The Period (per) Gene Plays an Important Role in Regulating Circadian Oscillation and Ovarian Development in the Ridgetail White Prawn, Exopalaemon carinicauda
Caijuan Tian, Yunhan Feng, Jixuan Zhu, Xuanjian Chen, Wanxin Ma, Panpan Niu, Hao Zhong, Huan Gao, Yuquan Li

TL;DR
This study shows that the period gene in ridgetail white prawns regulates circadian rhythms and ovarian development, offering insights into crustacean reproduction.
Contribution
The study identifies Ec-per as a key regulator of circadian rhythms and ovarian development in crustaceans through gene expression and functional analysis.
Findings
Ec-per exhibits rhythmic expression under various light-dark cycles, indicating its role as an endogenous oscillator.
RNA interference of Ec-per reduces expression of circadian genes and ecdysone receptors, affecting ovarian development.
In situ hybridization shows Ec-per involvement in oocyte proliferation and nutrient accumulation.
Abstract
This study elucidates the characteristics of the circadian clock gene (period) in the ridgetail white prawn, Exopalaemon carinicauda (Ec-per). Ec-per exhibited rhythmic expression under multiple light–dark (L–D) cycles, including constant darkness, confirming its role as an endogenous oscillator. Functional analysis using RNA interference indicated that Ec-per, as an important component of the circadian feedback loop, influenced reproductive physiology by synergistically regulating circadian genes (timeless and cryptochrome1) and upregulating ecdysone receptors. In situ hybridization results further suggested Ec-per is involved in oogenesis. Together, these findings highlight the important role of Ec-per in circadian rhythm regulation and ovarian development, offering new insights into photoperiod-mediated reproductive regulation mechanisms in crustaceans. To explore the function of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3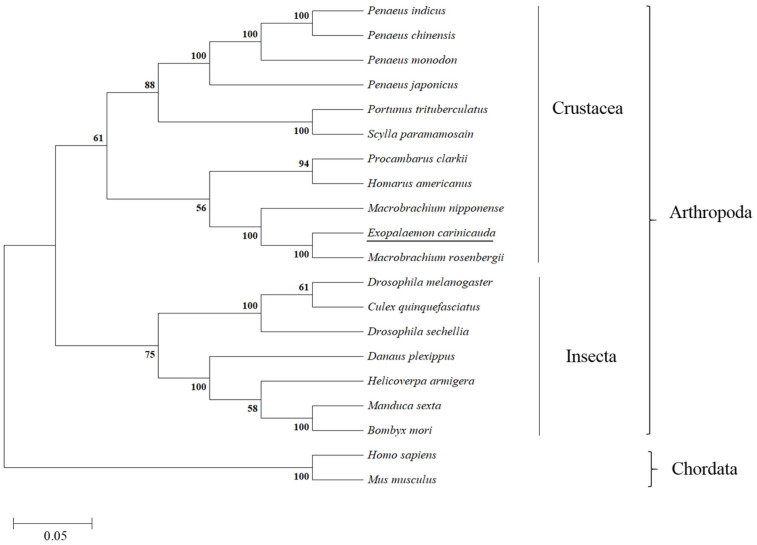 Figure 4
Figure 4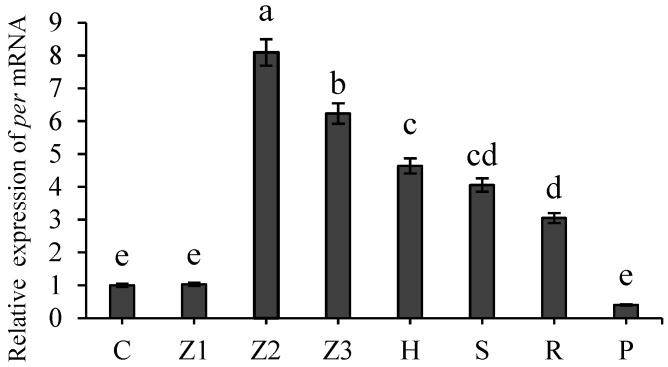 Figure 5
Figure 5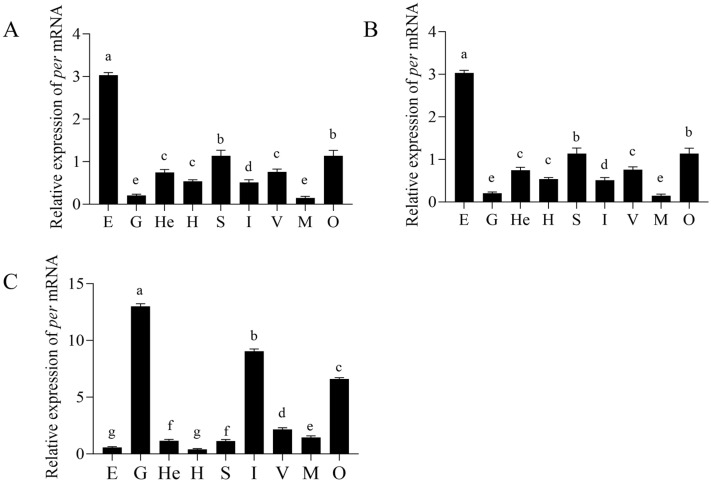 Figure 6
Figure 6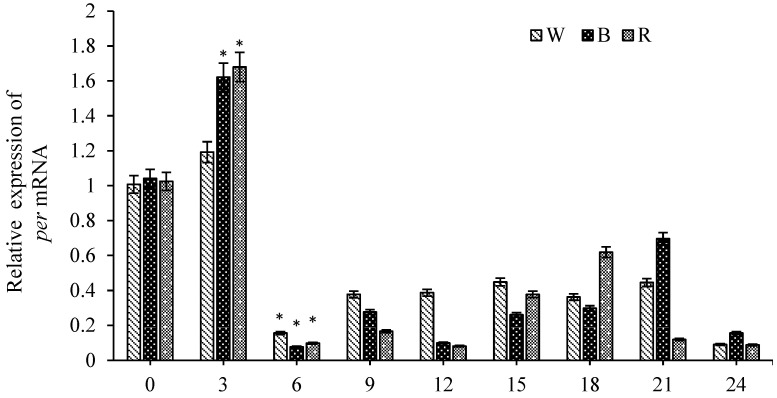 Figure 7
Figure 7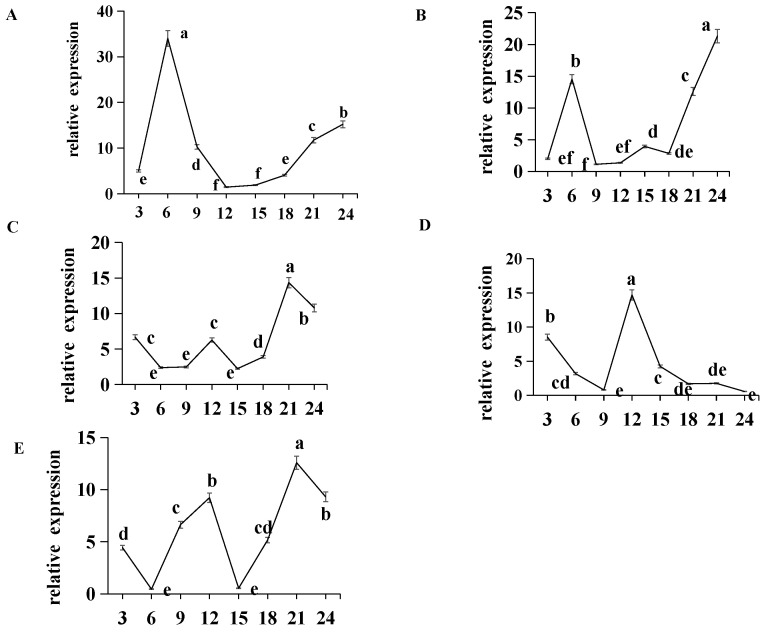 Figure 8
Figure 8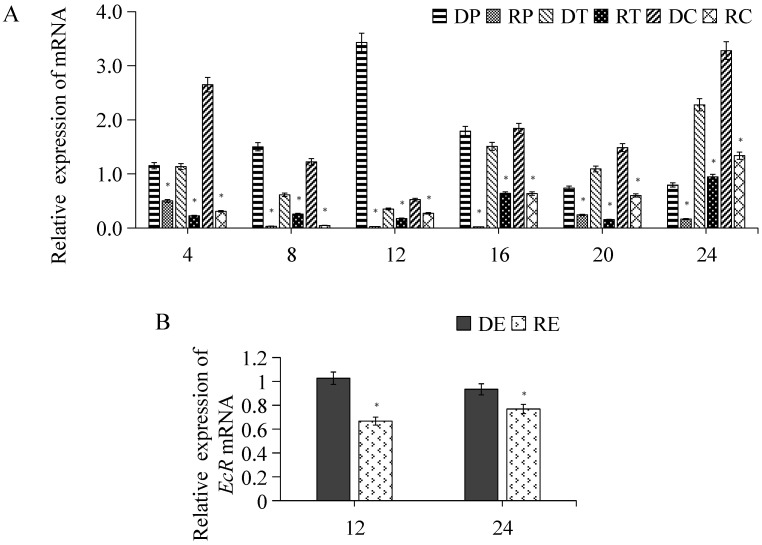 Figure 9
Figure 9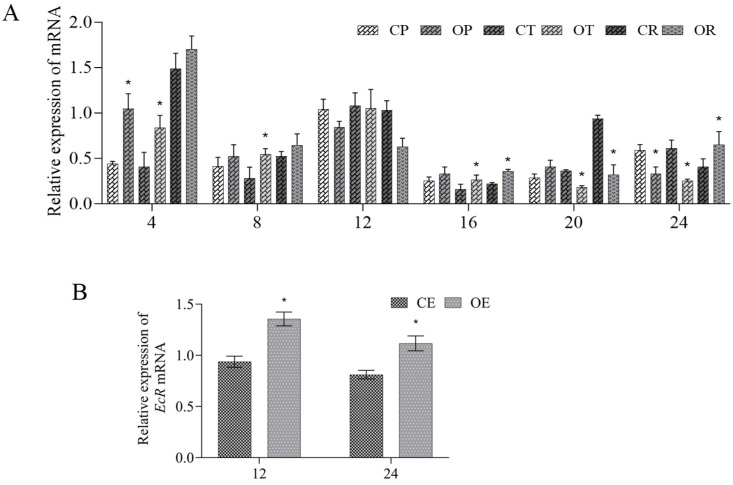 Figure 10
Figure 10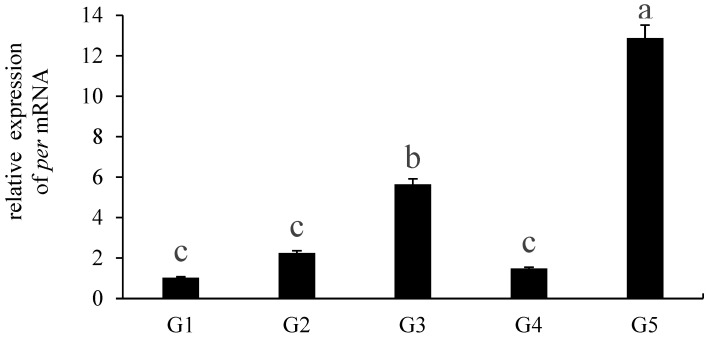 Figure 11
Figure 11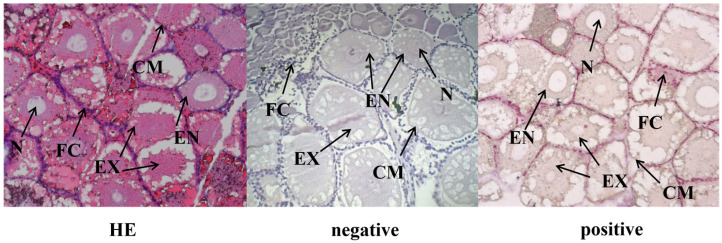 Figure 12
Figure 12- —Key R&D Program of Shandong Province
- —Excellent Teaching Team of Aquaculture Major under the “Qinglan Project” in Education Department of Jiangsu (2023)
- —“JBGS” Project of Seed Industry Revitalization in Jiangsu Province
- —Innovation and Entrepreneurship training program for college students of Jiangsu Ocean University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircadian rhythm and melatonin · Neurobiology and Insect Physiology Research · Developmental Biology and Gene Regulation
1. Introduction
Research on the molecular mechanisms of biological clock genes began with the breakthrough discovery of the Drosophila in the 1970s. In 1971, Konopka et al. first identified the period (per) gene by screening mutant Drosophila, demonstrating that its mutation led to the disruption of circadian rhythms and abnormal pupation cycles [1]. The gene was successfully cloned from Drosophila in 1984 [2], marking the beginning of the molecular era in circadian rhythm research. As a core component of the molecular oscillator of the biological clock, the per is widely expressed in the central nervous system and peripheral tissues of the organism [3,4,5]. It forms complexes with factors such as TIMELESS (TIM) and CRYPTOCHROME (CRY) to regulate key physiological processes, including circadian rhythm, cell cycle progression, reproductive behavior, and even tumorigenesis [6,7,8].
