Functional Divergence of adcyap1b Splice Variants in Regulating Pituitary Hormone Expression in the Chinese Tongue Sole (Cynoglossus semilaevis)
Qian Zhang, Xihong Li, Yue Zhang, Wenjie Li, Zhenyu Cai, Wenteng Xu, Songlin Chen, Na Wang

TL;DR
This study explores how different versions of a gene in a type of fish affect pituitary hormone production, which may influence sexual size differences.
Contribution
The study identifies and functionally characterizes two splice variants of adcyap1b in Chinese tongue sole, revealing their divergent roles in regulating pituitary hormones.
Findings
Two adcyap1b splice variants were identified, differing by the inclusion or skipping of exon 4.
Both variants increased gh, pomc, and crh expression, but ADCYAP1b_tv1 uniquely stimulated prl and sl.
ADCYAP1b_tv1 showed stronger and broader effects on pituitary hormone expression compared to ADCYAP1b_tv2.
Abstract
Sexual size dimorphism (SSD) refers to the phenomenon where males and females of the same species exhibit differences in overall or partial body size, and it is widespread among mammals, birds, reptiles, and fish. Notably, this dimorphism is significantly influenced by the sexually dimorphic secretion of growth hormone (gh), a key pituitary-derived growth regulator. Commonly, the secretion of gh is positively regulated by glucagon family members such as growth hormone-releasing hormone (ghrh) and adenylate cyclase-activating polypeptide 1 (adcyap1). To explore the stimulators for pituitary hormones (especially gh) in the teleost, we performed genome-wide identification and functional characterization of the glucagon family on Chinese tongue sole (Cynoglossus semilaevis) that exhibits typical female-biased sexual size dimorphism. Four members of adcyap1/vasoactive intestinal…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
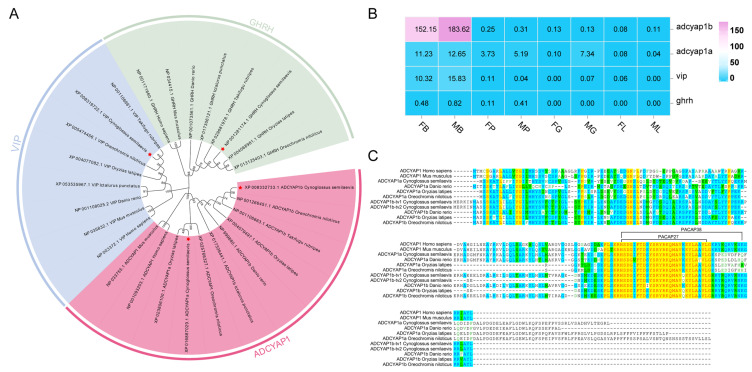 Figure 1
Figure 1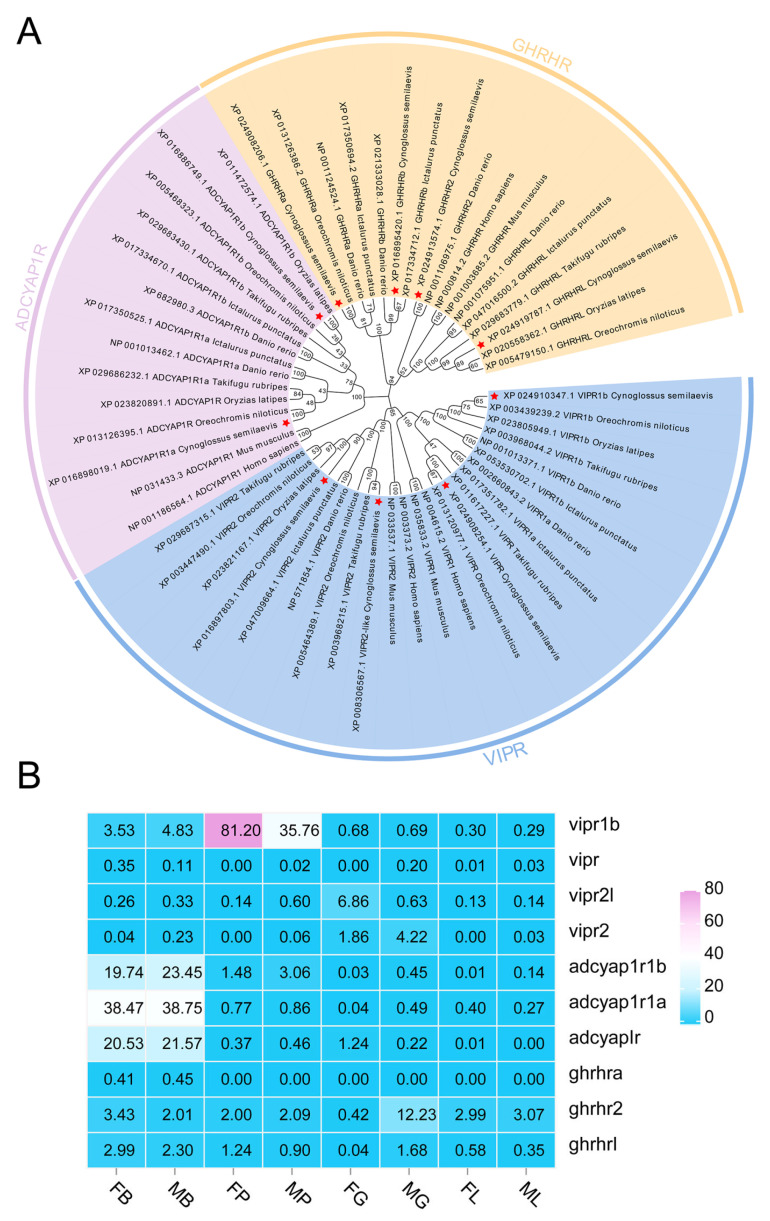 Figure 2
