SARS-CoV-2 Spike Protein Induces Time-Dependent and Brain-Region-Specific Alterations in Ferroptosis Markers: A Preliminary Study in K18-hACE2 Mice
Asmaa Yehia, Chirine Toufaily, Dalia M. Abdel Ghaffar, Gehan El Wakeel, Mohamed Adel, Abeer F. Mostafa, Osama A. Abulseoud

TL;DR
This study explores how the SARS-CoV-2 spike protein affects brain regions in mice, showing time-dependent changes in ferroptosis markers, which could explain some post-COVID symptoms.
Contribution
The study reveals time- and region-specific alterations in ferroptosis markers in mice brains after spike protein exposure, suggesting a potential mechanism for post-COVID neuropsychiatric symptoms.
Findings
Spike protein exposure modulates ferroptosis markers in a brain-region-specific manner.
Ferroptosis-related changes were observed in hippocampus and prefrontal cortex via TEM.
Markers like FPN1, DMT1, and GPx4 showed time-dependent expression patterns across brain regions.
Abstract
Post-COVID syndrome comprises persistent neuropsychiatric manifestations for more than 12 weeks after recovery from acute SARS-CoV-2 infection, yet its underlying pathophysiology is unclear. Ferroptosis, an iron-dependent form of cell death with three hallmarks, iron dysregulation, antioxidant