Circadian rhythms arise from the oscillatory expression of endogenous clock genes, which exhibit autonomous and plastic homeostatic regulation [9]. In addition to the core molecular oscillators encoded by genetics, exogenous timing factors (zeitgeber) include photoperiods, temperature fluctuations, feeding rhythms, and electromagnetic fields [1,10]. Among these, photoperiod is considered the most critical time factor: organisms integrate changes in spectral intensity/duration between day and night, synchronize physiological activities with the external environment [11]. In recent years, research on per has focused primarily on mammals and arthropods to elucidate the biological functions and regulatory mechanisms of circadian rhythms. In mammals, the suprachiasmatic nucleus (SCN) of the hypothalamus acts as the central pacemaker regulating the circadian system. The SCN, through the autonomic nervous system, directly or indirectly innervates the pineal gland and adrenal gland: it stimulates nocturnal melatonin secretion from the pineal gland and, via the hypothalamic–pituitary–adrenal (HPA) axis, induces a glucocorticoid peak preceding the active phase. These two hormones serve as key rhythmic signals that not only synchronize the peripheral biological clock but may also provide feedback regulation to the SCN [12]. Regarding the potential indirect effect of feeding behavior on ovarian development, studies indicate that circadian clock genes act as core regulators and rhythmic controllers of ovarian development [13,14]. The disruption of circadian genes leads to a 24 h weight loss under normal dietary conditions, thereby affecting the body’s nutrient and energy metabolism. Collectively, these mechanisms supply essential material and signaling substrates, further modulate the reproductive axis, and target genes within the ovary, ultimately influencing the process and quality of ovarian development. For instance, knockdown of per in mice (Mus musculus) leads to a loss of light-induced physiological rhythms and a shortened cell division cycle [11,15]. PERIOD (PER) forms positive and negative regulatory loops by interacting with CLOCK, brain muscle ARNT-like protein (BMAL), and TIM, finely tuning the oscillatory rhythm of the biological clock [16,17]. It also regulates circadian rhythms through trimer formation with CRY1 and TIM [18,19].
Exopalaemon carinicauda, a medium-sized marine economic shrimp endemic to China, has strong environmental adaptability and high reproductive ability with a short breeding cycle [20,21,22]. Females spawn annually from March to October. Nevertheless, the large-scale artificial seedling production for this species has not been successfully established, and the seedlings are still largely obtained from fertilized broodstock captured in natural waters [23,24]. In mammals and insects, studies have shown that circadian genes play a crucial role in reproduction [25,26,27]. Although research on clock genes in crustaceans has progressed in species such as Macrobrachium nipponense [28,29], Litopenaeus vannamei [6], and Euphausia superba [30,31], focusing on reproductive strategies in shrimp remains relatively limited. To explore the role of clock genes in the ovarian development of Exopalaemon carinicauda, this study cloned the per cDNA sequence and investigated the expression patterns of the per gene in E. carinicauda under varied light colors and photoperiods. Our results found that the per gene participates in regulating the circadian rhythm and positively affects ovarian development. These findings provide new theoretical insights into light-regulated reproductive regulation in crustaceans.
2. Materials and Methods
2.1. Ethics Statement
All experimental procedures in this study adhered to principles of laboratory animal care and use. Although E. carinicauda is an invertebrate of small size and not subject to review by institutional animal ethics committees, we still made efforts to minimize potential pain and stress throughout the experiments.
2.2. Experimental Materials
Healthy E. carinicauda from the self-breeding families of the research group were identified and selected for study. Prior to the experiments, shrimp (body length of 5.11 ± 0.41 cm, body weight of 2.10 ± 0.36 g) were acclimated for one week in 12 L:12 D in aerated tanks (50 × 40 × 30 cm) with a water temperature of 25 °C, salinity of 26, and pH of 8.0. Shrimp were fed daily at 08:00 and 18:00 with commercial feed equivalent to 3% of their body weight.
Samples of 24 individuals were collected at embryonic and larval stages, including: fertilized eggs, Zoea stages I–III, post-larval stage, adult stage, gonadal maturity stage, and postpartum recovery period. Furthermore, nine tissues from 9 shrimp were also collected, including: eyestalk, gill, heart, hepatopancreas, stomach, intestine, muscle, ovary, and ventral nerve cord at the stages of the adult, gonadal maturity, and postpartum recovery, following the method described in Ref. [32]. Ovarian tissues were further collected according to the five defined developmental stages [33]. All samples were taken with at least three biological replicates for each stage or tissue type (n ≥ 3).
2.3. Total RNA Extraction and cDNA Synthesis
Total RNA was extracted from all the samples using Trizol reagent (TransGen, Beijing, China) according to the manufacturer’s protocol. cDNA was synthesized using the HiScript III RT SuperMix for qPCR (+gDNA wiper) (Vazyme Biotech Co., Ltd., Nanjing, China), diluted for subsequent experiments, and then stored at −20 °C.
2.4. Cloning of Ec-per cDNA
The Ec-per cDNA sequence was obtained from the transcriptome database [34] of E. carinicauda with 5′/3′-RACE amplification kits (Takara, Beijing, China) along with gene-specific primers (as shown in Table 1). The RACE PCR was conducted following the manufacturer’s instructions of the Ex Taq kit (Takara).
