Runx2 induced and reduced Nell1 expression, respectively. These findings, combined with previous reports, indicate that Runx2 in the perichondrium positively regulates the terminal differentiation of chondrocytes by inducing Nell1 expression.
Removal of the perichondrium from chick tibiotarsi elongates the tibiotarsi and increases the layer of proliferating chondrocytes, followed by an increase in the Col10a1 expressing region during organ culture, suggesting that the perichondrium inhibits chondrocyte proliferation and maturation [8]. However, the growth of perichondrium-removed chick tibiotarsi was similar to that of perichondrium-intact tibiotarsi when cultured in the chorioallantoic membrane [23]. Thus, the function of the perichondrium against chondrocytes remains controversial. In contrast to our findings, Runx2 in the perichondrium of long bones was shown to negatively regulate chondrocyte proliferation and maturation by inducing Fgf18, which is expressed in the perichondrium [9]. Twist1, which is expressed in the perichondrium, inhibits Runx2 function, and Twist1 mutant mice were shown to inhibit chondrocyte maturation, which was interpreted in terms of the shortening of the Col10a1-expressing layer [9]. However, the phenotypes of Twist1 mutant mice showed that the reduced Col10a1 layer was caused by enhanced chondrocyte maturation and endochondral ossification. Thus, the controversy was caused by the misinterpretation of the phenotypes of Twist1 mutant mice, and the correct interpretation of the phenotype indicates that Runx2 in the perichondrium enhances chondrocyte maturation. Furthermore, Fgf18 enhances the terminal differentiation of chondrocytes [10,11]. Thus, it is still possible that Runx2 in the perichondrium enhances chondrocyte maturation through Fgf18 induction. However, Fgf18 expression was similar in Runx2^fl/+^, Runx2^fl/−^, and Runx2^fl/−Cre^ mice.
Nell1 and Runx2 have similar expression patterns in the calvaria and long bones, and Nell1 expression is severely reduced in Runx2^−/−^ mice [15,16,19]. In long bones, both are strongly expressed in prehypertrophic, hypertrophic, and terminal hypertrophic chondrocytes, perichondrium, and osteoblasts in the bone marrow and bone collar [15,16]. Both Runx2 and Nell1 enhance chondrocyte maturation and osteoblast differentiation, and Nell1 expression is directly regulated by Runx2 [3,13,16,18,19,20]. We also confirmed that Nell1 expression is regulated by Runx2 (Figure 8L–N). Col10a1-expressing hypertrophic chondrocytes were reduced in chondrocyte-specific Nell1-deficient mice [18], whereas maturation to Col10a1-positive hypertrophic chondrocytes was not impaired in Runx2^fl/−Cre^ mice. Therefore, Nell1 in chondrocytes may be sufficient for maturation into Col10a1 expressing hypertrophic chondrocytes. As the differentiation of Col10a1-expressing hypertrophic chondrocytes to Spp1 expressing terminal hypertrophic chondrocytes was retarded in Runx2^fl/−Cre^ mice, Nell1 in the perichondrium, in addition to that in chondrocytes, is likely to be necessary for normal terminal differentiation.
4. Materials and Methods
4.1. Mice
Runx2 flox mice were generated by homologous recombination using a targeting vector containing Loxps upstream and downstream of exon 2, as previously described [2]. Runx2^+/−^ mice were generated as previously described [22], and 2.3 kb Col1a1 EGFP-Cre transgenic mice, which produce EGFP-Cre fusion protein specifically in osteoblasts, were generated using the 2.3 kb Col1a1 promoter and EGFP-Cre DNA, as previously described [21]. Runx2^fl/−^ and Runx2^fl/−Cre^ mice were generated by crossing Runx2^fl/+Cre^ with Runx2^+/−^ mice. Runx2^fl/−Cre^ mice express Runx2 in one allele and in chondrocytes but not in osteoblasts. Runx2^fl/+^ mice were generated and maintained on a C57BL/6N background; 2.3-kb Col1a1 EGFP-Cre transgenic mice were generated on a B6C3H F1 background and backcrossed with C57BL/6N mice more than eight times; and Runx2^+/^^−^ mice were generated on a 129Ola/C57BL6 background and backcrossed with C57BL/6N at least 12 times. Before the study, all experimental protocols were reviewed and approved by the Animal Care and Use Committee of the Nagasaki University Graduate School of Biomedical Sciences (No. 2401311920-4). Animals were housed three per cage in a pathogen-free environment on a 12 h light cycle at 22 ± 2 °C, with standard chow (CLEA Japan, Tokyo, Japan) and free access to tap water. All relevant guidelines for working with animals were followed.