failure, and lipid peroxidation, seems to be involved in COVID-19/post-COVID-19 pathophysiology. Here, we administered the SARS-CoV-2 spike protein S1 subunit intranasally to K18-hACE2 transgenic mice and quantified ferroptotic marker protein expression in four brain regions (hippocampus, prefrontal cortex, cerebellum, and olfactory bulb) at 2, 6, and 12 weeks post-administration, alongside ultrastructural assessment by transmission electron microscopy (TEM) that was limited to the hippocampus and prefrontal cortex. Two-way ANOVA revealed region- and time-dependent modulation of iron-handling,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
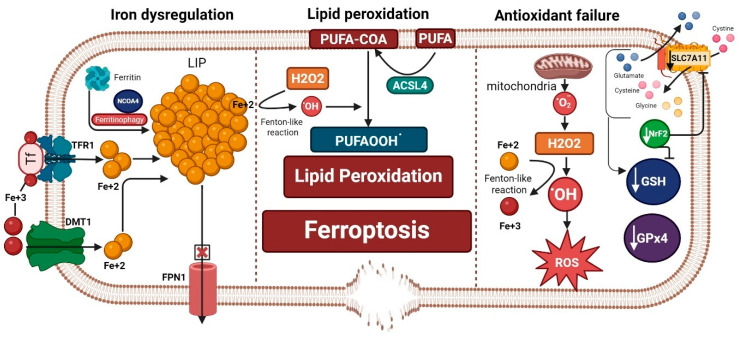 Figure 1
Figure 1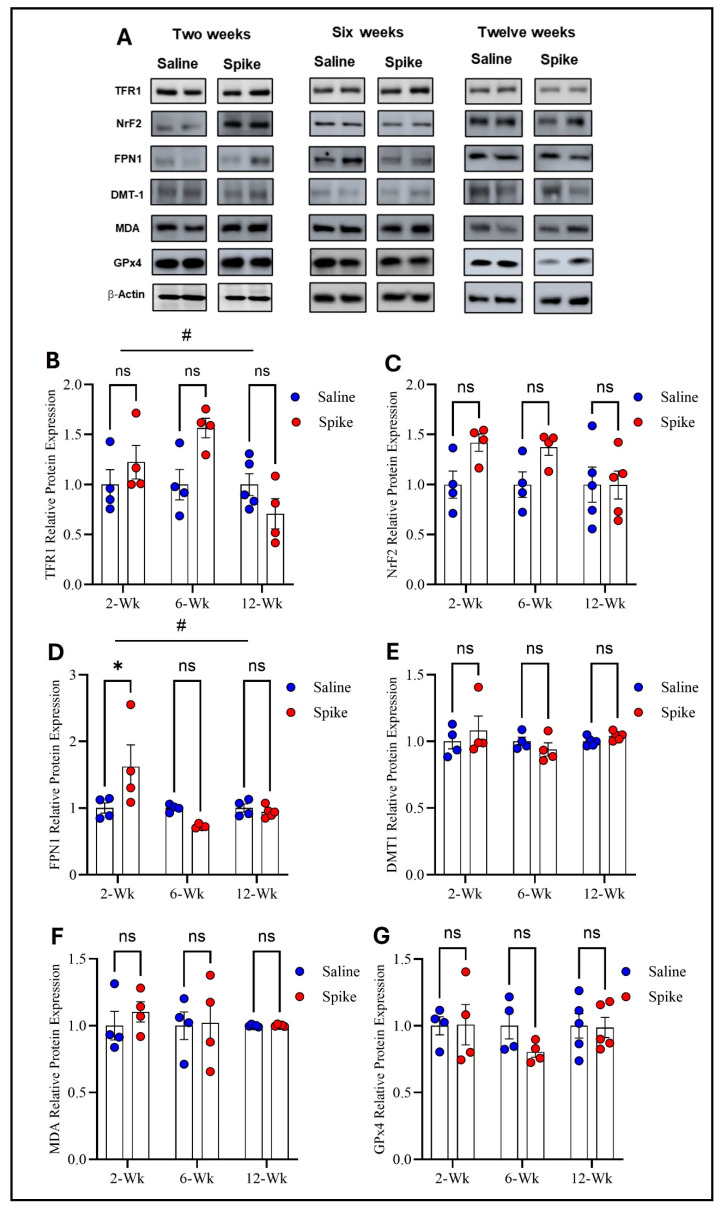 Figure 2
Figure 2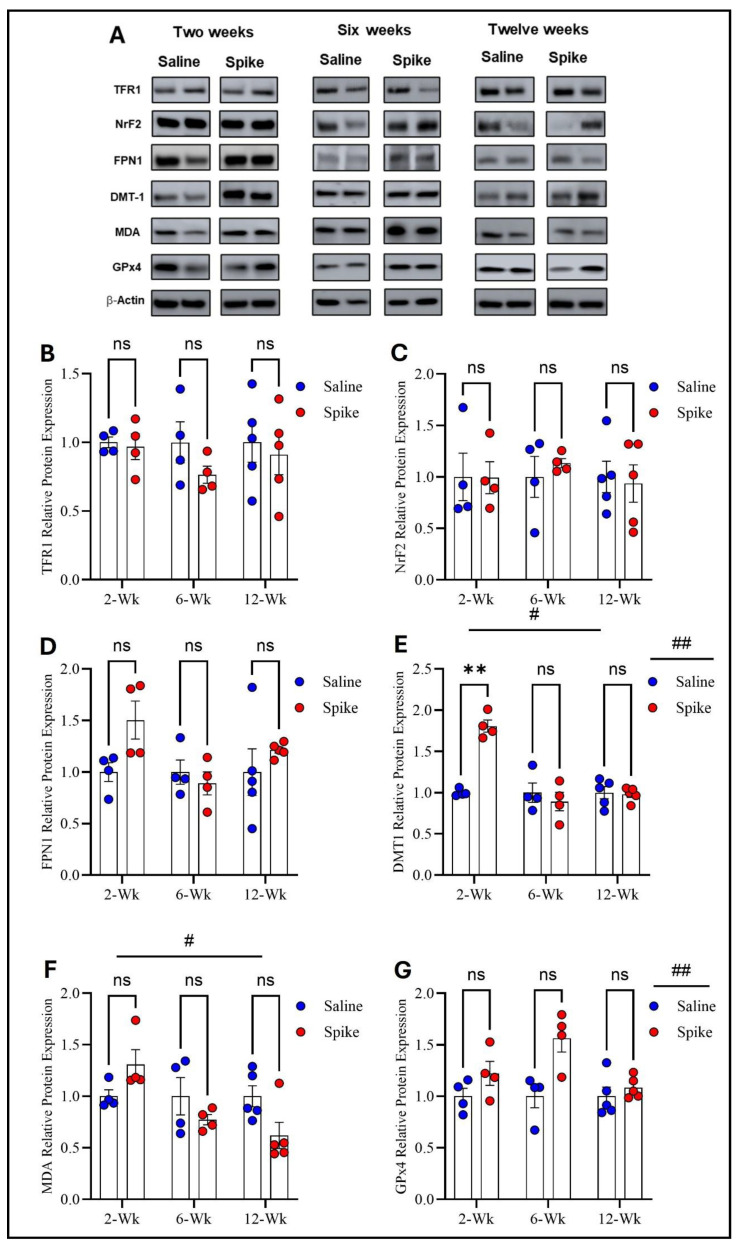 Figure 3
Figure 3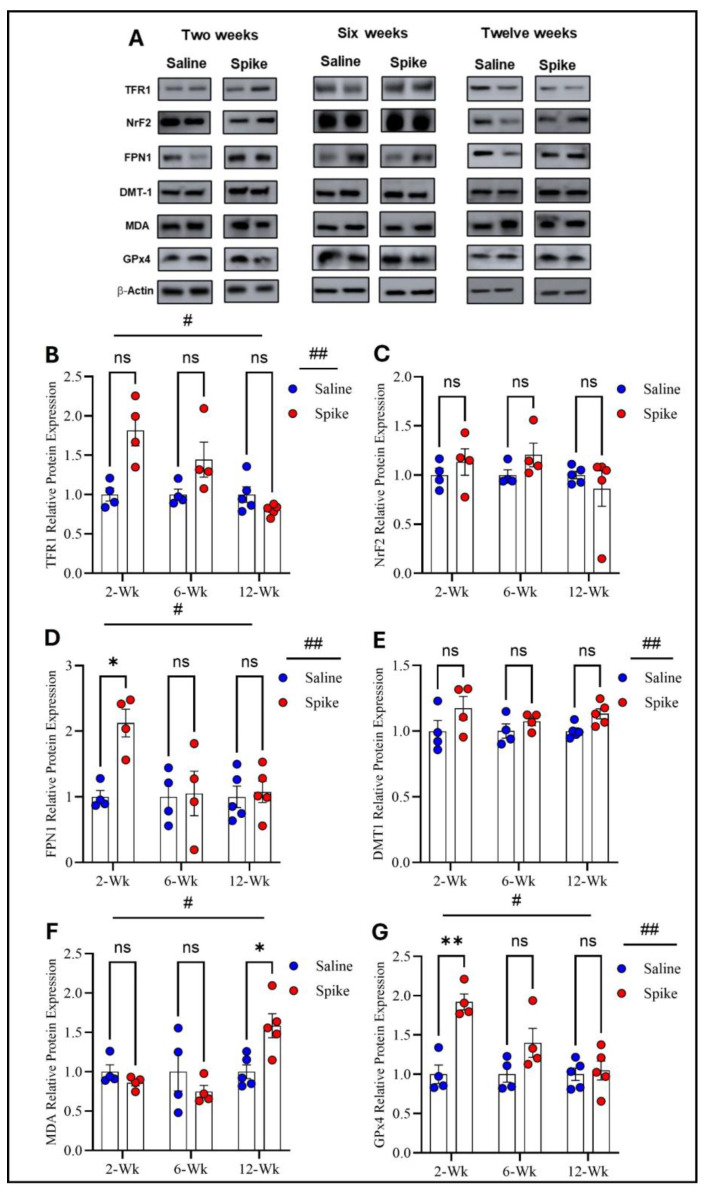 Figure 4
Figure 4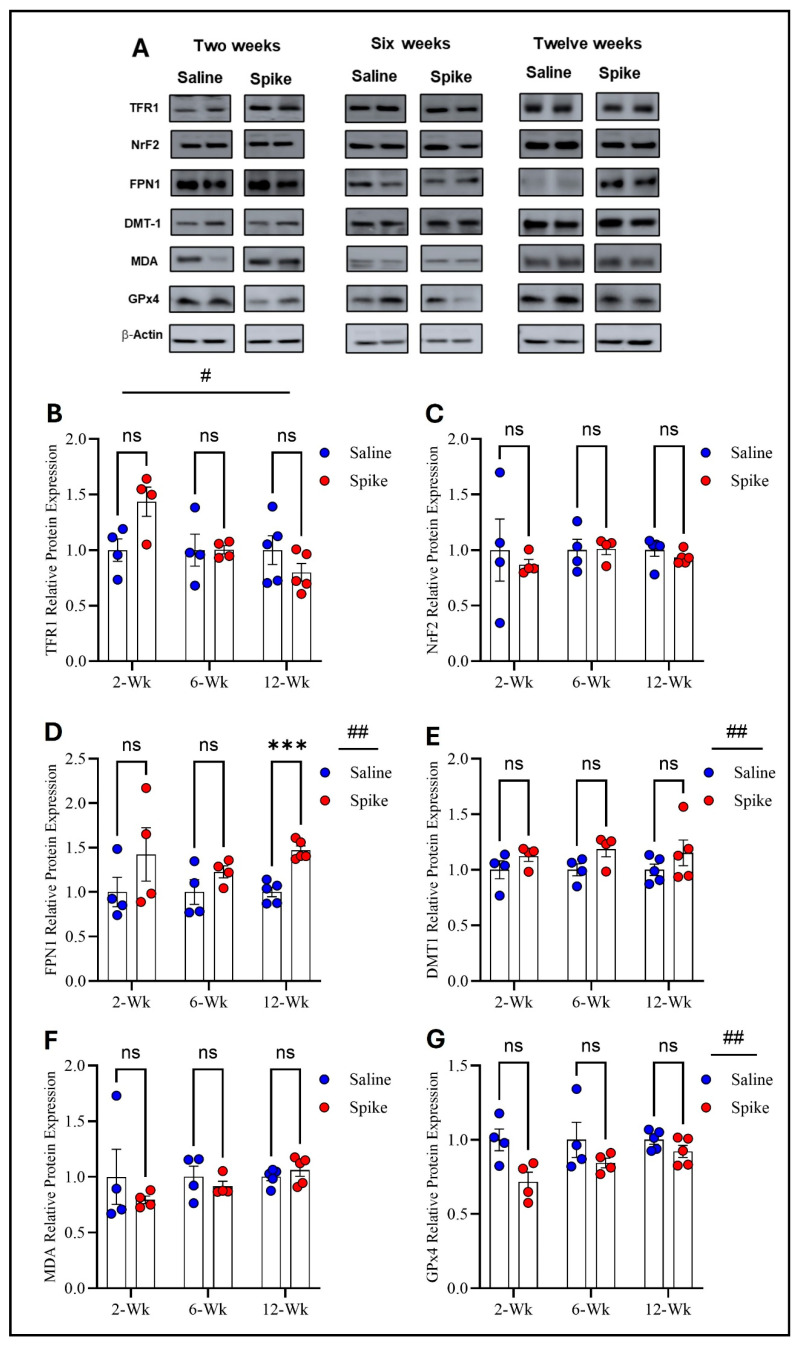 Figure 5
Figure 5 Figure 6
Figure 6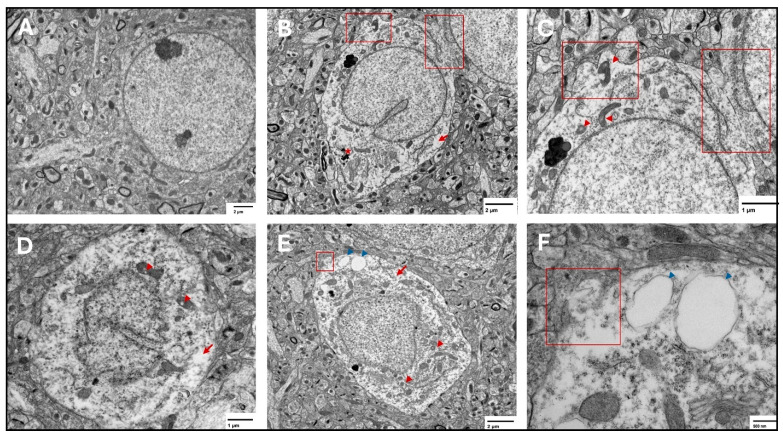 Figure 7
Figure 7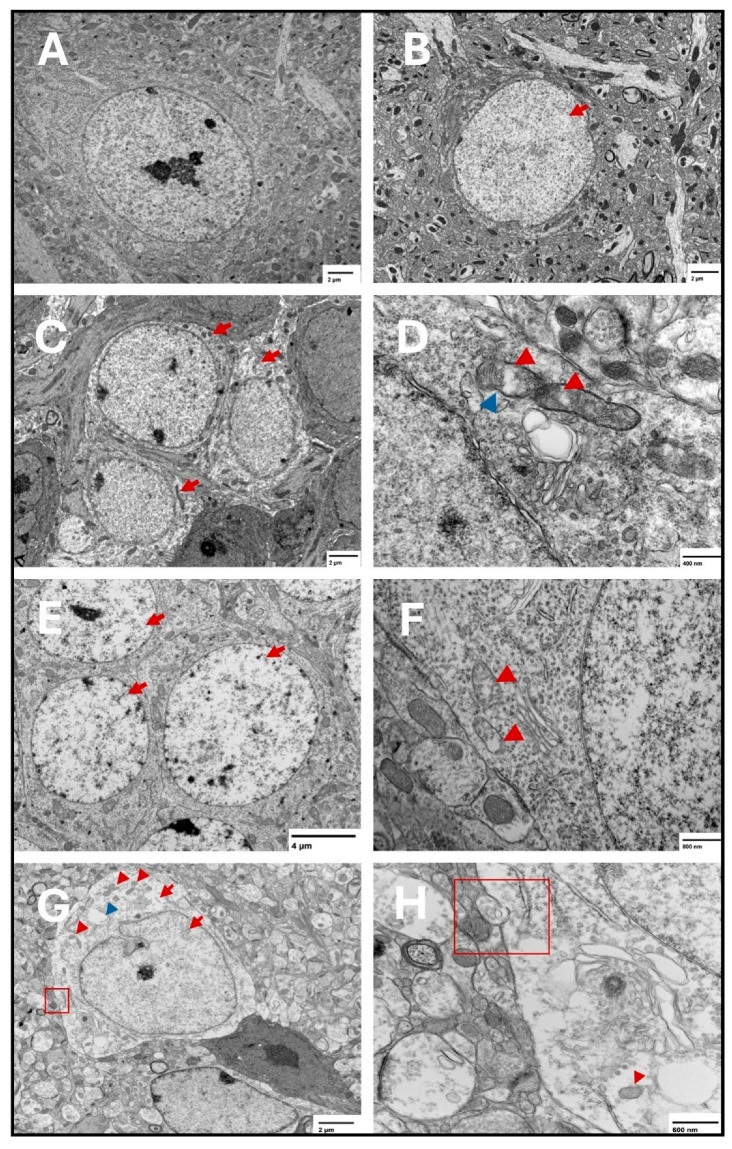 Figure 8
Figure 8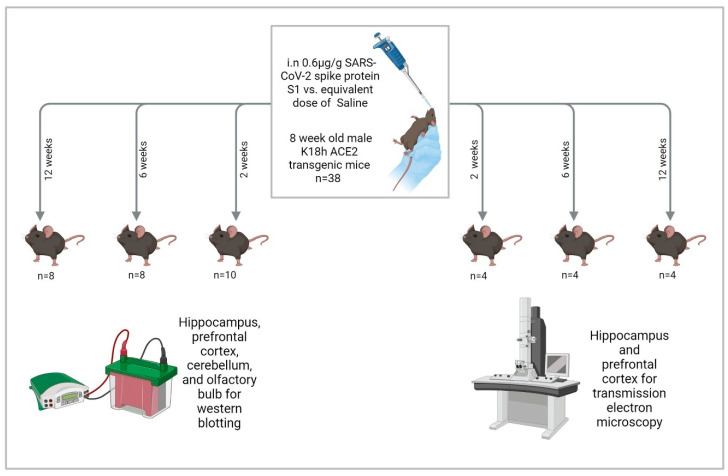 Figure 9
Figure 9- —Department of Psychiatry and Psychology at the Mayo Clinic, Arizona
- —Edli Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFerroptosis and cancer prognosis · Tryptophan and brain disorders · Single-cell and spatial transcriptomics
1. Introduction
Ferroptosis is an iron-dependent form of cell death that was only discovered a decade ago [1]. The occurrence of ferroptosis has three prerequisites: first, iron dysregulation and accumulation of free labile iron capable of redox (oxidation-reduction) reactions and the production of reactive oxygen species (ROS); second, the failure of antioxidant lipid peroxide repair systems, especially the glutathione (GSH)–glutathione peroxidase 4 (GPx4) system, in the face of unopposed production of ROS; and third, lipid peroxidation and the production of lipid ROS, which is amplified by iron and failed antioxidant systems. Lipid peroxidation is considered the executor of ferroptosis, which ultimately produces highly toxic lipid peroxidation degradation products, such as malondialdehyde (MDA), that react with cell proteins and DNA, altering their structure and function and fostering a state of cytotoxicity. In addition, lipid peroxidation of membrane phospholipid polyunsaturated fatty acids (PUFAs) compromises the cell membrane integrity, leading to its thinning and rupture [1,2] (Figure 1).