2.5. Bioinformatics and Phylogenetic Analyses of Ec-per
The open reading frame (ORF) and encoded amino acid sequences of the PER were identified using ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html (accessed on 3 February 2026)). The theoretical isoelectric point and molecular weight were calculated by DNASTAR (V 17.5) software. The amino acid sequences of the PER from different species were multiple aligned. A phylogenetic tree was constructed with MEGA (version 11) software using the neighbor-joining method, with 1000 bootstrap replications [35].
2.6. qRT-PCR
The qRT-PCR primers used for the gene expression analysis are listed in Table 1. qPCR amplification was performed on a StepOne Plus system using SYBR Premix Ex Taq II kit (TaKaRa, San Jose, CA, USA) according to the manufacturer’s protocol, with 100 ng of cDNA from each sample as template. Relative expression levels were calculated using the 2^−ΔΔCt^ method [36] and are presented as mean ± standard deviation (SD) from at least three independent experiments. The selection of 18S rRNA as the reference gene was based on its demonstrated stability in prior validations performed by our research team [21,23,24] (p > 0.05). All statistical analyses were conducted with GraphPad Prism (version 10). Differences between the two groups were assessed using an independent-samples t-test, while comparisons among three or more groups were performed by Analysis of Variance (ANOVA) followed by Duncan’s multiple comparisons. p < 0.05 indicates a significant difference.
2.7. Experiments on Different Light Colors and Photoperiods
The experimental design comprised two parts: (1) Light-color experiment: A total of 180 shrimp were distributed into white, blue, and red light groups under continuous illumination (15 W, 1500 Lx). (2) Photoperiod experiment: 200 shrimps were randomly assigned to five light:dark ratio environments (0 L:24 D, 8 L:16 D, 12 L:12 D, 16 L:8 D, and 24 L:0 D). In both experiments, the eyestalks were taken at 0 h (the control group), 3 h, 6 h, 9 h, 12 h, 15 h, 18 h, 21 h, and 24 h. Each experiment included three parallel groups, and six replicate samples were taken at different time points.
2.8. RNA Interference
The siRNA primers of per (Table 1) were designed using the Thermo Fisher (Waltham, MA, USA) primer design online tool (http://rnaidesigner.thermofisher.com/rnaiexpress/design.do (accessed on 3 February 2026)), and were synthesized with TR102-T7 RNAi Transcription Kit (Vazyme, Nanjing, China) following the manufacturer’s instructions. A total of 200 shrimp were randomly assigned to the interference and control groups (three replicate groups/group). The siRNA (4 µg/g) or an equivalent volume of normal saline was injected into the pericardial cavity of shrimp in the respective groups. Eyestalk samples were collected at 4 h, 8 h, 12 h, 16 h, 20 h, and 24 h post-injection; ovaries tissues were also taken at 12 h and 24 h. Each time point had greater than three independent biological replicates.
2.9. Overexpression Experiment
According to the Ec-per core sequence, homologous recombination primers were designed using the CE Design software (V1.03) (as illustrated in Table 1). The pSPT18 plasmid was digested with BamHI and EcoRI, purified, and ligated with PCR-amplified and purified homologous recombination fragments. After successful sequencing (Sangon, Shanghai, China), the per mRNA for overexpression was transcribed in vitro using Easy Cap T7 Co-transcription Kit with CAG Trimer (Vazyme, China) according to the instructions. A total of 200 shrimp (each group includes 3 parallel groups) were injected into the pericardial cavity with per mRNA (4 µg/g) or an equal volume of normal saline, respectively. At least three duplicate samples were collected from different time points.
2.10. In Situ Hybridization (ISH)
According to the Ec-per core sequence, homologous recombination primers were designed using the CE Design software (V1.03). Probes primers for generating the SP6/T7 promoter for in situ hybridization (ISH) are listed in Table 1. The pSPT18 plasmid vector was digested with BamHI and EcoRI and then purified. After the preparation of homologous recombination primers for PCR amplification and purification, the recombination fragments were ligated into the vector. After successful sequencing (Sangon), the recombination vector was linearized with SP6/T7 probes primers and purified, and used as a template for mRNA probe synthesis using the Biotin RNA Labeling Kit (Beyotime, Shanghai, China) according to the manufacturer’s protocol. In vitro transcription with SP6 and T7 polymerases produced sense (negative control) and antisense (positive control) probes, respectively. Ovarian tissues from 20 individuals were fixed in 4% PFA for 24 h at 4 °C. The section of Hematoxylin–Eosin staining [37] was used for the control group. ISH was carried out with modifications to a previously described protocol [38]. Deparaffinized sections were incubated with 3% H_2_O_2_ for 10 min to remove endogenous peroxidase. Then it was digested with Proteinase K (20 ug/mL) at 37 °C for 10 min, prehybridized at 38 °C for 2–4 h, and hybridized with the Biotin-labeled (1:500) mRNA probe (1 ug/mL) against Ec-per mRNA overnight at 38 °C. Sense probes transcribed from the SP6 promoter served as the negative control, and probes targeting the T7 promoter were used as the positive control. Each group included three independent biological replicates. Finally, probe binding was visualized using DAB substrate (Beyotime, Shanghai, China).