4.2. Skeletal Staining and Micro-CT Analysis
The vertebral, thoracic cage, and upper limb bones were stained with alcian blue and alizarin red, as described previously [22]. Micro-CT analysis was performed using a micro-CT system (R_mCT; Rigaku Corporation, Tokyo, Japan). Data from the scanned slices were used for three-dimensional analysis to calculate the femoral morphometric parameters. Trabecular bone parameters were measured in the distal femoral metaphysis of the femur. Craniocaudal scans of approximately 2.4 mm (0.5 mm from the growth plate) for 200 slices in 12 μm increments were obtained. The cortical bone parameters were measured at the mid-diaphysis of the femurs. The threshold for the mineral density was set at 500 mg/cm^3^.
4.3. Histological Analyses
Mice were fixed in 4% paraformaldehyde/0.1 M phosphate buffer, E15.5 embryos were undecalcified, and other stages of mice were decalcified and embedded in paraffin. Sections (4 μm thick) were stained with hematoxylin and eosin (H&E), von Kossa, or tartrate-resistant acid phosphatase (TRAP). In situ hybridization was conducted using mouse Col2a1, Col10a1, Spp1, and Col1a1 antisense and sense probes, as described previously [2]. The sections were then counterstained with methyl green. To observe osteoblast differentiation in the bone collars of femurs, Runx2^fl/+Cre^ and Runx2^fl/−Cre^ embryos at E15.5 were fixed with 4% paraformaldehyde at 4 °C for 2 h, washed with PBS at 4 °C for 1 h, immersed in 20% sucrose at 4 °C overnight, and embedded in O. C. T. compound (Sakura Finetek, Tokyo, Japan), frozen in a refrigerated installation (Rikakikai UT-2000F, Tokyo, Japan) containing −100 °C hexane and pentane (10:3), and sectioned at 7 μm thickness using a Leica CM3050S (Leica Biosystems, Tokyo, Japan). The same section was first subjected to fluorescence imaging, followed by H&E staining. Immunohistochemistry was performed using a polyclonal sheep anti-Nell1 antibody (R&D Systems, Catalog #AF7109, Minneapolis, MN, USA) and a polyclonal donkey anti-sheep IgG horseradish peroxidase-conjugated antibody (R&D Systems, Catalog #HAF016).
4.4. Real-Time Reverse Transcription (RT)-PCR
Total RNA was extracted using ISOGEN (Wako, Osaka, Japan). For cDNA synthesis, 500 ng of total RNA was reverse-transcribed using ReverTra Ace qPCR Master with gDNA Remover (Toyobo, Osaka, Japan). Real-time RT-PCR was performed using the THUNDERBIRD SYBR qPCR Mix (Toyobo) and a Light Cycler 480 real-time PCR system (Roche Diagnostics, Tokyo, Japan). The values were normalized to that of Actb. Primer sequences are shown in Supplementary Table S1. The expression of Col10a1 was examined using TaqMan probes Mm00487041_m1 for Col10a1 and Mm02619580_g1 for Actb (Thermo Fisher Scientific, Tokyo, Japan).
4.5. RNA Sequencing Analysis
Total RNA was prepared from the femurs and tibiae, in which skin and muscle had been removed, of three Runx2^fl/−^ and Runx2^fl/−Cre^ embryos at E15.5 using ISOGEN (Wako). RNA sequencing was performed on the DNBSEQ platform (MGI Tech, Shenzhen, China) by Tokai Biotechnology Co., Ltd. (Osaka, Japan). Sequence quality was first surveyed with FastQC (version 0.12.1). Paired-end raw reads were trimmed using Trimmomatic and mapped to the mouse genome assembly mm10. The GRCm38 reference genome was obtained from the UCSC Genome Browser database using Bowtie2 (version 2.7.11b). mRNA expression was quantified using featureCounts (version 2.0.3). Differentially expressed mRNA was calculated using the DESeq2 R package (version 1.38.3) with a cutoff fold change >2 and FDR-adjusted p-value (p-value) < 0.05. The GO enrichment analysis was performed using the function “enrichGO” from the R package clusterProfiler (version 4.6.2).