Regulated ferroptosis plays a critical role in development, normal physiological aging, tumor suppression, and antiviral and overall immune function [3]. However, specific cellular pathophysiological mechanisms trigger uncontrolled ferroptosis in multiple neuropsychiatric disorders, such as depression and anxiety-like behavior [4,5], bipolar disorder [6], ischemic stroke [7,8], traumatic brain injury and its neuropsychiatric complications [9,10], Alzheimer’s [11,12,13], Parkinson’s [14,15], multiple sclerosis [16,17], amyotrophic lateral sclerosis [18,19], and possibly post-COVID syndrome [20].
Post-COVID syndrome, post-acute sequelae of Severe Acute Respiratory Syndrome Coronavirus type 2 (SARS-CoV-2) infection, and long COVID all refer to a group of neurological, cognitive, and psychiatric manifestations that persist for more than 12 weeks after recovery from acute SARS-CoV-2 infection regardless of illness severity and cannot be explained by an alternative diagnosis [21,22,23,24]. The most common manifestations include fatigue, anxiety, depression, headache, and cognitive impairment [25]. The magnitude of the post-COVID syndrome as a public health crisis cannot be underestimated. The incidence of post-COVID syndrome six months after recovery from acute infection among 236,379 patients was 33.6% (95% CI 33.17–34.07) and 46.4% (44.78–48.09) in patients who had been admitted to an intensive care unit [26]. A nationwide population cohort study of 198,096 Scottish adults estimated the prevalence of post-COVID syndrome to be 6.6–10.3% [27]. The pathophysiology of post-COVID syndrome remains under investigation [28]. However, all three elements of ferroptosis are documented in various studies.
Iron dysregulation serves as a protective mechanism during the acute illness since iron is a transition metal that acts as a catalyst for reactions that require electron transfer [29], such as the RNA-dependent RNA polymerase [30]. SARS-CoV-2 utilizes this iron-containing enzyme to replicate and synthesize its viral RNA. To deprive the virus of this essential element, the body sequesters iron into ferritin [31]. However, excessive iron storage into ferritin could be detrimental if it triggers nuclear receptor coactivator 4 (NCOA4)-mediated ferritinophagy [32]. Massive release of iron from ferritin generates oxidative stress and ferroptosis [20]. This state of acute iron dysregulation during COVID-19 illness manifests as high serum ferritin concentration and low serum iron content, signaling a worse outcome [33,34,35,36,37]. In addition, patients with moderate to severe COVID-19 who later developed post-COVID manifestations were found to have overexpression of monocyte iron homeostasis genes [38].
Failure of antioxidant systems is reported in acute SARS-CoV-2 infection in the form of glutathione (GSH) deficiency and a decrease in the total antioxidant capacity against the unopposed rise of oxidative stress [39,40,41,42,43]. GSH deficiency has been linked with COVID-19 severity and mortality [44]. In addition, SARS-CoV-2 directly suppresses mRNA expression of GPx4 [45] and indirectly affects its function by causing selenium deficiency, which was reported in COVID-19 patients and linked to higher mortality odds [46,47]. Selenium is integral for the function of GPx4, a selenoprotein that efficiently eliminates lipid hydroperoxides (lipid-ROS) [48]. These reports highlight an evident impact on both GSH and GPx4, the primary ferroptosis brakes [49], during COVID-19 acute infection. Nonetheless, the disruption of antioxidant defenses has been reported in post-COVID-19 patients, albeit less frequently. In a proof-of-concept study involving 120 post-COVID patients, Al-Hakeim et al. reported a significant correlation between post-COVID neuropsychiatric scores and reduced peripheral antioxidant defenses [50]. Using magnetic resonance spectroscopy, Saleh et al. detected lower levels of GSH in the frontal gray matter of 29 post-COVID patients, extending the finding of a failed antioxidant system to the brain [51]. Poletti et al. also depicted lower anterior cingulate cortex concentrations of GSH in 49 COVID-19 survivors, which are associated with increased depressive symptoms [52].
Lipid peroxidation biomarkers, including MDA and 4-hydroxynonenal (HNE-4), have been repeatedly linked with COVID-19 severity and mortality [53,54,55], besides a dysregulation of the lipid landscape [56,57]. These findings manifest in post-COVID-19 patients, where López-Hernández et al. found higher levels of several species of plasma phospholipid, phosphatidylcholines, and sphingomyelins in 20 post-COVID-19 patients compared to controls two years after recovery [58]. Also, Garrido et al. reported higher levels of other subclasses of plasma phospholipid, lysophosphatidylglycerols, and phosphatidylethanolamines in 147 post-COVID-19 patients up to 15 months after infection [59]. Despite these studies addressing plasma, not brain phospholipids, their findings could still be an argument for the occurrence of ferroptosis since the availability of phospholipid PUFA commands the sensitivity to ferroptosis [60,61]. This data collectively demonstrates the potential presence of ferroptosis as part of the COVID-19/post-COVID-19 pathophysiology.