Figure 2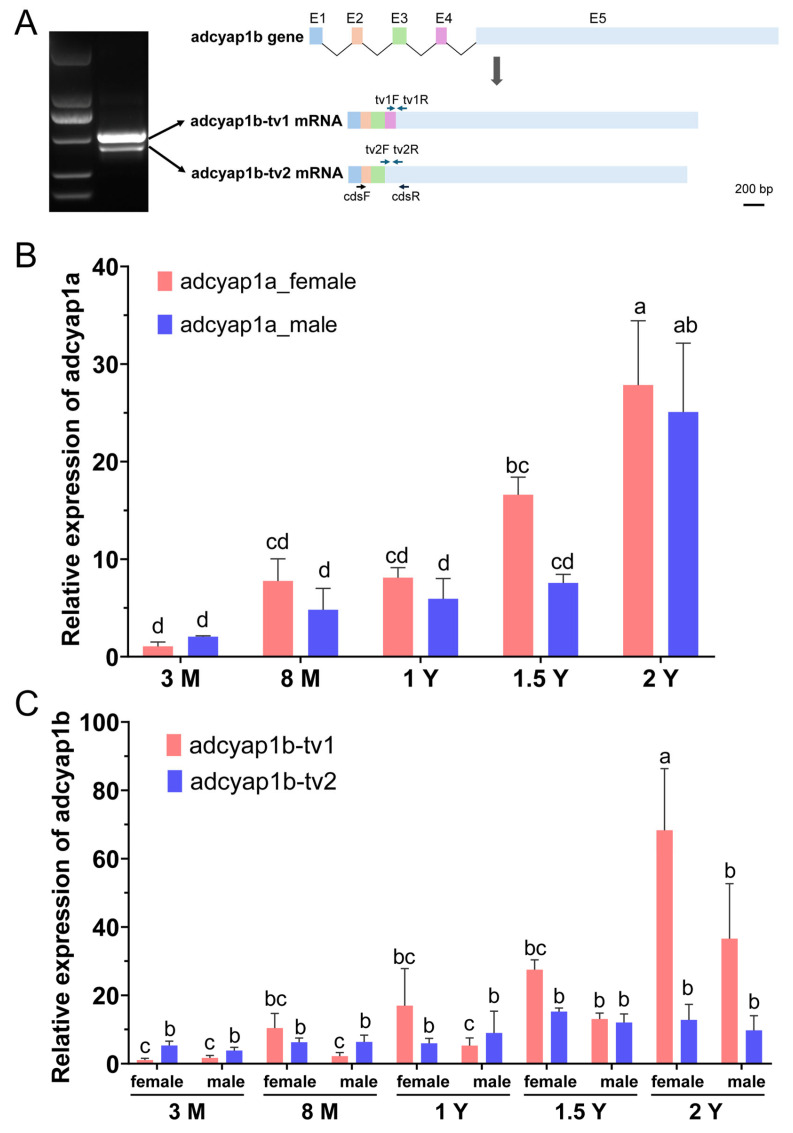 Figure 3
Figure 3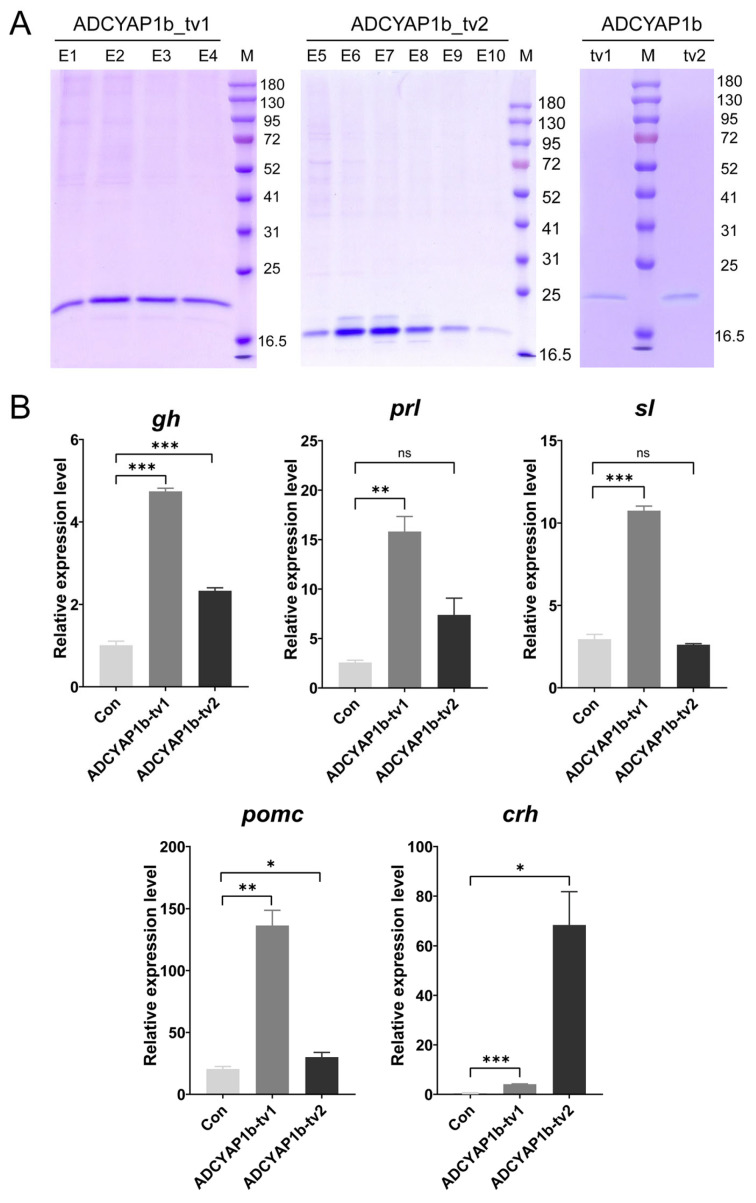 Figure 4
Figure 4- —National Natural Science Foundation of China
- —Taishan Young Scholar Project of Shandong Province
- —Central Public-interest Scientific Institution Basal Research Fund, CAFS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGrowth Hormone and Insulin-like Growth Factors · Regulation of Appetite and Obesity · Hypothalamic control of reproductive hormones
1. Introduction
Sexual size dimorphism, the difference in body size between sexes, occurs across various species including mammals, birds, reptiles, and fish [1,2]. The secretion of growth hormone (gh) from the pituitary is a pivotal regulator of organismal growth [3] and itself frequently displays sexual dimorphism. Sexually dimorphic expression of gh gene drives sex-specific differences in growth, development, and physiological functions, thereby contributing to sexual growth dimorphism [4].