4.6. Cell Culture and Runx2 Overexpression and Knockdown
Primary osteoblasts were isolated from the calvariae of newborn wild-type mice by sequential digestion with 0.1% collagenase A and 0.2% dispase (Sigma, St. Louis, MO, USA) for 10 min at 37 °C. Osteoblastic cells from the third to fifth fractions were pooled and plated on 48-well plates at a density of 4.0 × 10^4^ cells/cm^2^ in alpha-modified Eagle’s Minimum Essential Medium (αMEM; Merck, Darmstadt, Germany) supplemented with 10% fetal bovine serum (Nichirei, Tokyo, Japan), L-glutamine (Merck), and 100 U/mL penicillin (Nacalai Tesque, Inc., Kyoto, Japan) and 100 μg/mL streptomycin (Nacalai Tesque). Cells were transfected with 0.1 μg of control (pSG5-Mock) or Runx2-expressing (pSG5-Runx2) vectors using X-tremeGENE9 (Roche Diagnostics). Cells were also transfected with 90 pmol Runx2 siRNA (triplex; Thermo Scientific) or scrambled RNA using X-tremeGENE360 (Roche Diagnostics). The transfected cells were cultured for 24 h or 48 h.
4.7. Statistical Analysis
Values are expressed as the mean ± SD. Statistical analyses of two groups were performed by the Student t-test, and those of more than three groups were conducted by one-way ANOVA by GraphPad Prism (8.0.1). Statistical significance was set at p < 0.05.
4.8. Graphical Abstract
The graphical abstract was drawn using FigDraw 2.0.
5. Conclusions
In conclusion, contrary to a previous report [9], Runx2 in the perichondrium positively regulates chondrocyte terminal differentiation. Therefore, in endochondral ossification, Runx2 induces chondrocyte maturation directly and indirectly through the perichondrium by inducing Nell1 expression. This report added a novel function of Runx2 in endochondral ossification.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Karaplis A.C. Embryonic development of bone and the molecular regulation of intramembranous and endochondral bone formation Principles of Bone Biology Academic Press London, UK 2002 Volume 13358
- 2Qin X. Jiang Q. Nagano K. Moriishi T. Miyazaki T. Komori H. Ito K. Mark K.V. Sakane C. Kaneko H. Runx 2 is essential for the transdifferentiation of chondrocytes into osteoblasts P Lo S Genet.202016 e 100916910.1371/journal.pgen.100916933253203 PMC 7728394 · doi ↗ · pubmed ↗
- 3Komori T. Regulation of Skeletal Development and Maintenance by Runx 2 and Sp 7Int. J. Mol. Sci.2024251010210.3390/ijms 25181010239337587 PMC 11432631 · doi ↗ · pubmed ↗
- 4Vortkamp A. Lee K. Lanske B. Segre G.V. Kronenberg H.M. Tabin C.J. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein Science 199627361362210.1126/science.273.5275.6138662546 · doi ↗ · pubmed ↗
- 5Chung U.I. Schipani E. Mc Mahon A.P. Kronenberg H.M. Indian hedgehog couples chondrogenesis to osteogenesis in endochondral bone development J. Clin. Investig.200110729530410.1172/JCI 1170611160153 PMC 199199 · doi ↗ · pubmed ↗
- 6Ohba S. Hedgehog Signaling in Skeletal Development: Roles of Indian Hedgehog and the Mode of Its Action Int. J. Mol. Sci.202021666510.3390/ijms 2118666532933018 PMC 7555016 · doi ↗ · pubmed ↗
- 7St-Jacques B. Hammerschmidt M. Mc Mahon A.P. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation Genes Dev.1999132072208610.1101/gad.13.16.207210465785 PMC 316949 · doi ↗ · pubmed ↗
- 8Long F. Linsenmayer T.F. Regulation of growth region cartilage proliferation and differentiation by perichondrium Development 19981251067107310.1242/dev.125.6.10679463353 · doi ↗ · pubmed ↗