To investigate changes in ferroptotic markers, we administered the SARS-CoV-2 spike protein S1 subunit intranasally to the K18-hACE2 transgenic mice at 8 weeks of age. We examined differences in protein expression levels of multiple ferroptotic markers (TRF1, DMT1, FPN1, NrF2, GPx4, and MDA-conjugated proteins) at four brain regions (hippocampus, prefrontal cortex, cerebellum, and olfactory bulb) at three time points: 2, 6, and 12 weeks post-S1 administration. We also examined ferroptotic features of subcellular structures in the hippocampus and prefrontal cortex using transmission electron microscopy (TEM) at the same three time points post-S1 administration. We used the spike protein and not the virus itself because administering the virus to ACE2 transgenic mice is lethal [62]. In addition, the spike protein alone directly impacts human cells [63], with one study showing that persistence of the spike protein up to 12 months was associated with post-COVID syndrome [64]. Furthermore, in animal models, the spike protein induces behavioral alterations reminiscent of post-COVID neuropsychiatric manifestations [65,66].
2. Results
2.1. Dynamic Protein Expression Changes of Ferroptosis Markers
2.1.1. In the Hippocampus
Two-way ANOVA identified significant main effects of time on both TFR1 (F_1.584,15.05_ = 4.972, p = 0.028) and FPN1 protein expression (F_2,12_ = 6.879, p = 0.0102), as well as significant treatment × time interactions for TFR1 (F_2,19_ = 4.972, p = 0.0183) and FPN1 (F_2,12_ = 6.081, p = 0.015), indicating that expression of both proteins and the effect of S1 exposure differed across time points. No significant main effect of treatment was observed for either protein when averaged across all time points. Post hoc Šídák-adjusted comparisons showed no statistically significant differences between saline and spike groups for TFR1 at individual time points. In contrast, post hoc analyses for FPN1 demonstrated a significant increase in expression in the spike group at 2 weeks compared to controls (adjusted p = 0.013), with no significant differences detected at 6 or 12 weeks. In contrast to TFR1 and FPN1, no statistically significant effects were detected for DMT1, NrF2, MDA-conjugated proteins, or GPx4 protein expression. Two-way ANOVA identified no significant main effects of time or treatment or any significant treatment × time interactions for any of these markers (Figure 2 and Supplementary Table S1).
2.1.2. In the Prefrontal Cortex
Regarding TFR1, FPN1, and NrF2 protein expression, two-way ANOVA revealed no significant main effects of time or treatment or any significant interactions between the two factors. However, two-way ANOVA showed distinct patterns for DMT1, MDA-conjugated proteins, and GPx4 protein expression. For DMT1, two-way ANOVA identified significant main effects of time (F_1.541,15.41_ = 20.12, p = 0.0001) and treatment (F_1,20_ = 12.61, p = 0.002), as well as a significant treatment × time interaction (F_2,20_ = 20.12, p < 0.0001), indicating that DMT1 expression varied across time points and that the impact of S1 differed by time. Post hoc Šídák-adjusted comparisons demonstrated significantly higher DMT1 expression in the spike group at 2 weeks compared to controls (adjusted p = 0.002), with no significant differences at 6 or 12 weeks. For MDA-conjugated proteins, two-way ANOVA exhibited a significant main effect of time (F_1.858,11.15_ = 6.411, p = 0.015) and a significant treatment × time interaction (F_2,12_ = 6.712, p = 0.011), while the main effect of treatment was not significant. However, post hoc Šídák-adjusted comparisons did not identify significant differences between saline and spike groups at 2, 6, or 12 weeks. For GPx4, two-way ANOVA identified a significant main effect of treatment (F_1,20_ = 13.60, p = 0.001), with no significant main effect of time (F_1.994,19.94_ = 3.294, p = 0.058) and no significant interaction between the two factors (F_2,20_ = 3.294, p = 0.058). Post hoc Šídák-adjusted comparisons were not significant at any individual time points (Figure 3 and Supplementary Table S2).
2.1.3. In the Cerebellum
Two-way ANOVA identified significant main effects of time (F_1.654,16.54_ = 8.343, p = 0.004) and treatment (F_1,20_ = 11.44, p = 0.003) on TFR1 protein expression, as well as a significant treatment × time interaction (F_2,20_ = 8.343, p = 0.002). However, post hoc Šídák-adjusted comparisons between saline and spike groups at 2, 6, and 12 weeks did not reach statistical significance. For FPN1, two-way ANOVA also showed significant main effects of time (F_1.829,18.29_ = 4.339, p = 0.03) and treatment (F_1,20_ = 6.266, p = 0.02), with significant treatment × time interaction (F_2,20_ = 4.339, p = 0.027). Post hoc analyses exhibited significantly higher FPN1 expression in the spike group at 2 weeks compared to controls (adjusted p = 0.021), with no significant differences at 6 or 12 weeks. For DMT1, a significant main effect of treatment was detected (F_1,8_ = 6.296, p = 0.036), while the main effect of time and the treatment × time interaction were not significant. Post hoc comparisons did not identify significant differences at individual time points. NrF2 showed no significant main effects or treatment × time interaction by two-way ANOVA and no significant post hoc differences at 2, 6, or 12 weeks. For MDA-conjugated proteins, two-way ANOVA revealed a significant main effect of time (F_1.830,10.98_ = 9.883, p = 0.004) and a significant treatment × time interaction (F_2,12_ = 10.74, p = 0.0021), while the main effect of treatment was not significant. Post hoc analyses identified a significant increase in MDA-conjugated protein expression in the spike group at 12 weeks (adjusted p = 0.043), with no significant differences at earlier time points. Finally, GPx4 exhibited significant main effects of time (F_1.842, 11.05_ = 15.17, p = 0.0008) and treatment (F_1,8_ = 12.47, p = 0.0077), along with significant treatment × time interaction (F_2,12_ = 14.87, p = 0.0006). Post hoc Šídák-adjusted comparisons demonstrated significantly higher GPx4 expression in the spike group at 2 weeks (adjusted p = 0.0033), with no significant differences at 6 or 12 weeks (Figure 4 and Supplementary Table S3).
2.1.4. In the Olfactory Bulb
Two-way ANOVA identified a significant main effect of time (F_1.998,11.99_ = 11.93, p = 0.001) on TFR1 expression and a significant treatment × time interaction (F_2,12_ = 8.72, p = 0.004), while no significant main effect of treatment was identified. Post hoc Šídák-adjusted comparisons between saline and spike groups at 2, 6, and 12 weeks were not significant. For FPN1, a significant main effect of treatment was observed (F_1,8_ = 8.329, p = 0.02), whereas the main effect of time and the treatment × time interaction were not significant. Post hoc analyses identified a significant increase in FPN1 in the spike group at 12 weeks (adjusted p = 0.0006), with no significant differences at 2 or 6 weeks. For DMT1, two-way ANOVA also detected a significant main effect of treatment (F_1,20_ = 5.962, p = 0.024), with no significant main effect of time or treatment × time interaction and no significant post hoc differences at individual time points. NrF2 and MDA-conjugated proteins showed no significant main effects of time, treatment, or treatment × time interactions by two-way ANOVA and no significant post hoc differences at any time point. Finally, GPx4 exhibited a significant main effect of treatment (F_1,8_ = 8.051, p = 0.021) with no significant main effect of time or treatment × time interaction. Post hoc Šídák-adjusted comparisons were not significant at individual time points (Figure 5 and Supplementary Table S4).