3. Results
3.1. Cloning and Phylogenetic Analysis of the per cDNA
The per gene sequence has been submitted to the GenBank database (GenBank accession number PX552620). The Ec-per cDNA is 4611 bp in length, consisting of a 201 bp 5′-untranslated region and an 813 bp 3′-untranslated region. The ORF of per is 3597 bp, encoding a 1198 bp amino acid protein. The predicted molecular mass of 132.6 kDa, with a theoretical isoelectric point of 6.46 (Figure 1). The PER contains the two PAS (Period-Arnt-Single-minded) binding domains at 204279 AA and 340446 AA, respectively, and one PER domain at 1060~1178 AA. Phylogenetic analysis further reveals that the Ec-per has the closest phylogenetic relationship with Macrobrachium rosenbergii, followed by that with M. nipponense (Figure 2).
3.2. Expression Characteristics of Ec-per at Different Developmental Stages
The expression profile of Ec-per was further examined during the embryonic and larval stages. qPCR analysis revealed low transcript levels at stages of fertilized eggs and Zoea larvae I, with a significant peak observed at Zoea II (p < 0.05) (Figure 3). As development proceeded, the per expression declined markedly (p < 0.05), reaching its lowest level in the postpartum recovery period.
3.3. Tissue Distribution of Ec-per mRNA
The expression profiles of Ec-per across tissues were further analyzed during the different adult developmental stages. Ec-per expression varied significantly among tissues at each stage (Figure 4). At the adult shrimp stage, Ec-per transcripts were primarily enriched in the heart and intestine (p < 0.05), followed by the gill, stomach, and ventral cord nerves. Expression was relatively low in the hepatopancreas and muscle, and the lowest in the eyestalk and ovary. During gonadal maturation, the highest expression level was detected in the eyestalk (p < 0.05), while other tissues showed lower expression, with the gill and muscle exhibiting the lowest levels. In the postpartum recovery period, per mRNA accumulated most abundantly in the gill, intestine, and ovary, significantly higher than in other tissues (p < 0.05), whereas the hepatopancreas showed the lowest expression.
3.4. Ec-per Expression in Response to Different Light Colors and Photoperiods
The effect of different light colors on the expression of the per gene in E. carinicauda was studied (Figure 5). Following blue and red light exposures, per expression significantly increased, peaking at 3 h (p < 0.05), and subsequently exhibited low-amplitude oscillations under all three light conditions.
We further examined the expression pattern of per in the circadian rhythm of E. carinicauda. qPCR results indicated that Ec-per expression exhibited regular oscillations under various light-to-dark (L:D) ratios, including continuous darkness (0 L:24 D) (Figure 6A). Although the oscillatory rhythm of per remained relatively stable under constant darkness, both the amplitude and phase of oscillation shifted under varying photoperiods. Specifically, compared to the complete darkness, the oscillation phase shifted backward by 3 h under an 8 L:16 D photoperiod, and further delayed by 9 h under 16 L:8 D. Under constant light (24 L:0 D), the phase shift extended to 15 h (Figure 6B–E).
3.5. RNA Interference and Overexpression of Ec-per and Expression of Related Genes
After interfering with per expression, its mRNA level in the eyestalk was significantly reduced (p < 0.05) compared to the control group (interference efficiency >60%), confirming effective knockdown. Other core clock genes, including tim and cry1, also showed significantly decreased expression (Figure 7A). Further analysis of ovarian development revealed that the expression level of the Ecdysone receptor (EcR), which is involved in gonadal development, was also significantly lower than that of the control group at 12 h and 24 h post-interference (p <0.05) (Figure 7B).
After the overexpression of per mRNA, qPCR results indicated that the expression of per was significantly lower at 4 h (overexpression efficiency >60%, p < 0.05), confirming successful overexpression (Figure 8A). Interestingly, its expression subsequently declined relative to the control at 8 h, 12 h, and 16 h, before increasing again at 20 h and 24 h. Furthermore, the EcR mRNA level was significantly higher than that of the control group at 12 h and 24 h post-overexpression (p < 0.05) (Figure 8B).