Understanding the regulation of this differential gh secretion is essential. The conventional view holds that in mammals and birds, it is primarily under the positive control of growth hormone-releasing hormone (GHRH), a glucagon family member [5,6]. However, in fish models such as goldfish (Carassius auratus) and medaka (Oryzias latipes), the positive regulation of gh is primarily mediated by pituitary adenylate cyclase-activating polypeptide (PACAP) encoded by the adcyap1 gene—another member of the glucagon family [7,8,9]. PACAP was first isolated from the cattle hypothalamus [10,11] and influenced the secretion of multiple hormones within both the hypothalamic–pituitary–gonadal (HPG) and hypothalamic–pituitary–adrenal (HPA) axes by activating the ERK and cAMP/PKA signaling pathways [12,13,14].
The Chinese tongue sole (Cynoglossus semilaevis) is a flatfish endemic to China and is known for its pronounced female-biased growth dimorphism [15]. Although previous studies have begun to elucidate the mechanisms behind this phenomenon [16,17,18,19], the sexual dimorphic regulation of gh secretion remains a key unresolved question. Recently, using single-cell multi-omics analysis, we found that adcyap1-positive GABAergic neurons are 2.32 times more abundant in females than in males (unpublished data). It remains to be investigated whether adcyap1 serves as a central regulatory function in pituitary hormone secretion, rather than ghrh.
To address this issue, this study firstly conducted a genome-wide identification and evolutionary analysis of the glucagon family genes (ghrh/adcyap1/vip) and their receptors in the Chinese tongue sole. Furthermore, we examined the expression patterns of ligands and their receptors in key growth-axis tissues across different sexes. Finally, the functional role of ADCYAP1b in regulating pituitary hormone secretion was elucidated through in vitro incubation assays. This work provides a crucial foundation for understanding the regulatory mechanisms behind pituitary hormone secretion.
2. Results
2.1. Genome-Wide Identification and Evolutionary Analysis of Members of the Glucagon Family and Its Receptors in C. semilaevis
By genome-wide screen, four glucagon family members were identified from C. semilaevis (Table 1). A phylogenetic tree of glucagon family genes was constructed by aligning homologous protein sequences in C. semilaevis, Oryzias latipes, Oreochromis niloticus, Takifugu rubripes, Danio rerio, Ictalurus punctatus, Homo sapiens, and Mus musculus. The results (Figure 1A) show that the sequences are primarily clustered into the following three major branches: ADCYAP1, VIP, and GHRH. Furthermore, ADCYAP1 branch is subdivided into the ADCYAP1B and ADCYAP1A subclades. C. semilaevis ADCYAP1A and ADCYAP1B are both closely related with ADCYAP1B of Oreochromis niloticus and Oryzias latipes. In VIP branch, C. semilaevis vip clusters closely with the VIP sequences of T. rubripes and O. latipes. In GHRH branch, C. semilaevis GHRH clusters closely with the GHRH sequences of O. latipes and O. niloticus. In addition, the evolutionary tree clearly illustrates the phylogenetic patterns of the ADCYAP1, VIP, and GHRH families in teleost fishes and mammals.
By using our previous data of two-year-old C. semilaevis tissues RNA-seq, adcyap1b showed the highest expression level in the brains, compared with the other members of glucagon family members (Figure 1B). Noticeably, the expression of ghrh was barely detectable in the brain. By comparing the ADCYAP1 protein sequences of teleosts and mammals, it was found that most proteins have conserved sequences near the C-terminus. Among them, ADCYAP1b is more like mammalian PACAP38, while PACAP27 shows higher similarity between fish ADCYAP1a and mammalian ADCYAP1 proteins (Figure 1C).