The overall summary of the results is demonstrated in Figure 6.
2.2. Ferroptosis Features Detected by Transmission Electron Microscopy
At the three tested points, features of ferroptosis were observed in the prefrontal cortex (Figure 7) and hippocampus (Figure 8), including electron-lucent nuclei and cytoplasm, cell membrane disruption, and mitochondrial changes that included shrunken mitochondria, disrupted mitochondrial cristae, and ruptured outer membranes.
3. Discussion
The present study demonstrates that a single intranasal (IN) administration of SARS-CoV-2 spike protein S1 to K18-hACE2 transgenic mice elicits a persistent molecular response in ferroptosis-related markers, alongside ultrastructural features consistent with ferroptotic injury in the hippocampus and prefrontal cortex [3,67]. This response is both time-dependent and brain-region-specific, which is shown in the olfactory bulb, prefrontal cortex, hippocampus, and cerebellum, where a distinct statistical pattern was observed across regions and tested time points.
In the hippocampus, significant treatment × time interactions were identified for both TFR1 and FPN1, indicating a time-dependent treatment influence on iron-handling markers. FPN1 showed a significant increase at 2 weeks in the spike group, while TFR1 showed a trend toward higher expression in the spike group at 6 weeks. These findings suggest that hippocampal iron-export machinery may be engaged early after S1 exposure (increased FPN1), while TFR1 changes may be more temporally related rather than dominated by a single time point. Notably, hippocampal DMT1, NrF2, GPx4, and MDA-conjugated proteins did not show statistically significant effects, indicating that in the hippocampus and within the tested markers, iron-handling shifts were more prominent than detectable antioxidant or lipid peroxidation markers across the three time points. Dysregulation of iron-related markers aligns with the hippocampus high metabolic demand and iron content, which may disrupt hippocampal synaptic integrity and neuronal viability, potentially contributing to memory deficits and cognitive impairment reported following COVID19 infection [68,69].
In the prefrontal cortex, TFR1, FPN1, and NrF2 did not show significant time, treatment, or interaction effects. In contrast, DMT1 showed higher expression in the spike group at 2 weeks as well as a robust time and treatment effects with a significant treatment × time interaction. This pattern supports early enhancement of iron-import capacity in the prefrontal cortex following S1 exposure. Additionally, significant time and treatment × time effects for MDA-conjugated proteins, without significant post hoc differences at individual time points, suggest temporally related impact of S1 on oxidative damage that may not be strongly localized to a single time point after multiplicity adjustment. Finally, GPx4 showed a significant main effect of treatment, with a non-significant trend of increased expression at 6 weeks. Together, these results indicate that in the prefrontal cortex, S1 exposure is accompanied by early iron-transport modulation (via DMT1) and treatment-associated differences in GPx4, consistent with engagement of antioxidant defenses.
The cerebellum showed the most consistent evidence for time-dependent modulation across multiple ferroptosis-related markers. Tested iron-related markers, TFR1, FPN1, and DMT1 showed significant treatment effects, while only TFR1 and FPN1 showed significant treatment × time interaction. No statistically significant differences were observed for TFR1 between the spike and saline groups at any time point. FPN1 showed a clear post hoc increase at 2 weeks. Similarly, DMT1 did not show statistically significant differences at individual time points. Importantly, cerebellar MDA-conjugated proteins showed significant interaction effects, with a significant increase in the spike group at 12 weeks, pointing to late-emerging oxidative damage signal in the cerebellum. In parallel, GPx4 displayed a significant treatment effect, significant interaction, and a significant post hoc increase at 2 weeks. Taken together, the cerebellum demonstrates an early pattern compatible with compensatory antioxidant engagement (increased GPx4 and increased FPN1 suggesting iron export) alongside later evidence of lipid peroxidation-associated damage (increase in MDA-conjugated proteins at 12 weeks). This temporal change suggests early protective responses that may be insufficient to prevent later oxidative injury under persistent or evolving stressors, suggesting delayed ferroptotic vulnerability in this region. This delayed vulnerability in the cerebellum could manifest as subtle motor or coordination deficits, which is reported in post-COVID-19 patients [70]. However, the upregulation of GPx4 is inconsistent with the classic ferroptotic profile of antioxidant failure. In addition, the lack of TEM assessment of cerebellar ferroptotic features renders the data in the cerebellum to most likely indicate cellular stress and compensatory resistance, rather than the execution of ferroptosis.
In the olfactory bulb, TFR1 demonstrated a significant treatment × time interaction but without significant post hoc differences at individual time points. FPN1 showed a significant main effect of treatment and a significant post hoc increase at 12 weeks, suggesting altered iron export capacity at later stages. DMT1 showed a significant main effect of treatment without significant post hoc week-specific differences. NrF2 and MDA-conjugated proteins did not show significant effects. GPx4 displayed a significant main effect of treatment. Collectively, these findings suggest that the olfactory bulb displays treatment-associated shifts in iron-handling (notably FPN1 at 12 weeks). These changes imply delayed olfactory bulb vulnerability after S1 administration. The treatment-related effect of the spike protein on multiple ferroptotic markers in the olfactory bulb could underline the olfactory dysfunction, which is reported to persist in long-COVID conditions [71,72].
Notably, the measured markers are heavily implicated in ferroptosis machinery. TFR1 elevated expression is a strong indication of ferroptosis since TFR1 is considered to be a specific ferroptosis marker that can even increase sensitivity to ferroptosis [73,74,75,76]. Interestingly, TFR1 was found to act as an alternative receptor for the SARS-CoV-2 spike protein to allow cellular viral entry [77]. In addition, DMT-1 overexpression emphasizes ferroptotic changes as it acts as a ferroptosis driver in early brain injury as well as in other contexts [78,79,80,81,82]. GPx4, a ferroptosis brake, displayed region-specific favoritism, where it increased in the cerebellum and decreased in the olfactory bulb at the same time (two weeks). This could indicate an earlier vulnerability of the olfactory bulb and room for compensatory mechanisms in the cerebellum [83]. Furthermore, MDA-conjugated proteins not only mark lipid peroxidation occurrence but also indicate that MDA formed covalent adducts on proteins and caused oxidative lipid-derived protein damage [84].
Notably, interaction effects without strong week-specific post hoc contrasts for certain markers suggest that some treatment influences manifest as distributed temporal shifts rather than isolated peaks at one time point.