3.6. Effect of per on the Ovarian Development of E. carinicauda
Expression characteristics of per were further examined across five ovarian developmental stages. Our results showed that per expression was significantly higher at the G3 and G5 stages than at other stages (p < 0.05), while no significant differences were observed at the G1, G2, and G4 stages (Figure 9). ISH was used to localize the per mRNA in the ovary of E. carinicauda (Figure 10). The strongest signal of per was detected in oocytes, including both exogenous yolk synthesis oocytes and endogenous yolk synthesizer oocytes, suggesting a potential role in the nutrient accumulation during oocyte maturation.
4. Discussion
4.1. The per Gene Exhibits Tissue-Specific and Developmental Stage-Dependent Characteristics
In this study, the per gene was first cloned from E. carinicauda. Sequence analysis revealed that the encoded PER protein contains two PAS domains and one PER domain. As critical structural motifs for the synergistic interaction of clock proteins [39,40,41], PAS domains are evolutionarily conserved across species [3,42] and have also been identified in zebrafish and mammals [43,44,45].
Expression analysis showed that Ec-per transcript levels were significantly higher during the Zoea II larval stage compared to the other developmental stages (p < 0.05), consistent with the findings in zebrafish [11], suggesting a conserved role for per in the larval development. Tissue-specific expression profiling in adult shrimp indicated high Ec-per expression in the heart and intestine, which may support the gonadal development by enhancing physical activity and energy metabolism. As the gonads mature, the per expression was the highest in the eyestalk. The eyestalk is a key organ for light perception and neurohormone secretion in crustaceans [40]. This expression pattern suggests that Ec-per may participate in regulating light-signal input to modulate circadian rhythms in peripheral tissues. During the postpartum recovery period, shrimp undergo physiological preparations for subsequent gonadal development and spawning. In response to factors such as organ aging, decreased environmental adaptability, and increased oxygen consumption [16], Ec-per expression was significantly upregulated in the gill, intestinal, and gonadal tissues, likely to meet energy requirements. Collectively, these results demonstrate that Ec-per exhibits both tissue-specific and developmental stage-dependent expression patterns.
4.2. Response of the per to Different Light Colors and Photoperiods
As a core component of the circadian clock genes, per plays a crucial regulatory role in the molecular oscillation system of biological rhythms and is involved in modulating the periodic rhythms of various biological activities [41,46]. Our findings indicate that per expression responds to white, blue, and red light, peaking at 3 h of light exposure. Following this peak, expression continued to oscillate, suggesting sensitivity to photic stimulation—a pattern similar to that observed for cry1 under the same light conditions [47]. In the later stages of E. carinicauda aquaculture, exposure to specific light colors (blue or red color) for 3 h (based on light color and RNA interference results) may promote gonadal development in shrimp. Previous studies have shown that CRY1 participates in circadian regulation by sensing blue and purple light signals [48], and that CRY1-PER products can form heterodimers that translocate into the nucleus to exert their function [49]. Blue light is thought to activate the CRY protein, which interacts with the PER-TIM heterodimer, leading to its degradation or functional inhibition, thereby resetting the light signal and synchronizing the circadian oscillator. This process ultimately influences the transcriptional rhythm of the per gene. Thus, we speculated that per may cooperate with cry1 in a signaling cascade to mediate responses to changes in external light color, though the precise mechanisms warrant further investigation.
To examine the photoperiod response of per, its expression profile was analyzed under various light conditions. Results showed that the per expression level exhibited regular oscillations under various light conditions, including continuous darkness. The pattern of alternating peaks and troughs was consistent with previous reports [50,51]. Light not only elicits acute behavioral and physiological responses but also drives phase shifts in the circadian clock. Altered photoperiods reduced amplitude and induced phase shift in the per expression rhythm, potentially due to light-mediated effects on the binding efficiency of the PER–TIM complex and the phosphorylation of PER protein. These changes could influence its nuclear translocation timing, thereby shifting the rhythmic phase [52,53]. Overall, the findings indicate per as an endogenous circadian clock gene whose expression remains largely independent of external light conditions; it may participate in regulating physiological rhythms through interactions with other clock genes.
4.3. Function of the per and Its Effect on Ovarian Development in E. carinicauda
After knocking down per expression, we found that the tim and cry1 genes were also significantly downregulated, indicating that per likely participates in a molecular feedback loop with these genes to synergistically regulate the circadian rhythms. In Drosophila, CLK and CYC heterodimers bind to the E-box (CACCTG), activating the transcription of core clock genes, per and tim, and driving circadian oscillations in the clock genes expression [54]. The resulting PER and TIM proteins form heterodimers that translocate into the nucleus and inhibit CLK/CYC activity, thereby repressing their own transcription in a negative feedback loop [55]. In mammals, the co-regulatory molecular mechanism mediated by the PER-TIM-CRY complex is highly conserved [56,57,58]. CLK/CYC and its mammalian homolog CLK/BMAL1 serve as central transcriptional activators in the circadian molecular network.