To further explore ligand–receptor interaction mechanisms, the genome-wide identification for the glucagon family receptors was also conducted. As a result, ten members were screened from C. semilaevis and aligned with other teleosts including O. latipes, O. niloticus, T. rubripes, D. rerio, I. punctatus, as well as H. sapiens and M. musculus. The phylogenetic tree (Figure 2A) reveals that the sequences are clustered into the following three major branches: VIPR branch, ADCYAP1R branch, and GHRHR branch. For mammals, VIPR branch contained VIPR1 and VIPR2. In teleosts, four members including VIPR1a, VIPR1b, VIPR2, and VIPR2l are included. Similarly, teleost ADCYAP1R branch includes ADCYAP1R1a and ADCYAP1R1b subtypes, different from one member in mammals. For GHRHR branch, still one member is identified from mammals, while four members including GHRHRa, GHRHRb, GHRHR2, and GHRHRl are screened from the teleost species. The RNA-seq data of two-year-old C. semilaevis tissues (Figure 2B) revealed that three receptors of adcyap1r showed a relatively high expression level in the brains. In the pituitary gland, vipr1b exhibited the highest abundance.
2.2. Expression Patterns and Alternative Splicing Analysis of the adcyap1 Gene in C. semilaevis
During the verification process of the adcyap1b gene sequence, two specific PCR bands were amplified by primers cdsF and cdsR (Figure 3A). Sequencing analysis revealed that the sizes of these two bands were 546 bp and 441 bp, respectively, with the difference primarily resulting from exon 4 skipping. This indicates that the adcyap1b gene of C. semilaevis undergoes alternative splicing, and the gene can be transcribed into two alternative splice variants, adcyap1b_tv1 and adcyap1b_tv2.
The relative expression levels of the genes were calculated using the 2^−∆∆Ct^ method. The results showed that the expression of the adcyap1a gene exhibited a gradual upregulation trend with increasing age, reaching a common peak in both female and male individuals at 2 years of age (2 Y). The adcyap1b gene displayed differential expression through its distinct transcript variants (adcyap1b_tv1 and adcyap1b_tv2), among which the expression dynamics of adcyap1b_tv1 were more specific.
2.3. Recombinant Eukaryotic Expression and Functional Analysis of Two Splice Variants of ADCYAP1b
Two splice variants of ADCYAP1b (ADCYAP1b_tv1 and ADCYAP1b_tv2) were successfully expressed in a recombinant eukaryotic system, exhibiting the following distinct subcellular localization patterns: ADCYAP1b_tv1 was expressed as an intracellular protein, while ADCYAP1b_tv2 was secreted into the cell culture supernatant. Both proteins were purified by His-tag affinity chromatography and eluted with an imidazole gradient. Approximately 200 µg of each purified protein was obtained, desalted to remove imidazole, and stored in DMEM for subsequent analysis (Figure 4A).
Following in vitro incubation assay using pituitary cells, the quantitative real-time PCR (qRT-PCR) analysis based on the 2^−∆∆Ct^ method was conducted to assess the regulatory effects of ADCYAP1b_tv1 and ADCYAP1b_tv2 proteins on the expression of growth- and endocrine-related genes. Results demonstrated that both variants significantly promoted the transcriptional levels of gh, pro-opiomelanocortin (pomc), and corticoliberin (crh) compared to the control group (p < 0.05), with ADCYAP1b_tv1 exerting a notably stronger promotional effect on gh and pomc compared to ADCYAP1b_tv2. In contrast, a significant promotional effect on the prolactin (prl) and somatolactin (sl) genes was only observed in the recombinant ADCYAP1b_tv1 treatment group.
Combined with the expression patterns of two splice variants, the female-biased expression of adcyap1b_tv1 in the brain is implicated as a critical driver of sexual size dimorphism, likely through stimulating greater gh production in the female pituitary gland.