Our in vivo results add to the current literature which shows that the spike protein induces ferroptosis in different cell types [THP-1-derived macrophages [85], Huh7 and Calu-3 cells [83], and H9C2 cells [86]]. Here we show that a single dose of the spike protein administered intranasally to the K18-hACE2 transgenic mice induces long-lasting protein expression changes in several ferroptotic and anti-ferroptosis markers in different brain regions. The K18h-ACE2 mice have been used to model COVID-19 infection [87,88] through the IN administration of the SARS-CoV-2 virus itself. However, the mortality rate after IN viral administration was high, which did not allow for the disease progression to be studied [89,90,91]. Intracerebroventricular administration of the SARS-CoV-2 spike protein has been used to model post-COVID syndrome but not in K18-hACE2 mice [65]. Rhea et al. assessed whether the SARS-CoV-2 spike protein reaches the brain by intravenous (IV) and IN administration. They reported that the SARS-CoV-2 spike protein reaches the brain at a much lower concentration by IN compared to IV administration [92]. However, they used a dose of 12.5 ng/mouse, while we used a dose of (0.6 μg/g, or 12 μg/20g mouse). We used the S1 subunit of the SARS-CoV-2 spike protein based on iterative reports of how it is implicated in the pathogenesis of post-COVID syndrome [93]. The full-length spike protein is composed of two subunits, S1 and S2, with a furin cleavage site (FCS). S1 must be first cleaved by host cell protease to allow binding to the ACE2 receptor and viral entry into the cell [94]. Therefore, the use of K18-hACE2 mice was most relevant since these mice express excess human ACE2 to which the SARS-CoV-2 spike protein RBD mainly binds [95]. Intriguingly, when the SARS-CoV-2 spike protein RBD binds to ACE-2 receptors, ACE2 receptors are internalized and degraded, which in turn can increase angiotensin II (Ag II) which tends to act on angiotensin receptor 1 (ATR1). The activation of ATR1 would activate NADPH oxidase (NOX), promoting the production of hydrogen peroxides, lipid peroxidation, and ferroptosis [96]. The implication of ACE2 receptors in the induction or mitigation of ferroptosis was repeatedly reported [97,98]. However, these mechanisms remain to be investigated.
Despite the variability in the molecular response, ultrastructural examination with transmission electron microscopy showed classical ferroptotic features, including cell membrane disruption, shrunken mitochondria, disrupted cristae, and translucent nuclei and cytoplasm in the hippocampus and prefrontal cortex suggesting failed protective mechanisms against ferroptosis [3,67]. These ultrastructural findings, limited to the hippocampus and prefrontal cortex, support the notion that S1 exposure is associated with persistent cellular injury patterns compatible with ferroptosis in these regions, even when biochemical markers vary by region and time point or fail to reach week-specific statistical significance after correction.
4. Methods
4.1. Mice
All studies were approved by the Mayo Clinic Institutional Animal Care and Use Committee under Protocol number A00006724-22-R25 (7 December 2022) and carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. We used a total of 38 male K18-hACE2 transgenic mice (B6.Cg-Tg (K18-ACE2) 2Prlmn/J) purchased from the Jackson Laboratory (Bar Harbor, ME, USA). Mice were group-housed, 4–5 mice per cage, with available food and water ad libitum, at a temperature of 23–25 °C and humidity of 50–60% with a 14 h/10 h light/dark cycle (light: 5:00–19:00, dark: 19:00–5:00).
4.2. SARS-CoV-2 Spike Protein Receptor Binding Domain (RBD) Subunit 1 (S1) Administration
We administered 0.6 μg/g intranasal SARS-CoV-2 spike protein subunit 1 (S1) (R&W Systems, Singapore, cat. no. 10522-CV) or an equivalent volume of phosphate-buffered saline (PBS) to mice once they reached 8 weeks of age. This dose is about 12 μg/20 g mouse, which is comparable to other studies [65,66,99]. S1 was reconstituted in PBS and administered to mice anesthetized with isoflurane (3–4% for induction and 1.5–2% for maintenance) in oxygen using a precision vaporizer. Reconstituted S1 was administered as 3 μL drops in the left nostril and 3 μL drops in the right nostril, separated by 15 s, then the same 2 × 3 μL drop administration was repeated after 30 s until the full dose had been administered [100]. Mice were euthanized at 2, 6, and 12 weeks after S1 administration by cervical dislocation followed by rapid decapitation, brain dissection on ice, and flash freezing of brain tissues in liquid nitrogen. We collected the olfactory bulbs, prefrontal cortices, hippocampi, and cerebellums bilaterally. Tissues were stored in a −80 °F freezer until they were assayed (Figure 9).
4.3. Western Blotting
Tissue extracts were isolated using RIPA lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 10 mM EDTA, 1% Triton X-100), supplemented with protease and phosphatase inhibitors (Thermo Scientific, Waltham, MA, USA, cat. No. PIA32959). Total protein concentrations were quantified using Pierce BCA Protein Assay Kit (Thermo Scientific, Waltham, MA, USA, cat. No. 23225) following the manufacturer’s instructions. Equal amounts of protein (30 µg per sample) were resolved on 4–20% Mini-PROTEAN^®^ TGX™ precast protein gels (Bio-Rad, Hercules, CA, USA, cat. no. 4561096) and transferred onto PVDF membranes (Thermo Scientific, Waltham, MA, USA, cat. no. 88518). Membranes were blocked for 1 h at room temperature with blocking solution (Tris-buffered saline with 0.05% Tween (TBS-T) containing 5% skim milk), then incubated overnight at 4 °C with primary antibodies. We used primary antibodies for transferrin receptor 1 (TFR1) (Cell Signaling Technology, Danvers, MA, USA, Cat. no. 46222) (1:1000), ferroportin 1 (FPN1) (Proteintech, Chicago, IL, USA, Cat. no. 26601-1-AP) (1:2000), divalent metal transporter-1 (DMT-1) (Abcam, Washington, DC, USA, Cat. no. ab55735) (1:1000), MDA-conjugated proteins (Abcam, Washington, DC, USA, Cat. no. ab243066) (1:1000), Nuclear Factor Erythroid 2-Related Factor 2 (NrF2) (Abcam, Washington, DC, USA, Cat. no. ab62352) (1:2000), GPx4 (Cell Signaling Technology, Danvers, MA, USA, Cat. no. 52455) (1:1000), and beta-actin (Cell Signaling Technology, Danvers, MA, USA, Cat. no. 3700T) (1:1000). Following 3 washes with TBS-T, membranes were further incubated with either horseradish peroxidase (HRP)-conjugated goat anti-rabbit antibody (BioRad, Hercules, CA, USA, Cat. no. 1706515) (1:7500) or horseradish peroxidase (HRP)-conjugated goat anti-mouse antibody (VWR International LLC, Radnor, PA, USA, Cat. no. P001982153) (1:5000) in blocking solution for 2 h at room temperature. For the assessment of beta-actin expression, membranes were stripped with 0.2 M NaOH, washed, and incubated with beta-actin antibody using the same procedure as above. Blots were incubated in SuperSignal™ West Femto Maximum Sensitivity Substrate (Thermo Scientific, Waltham, MA, USA, cat. no. 34094). Bands were detected with a digital GE Amersham Imager 680. Band intensities were measured in arbitrary units using QuantityOne software V.4.6.8 (BioRad, Hercules, CA, USA). Band intensity values were normalized over the beta-actin band of the same lane. Relative protein expression was determined as the ratio of each sample over the average of controls.
Statistical analyses were performed unblinded using GraphPad Prism software (V.10.4.1, San Diego, CA, USA). For Western blot experiments, relative protein expression was analyzed using a two-way ANOVA, with factor treatment (saline vs. SARS-CoV-2 S1) and time (2, 6, and 12 weeks). When main effects and/or treatment × time interactions were detected, Šídák-adjusted multiple comparisons were performed to compare saline and spike groups within each time point. Data were presented as mean ± SEM. Results were considered statistically significant when p < 0.05.
4.4. Transmission Electron Microscopy
Mice were transcardially perfused with 4% paraformaldehyde (PFA). After perfusion, brains were dissected and both hippocampus and prefrontal cortex were cut into 2–4 mm^3^ pieces and placed into 4% paraformaldehyde + 1% glutaraldehyde (in 0.1 M phosphate buffer pH 7.4) fixative immediately. Then, tissues were post-fixed in osmium tetroxide, dehydrated, and embedded in resin. Ultrathin sections were cut, stained, and imaged on a JEOL 1400 transmission electron microscope (JEOL Ltd., Akishima, Tokyo, Japan) at an accelerating voltage of 80 kV.
5. Conclusions
Collectively, our results suggest that a single intranasal dose of the SARS-CoV-2 spike protein induces alterations of ferroptotic markers through temporal and region-specific changes in a targeted panel of iron-handling, antioxidant, and lipid peroxidation-related markers, alongside ultrastructural ferroptotic features in brain tissue. However, these results raise multiple questions, including the following: What are the clinical implications for such changes in the ferroptotic markers? What are the consequences for the dichotomy of ferroptosis and anti-ferroptotic compensatory mechanisms, especially since ferroptosis is implicated in multiple pathologies yet has beneficial outcomes in certain cases such as tumorigenesis [101]?
Several limitations should be considered. First, the sample size at each time point (n = 4–5) may reduce the power to detect week-specific post hoc differences even when ANOVA identifies significant time or interaction effects, especially with the unblinded nature of the analysis. Second, experimental design introduces potential observer bias, particularly in the selection of representative microscopy images and Western blot quantification, and limits the statistical power and generalizability of the findings. Future studies must validate these results in larger, blinded cohorts. Third, our marker panel captures key nodes of ferroptosis biology. However, more depth in investigating the ferroptosis machinery is needed by examining more ferroptosis-related markers and examining the effect of anti-ferroptosis agents such as ferrostatin-1. Fourth, Western blotting reflects bulk tissue protein levels and cannot resolve cell-type-specific effects. Fifth, ultrastructural examination of ferroptotic features was limited to the hippocampus and prefrontal cortex and depended on qualitative assessment rather than quantitative one. Therefore, future studies will benefit from expanding ferroptosis markers, incorporating anti-ferroptotic interventions (e.g., ferrostatin-1), assessing earlier and later time points. In addition, utilizing cell-type-resolved approaches such as single-cell/spatial transcriptomics or proteomics is needed to determine whether these region-specific signals arise from specific cell populations, which would better clarify our findings and uncover the underlying pathological mechanisms of the spike protein in relation to post-COVID syndrome.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dixon S.J. Lemberg K.M. Lamprecht M.R. Skouta R. Zaitsev E.M. Gleason C.E. Patel D.N. Bauer A.J. Cantley A.M. Yang W.S. Ferroptosis: An iron-dependent form of nonapoptotic cell death Cell 20121491060107210.1016/j.cell.2012.03.04222632970 PMC 3367386 · doi ↗ · pubmed ↗
- 2Dixon S.J. Stockwell B.R. The hallmarks of ferroptosis Annu. Rev. Cancer Biol.20193355410.1146/annurev-cancerbio-030518-05584441613499 PMC 12851585 · doi ↗ · pubmed ↗
- 3Stockwell B.R. Ferroptosis turns 10: Emerging mechanisms, physiological functions, and therapeutic applications Cell 20221852401242110.1016/j.cell.2022.06.00335803244 PMC 9273022 · doi ↗ · pubmed ↗
- 4Cao H. Zuo C. Huang Y. Zhu L. Zhao J. Yang Y. Jiang Y. Wang F. Hippocampal proteomic analysis reveals activation of necroptosis and ferroptosis in a mouse model of chronic unpredictable mild stress-induced depression Behav. Brain Res.202140711326110.1016/j.bbr.2021.11326133775778 · doi ↗ · pubmed ↗
- 5Dai Y. Guo J. Zhang B. Chen J. Ou H. He R.-R. So K.-F. Zhang L. Lycium barbarum (Wolfberry) glycopeptide prevents stress-induced anxiety disorders by regulating oxidative stress and ferroptosis in the medial prefrontal cortex Phytomedicine 202311615486410.1016/j.phymed.2023.15486437182278 · doi ↗ · pubmed ↗
- 6Yehia A. Melhuish Beaupre L.M. Ho M.C. Biernnacka J.M. Frye M.A. Abulseoud O.A. Ferroptosis as a potential molecular mechanism of bipolar disorder Transl. Psychiatry 20251520510.1038/s 41398-025-03429-w 40537482 PMC 12179299 · doi ↗ · pubmed ↗
- 7Cui Y. Zhang Y. Zhao X. Shao L. Liu G. Sun C. Xu R. Zhang Z. ACSL 4 exacerbates ischemic stroke by promoting ferroptosis-induced brain injury and neuroinflammation Brain Behav. Immun.2021933123213344473310.1016/j.bbi.2021.01.003 · doi ↗ · pubmed ↗
- 8Yang K. Zeng L. Yuan X. Wang S. Ge A. Xu H. Zeng J. Ge J. The mechanism of ferroptosis regulating oxidative stress in ischemic stroke and the regulation mechanism of natural pharmacological active components Biomed. Pharmacother.202215411361110.1016/j.biopha.2022.11361136081288 · doi ↗ · pubmed ↗