3. Discussion
Research on the regulation of gh secretion is crucial for understanding sexual size dimorphism (SSD) across species. In mammals, existing evidence indicates that gh secretion is regulated not only by classical ghrh signaling but is also influenced by other factors such as leptin and ghrelin [20,21,22]. In this study, female-biased SSD species C. semilaevis was used to investigate the regulatory mechanisms of gh.
Similar with other teleosts, two adcyap1 genes including adcyap1a and adcyap1b were identified from C. semilaevis. By comparison with the ADCYAP1 protein, teleost ADCYAP1b exhibited more similarity with mammalian PACAP38. It implied that adcyap1a gene may have originated from a fish-specific genome duplication event [23]. The adcyap1 gene is widely expressed in mammals, with a distribution that spans the central and peripheral nervous systems as well as non-neural tissues ranging from the adrenal glands and gonads to the pancreas (reviewed by [24]). In C. semilaevis, adcyap1b shows high expression exclusively in brain tissues, while adcyap1a exhibits moderate expression in brains, pituitary glands, and male gonad. These differences in mRNA localization between the two adcyap1 paralogs suggest their functions might have diverged concomitant with the occurrence of genome duplication.
Moreover, two alternative splicing transcripts of C. semilaevis adcyap1b, resulting from exon 4 skipping, were identified. In mammals, the adcyap1 gene employs multiple promoters to generate tissue-specific transcripts for unique physiological functions [25,26]. Illustratively, it has been demonstrated that in rats, a specific adcyap1 promoter expresses a distinct mRNA in the testis, which is crucial for maintaining the process of spermatogenesis [25,27].
Evidence from fish species including goldfish and medaka has suggested that adcyap1 may function as a hypophysiotropic factor regulating pituitary hormone secretion [7,8,9]. In the present study, two splice proteins of ADCYAP1b in C. semilaevis both promote the secretion of gh, pomc, and crh, with ADCYAP1b_tv1 exhibiting a stronger regulatory effect. It implied that adcyap1b may play crucial roles in growth and development, feeding behavior, and sex reversal [28,29,30,31]. Moreover, ADCYAP1b_tv1 can also regulate osmoregulation and reproductive behavior by promoting the secretion of prl [32,33] and sl [34,35].
In addition, the effects of ADCYAP1 on pituitary hormones appear to be species-specific, as evidenced by comparison across different fish species. For instance, medaka ADCYAP1 recombinant protein does not significantly promote gh but significantly stimulates both prl, sl, and luteinizing hormone subunit beta (lhb) in both females and males [9]. Additionally, it exhibits an inhibitory effect on male pomc. In another fish-goldfish, ADCYAP1 analogs could stimulate the release of pituitary hormones, including gh and lhb [7].
Overall, the study establishes ADCYAP1b, particularly its tv1 variant, as a potent and multifunctional pituitary hormone regulator in C. semilaevis.
4. Materials and Methods
4.1. Ethics Statement
All fish were euthanized by submerging in MS-222 (120 mg/L) solution prior to the experiments. The sampling and treatment of C. semilaevis was approved by Institute Animal Care and Use Committee of the Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences (approval number YSFRI-2025017).
4.2. Fish Sampling and Cell Culture
All healthy C. semilaevis at 3 months, 8 months, 1 year, 1.5 years, and 2 years of age were sourced from Huanghai Aquaculture Co., Ltd. (Haiyang, China). The fish were reared in a recirculating aquaculture system, fed a small amount of diet twice daily (at 8:00 a.m. and 4:00 p.m.), and maintained at a water temperature of 18–22 °C.
C. semilaevis pituitary cells were cultured in L-15 medium supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY, USA) and maintained in an incubator at 24 °C.
4.3. Genome-Wide Identification and Evolutionary Analysis of the Glucagon Family and Its Receptors in C. semilaevis
The protein sequences of adcyap1, vip, and ghrh from C. semilaevis were retrieved from the NCBI database for phylogenetic analysis. Corresponding gene sequences from other teleosts and mammals, including Oryzias latipes, Takifugu rubripes, Danio rerio, and Homo sapiens, were also selected. Sequence alignment was performed using ClustalW in MEGA (Version 11.0.13), and a phylogenetic tree was constructed using the neighbor-joining method. The resulting tree was subsequently visualized and refined using the EvolView webserver (http://www.evolgenius.info/evolview/#/treeview (accessed on 1 September 2025)).
We utilized RNA-seq data of the brain, liver, gonads, and pituitary from 2-year-old fish [16] to profile the mRNA distribution in C. semilaevis. The relative mRNA abundances of the above-mentioned adcyap1/vip/ghrh ligand and receptor family member genes were obtained from these data, and a heatmap was created using the Omicshare Tool (https://www.omicshare.com/tools (accessed on 1 September 2025) to visualize the expression profiles. The relevant ADCYAP1 protein sequences from various species were aligned with Vector NTI software (Version 11.5.1) to generate a protein sequence alignment figure.
4.4. Expression Profiling and Alternative Splicing Analysis of the adcyap1 Gene in C. semilaevis
To examine the expression patterns of the adcyap1b gene across developmental stages, PCR amplification was initially conducted using cDNA from the brain tissue of 1.5-year-old C. semilaevis. The reaction was performed in a 50 μL system containing 25 μL KOD Mix, 1.5 μL each of coding region-specific primers (cdsF and cdsR) (Table 2), 20 μL ddH_2_O, and 2 μL cDNA template. The PCR protocol consisted of an initial denaturation at 98 °C for 10 s, followed by 40 cycles of denaturation at 98 °C for 10 s, annealing at 58–62 °C for 5 s, and extension at 68 °C for 25 s, with a final extension at 72 °C for 7 min. This process led to the identification of two alternative splice variants of adcyap1b, designated adcyap1b_tv1 and adcyap1b_tv2.
Two pairs of quantitative PCR (qPCR) primers (tv1F/tv1R and tv2F/tv2R) (Table 2) were designed to specifically target each splice variant. qPCR was performed using cDNA derived from brain tissues of four female and four male C. semilaevis at different developmental stages. Each 10 µL reaction contained 5 µL SYBR Taq, 0.2 µL each of forward and reverse primers, 1 µL diluted cDNA, and 3.6 µL H_2_O. The thermal cycling conditions were as follows: 95 °C for 30 s; 40 cycles of 95 °C for 3 s; and 60 °C for 33 s, followed by a melting curve analysis (95 °C for 15 s, 60 °C for 60 s, and 95 °C for 15 s). Relative expression levels were calculated using the 2^−∆∆Ct^ method, analyzed by LSD multiple comparison tests, and visualized using GraphPad Prism 8.0.2.
4.5. Recombinant Eukaryotic Expression of Two ADCYAP1b Splice Variants
To functionally characterize adcyap1b in C. semilaevis, we performed recombinant eukaryotic expression and purification of its two splice variants using the HEK 293T mammalian cell line. The coding sequences of ADCYAP1b_tv1 and ADCYAP1b_tv2 were codon-optimized, and an exogenous signal peptide (MYRMQLLSCIALSLALVTNS) was added to the N-terminus, followed by a C-terminal Flag-His purification tag. The modified fragments were cloned into the pcDNA3.1(–) vector via EcoRI and HindIII restriction sites, resulting in the recombinant constructs pcDNA3.1-ADCYAP1b_tv1 and pcDNA3.1-ADCYAP1b_tv2.
These plasmids were separately transfected into HEK 293T cells using a liposome-based reagent. At 48–72 h post-transfection, cell culture supernatants and lysates were collected and analyzed by Western blot with an anti-Flag antibody to confirm recombinant protein expression. Recombinant ADCYAP1b_tv1 (intracellular) and ADCYAP1b_tv2 (secreted) were successfully purified by His-tag affinity chromatography. The purified proteins were then desalted to remove imidazole and stored in DMEM medium for subsequent functional assays.
4.6. Effect of ADCYAP1b Recombinant Proteins on the Pituitary Cells
To investigate the regulatory effects of the two recombinant adcyap1b splice variants (ADCYAP1b_tv1 and ADCYAP_tv2) on pituitary hormone secretion in C. semilaevis, we conducted an in vitro incubation assay using pituitary cells. The experiment utilized tenth-passage female pituitary cells cultured in the laboratory. Cells were divided into the following three groups: the ADCYAP1b_tv1 treatment group, the ADCYAP1b_tv2 treatment group, and an untreated control group maintained in normal culture medium. Each group consisted of two replicate flasks (25 cm^2^). For the treatment groups, 0.5 mL of recombinant protein (0.05 μg/μL) was administered to each flask, resulting in a total of 25 μg per flask. Following a 48 h incubation at 24 °C, total RNA was extracted using the Trizol method and reverse-transcribed into cDNA. The expression levels of pituitary genes (gh, prl, sl, pomc, and crh) (Table 2) were quantified by quantitative real-time PCR (qRT-PCR) with β-actin as the reference gene. Relative gene expression was calculated using the 2^−ΔΔCt^ method, and statistical analysis was performed using Student’s t-test in GraphPad Prism 8.0.2.
5. Conclusions
In conclusion, this study provides a comprehensive view of the glucagon family in C. semilaevis, from genome-wide identification to functional characterization. The results demonstrate that C. semilaevis adcyap1b gene produces two functionally distinct ligands via alternative splicing, which contributes to the regulation of pituitary hormone expression. Specifically, both variants promoted the expression of gh, pomc, and crh, but ADCYAP1b_tv1 had a significantly stronger effect and uniquely stimulated prl and sl. Nevertheless, the detailed molecular pathways and specific cell groups through which adcyap1b acts require further clarification. Targeted investigations combining single-cell multi-omics and molecular experiments are warranted to address these questions, thereby elucidating the detailed mechanism on sexual size dimorphism.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Janicke T. Fromonteil S. Sexual selection and sexual size dimorphism in animals Biol. Lett.2021172021025110.1098/rsbl.2021.025134520680 PMC 8440037 · doi ↗ · pubmed ↗
- 2Fairbairn D.J. Blanckenhorn W.U. Székely T. Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism Oxford University Press Oxford, UK 2007
- 3Ahmed S.F. Farquharson C. The effect of GH and IGF 1 on linear growth and skeletal development and their modulation by SOCS proteins J. Endocrinol.201020624925910.1677/JOE-10-004520631046 · doi ↗ · pubmed ↗
- 4Addison M.L. Rissman E.F. Sexual dimorphism of growth hormone in the hypothalamus: Regulation by estradiol Endocrinology 20121531898190710.1210/en.2011-198222315455 PMC 3320260 · doi ↗ · pubmed ↗
- 5Balthasar N. Mery P.F. Magoulas C.B. Mathers K.E. Martin A. Mollard P. Robinson I.C.A.F. Growth hormone-releasing hormone (GHRH) neurons in GHRH-enhanced green fluorescent protein transgenic mice: A ventral hypothalamic network Endocrinology 20031442728274010.1210/en.2003-000612746337 · doi ↗ · pubmed ↗
- 6Kühn E.R. Geelissen S.M. Van der Geyten S. Darras V.M. The release of growth hormone (GH): Relation to the thyrotropic- and corticotropic axis in the chicken Domest. Anim. Endocrinol.200529435110.1016/j.domaniend.2005.02.02215927766 · doi ↗ · pubmed ↗
- 7Wong A.O. Li W.S. Lee E.K. Leung M.Y. Tse L.Y. Chow B.K. Lin H.R. Chang J.P. Pituitary adenylate cyclase activating polypeptide as a novel hypophysiotropic factor in fish Biochem. Cell Biol.20007832934310.1139/o 00-05510949084 · doi ↗ · pubmed ↗
- 8Montero M. Yon L. Kikuyama S. Dufour S. Vaudry H. Molecular evolution of the growth hormone-releasing hormone/pituitary adenylate cyclase-activating polypeptide gene family. Functional implication in the regulation of growth hormone secretion J. Mol. Endocrinol.20002515716810.1677/jme.0.025015711013344 · doi ↗ · pubmed ↗