In M. nipponense, it was first reported that clock genes are involved in the ovarian development of crustaceans, revealing that the circadian function in the brain is closely related to ovarian development and female reproduction [28]. To further explore the role of per in ovarian development, its expression profiles were analyzed across five ovarian developmental stages. Our results indicated that per expression progressively increased prior to gonadal maturation, declined significantly after maturation, and peaked following ovulation. This expression pattern resembles that of the EcR gene in ovarian tissue [59], which has been reported to play a critical regulatory role in ovarian maturation [59]. Interfering with the per gene also reduced EcR mRNA levels, further supporting the involvement of per in regulating gonadal developmental cycles [59]. Beyond interactions between photoperiod-driven circadian rhythms and ovarian estrogen signaling, coordinated multi-system regulation is required for reproduction [60]. Since female shrimp rely on mating to complete spawning [61], we propose that the per gene integrates photoperiodic cues with endocrine pathways to regulate ovarian maturation rhythms and synchronize spawning events.
In D. melanogaster, core circadian clock genes (such as per, tim, Clk, and cyc) are crucial for reproductive fitness, and deficiencies in these genes can result in defective sperm incapable of fertilization [62]. This is likely due to disrupted circadian rhythms, which impair the energy and material supply necessary for yolk synthesis [63]. In our study, interference of the per gene downregulated reproductive-related genes, including EcR, potentially delaying ovarian development. We speculate that circadian clock genes do not directly regulate ovarian development processes or downstream hormone cascades, but instead act upstream by mediating photoperiodic timing [64]. The loss of per may thus disrupt circadian rhythmicity, leading to altered ecdysteroid secretion dynamics. This disruption may be through a transcriptional feedback loop involving coordinated regulation among these genes. Notably, even under short-day conditions in insects, RNAi-mediated per knockdown can promote ovarian development. ISH further revealed that the per gene could be detected in oocytes, including exogenous and endogenous yolk-synthesizing stages.
This study demonstrates rhythmic synchrony between the circadian gene per and key reproductive gene EcR in the ridgetail white prawn, offering key evidence for the coordination of biological clock and reproduction in crustaceans. While the observed correlation does not establish causation, EcR expression may be regulated by the circadian system or involved in reproductive feedback regulation, warranting further functional validation. The findings support the conservation of the “clock–endocrine–reproduction” axis across arthropods, while phase-specific expression and photoperiod sensitivity likely reflect habitat adaptation in shrimp. Comparative studies with other crustaceans could help distinguish conserved mechanisms from species-specific adaptations. In summary, this work outlines a molecular profile of circadian–reproductive coupling in the ridgetail white prawn, providing a foundation for mechanistic and cross-species studies, with implications for understanding crustacean reproductive adaptation and aquaculture management.
This study has certain limitations. First, the RNAi approach employed does not fully abrogate target gene function, and residual activity may lead to phenotypic effects weaker than those of a complete knockout. Second, while our findings indicate interactions among the per, tim, and cry genes, the precise molecular mechanisms linking these genes and their downstream pathways remain unclear. For instance, whether per directly regulates EcR to influence ovarian development or acts indirectly through the PER-TIM-CRY complex requires further investigation. Additionally, future studies should examine whether the disruption of per alters circadian rhythms in shrimp and further assess ovarian development status through gonadal indices and related gene expression profiles.
To address these limitations, future research studies should: (1) use gene editing technologies such as CRISPR/Cas9 to generate homozygous mutants [65,66,67] to validate and deepen current findings; (2) apply ChIP-seq to delineate the regulatory network of the PER–TIM–CRY protein complex and identify its potential downstream targets.
5. Conclusions
In this study, we first cloned the full-length cDNA sequences of the per gene from E. carinicauda. Our findings demonstrate that per plays a critical role in embryonic and larval development, with its tissue expression varying across developmental stages. Under different photoperiods, per exhibited phase shifts and altered oscillation amplitude, suggesting that the shrimp adjust their physiological activities in response to light cycles. Knockdown of the per gene resulted in significant downregulation of tim and cry1, as well as suppression of the EcR gene. ISH results further indicated that per is involved in oocyte proliferation and the accumulation of exogenous nutrients. Together, these results offer new insights into modulating circadian rhythms and reproduction in crustaceans through optimized light management.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Konopka R.J. Benzer S. Clock Mutants of Drosophila melanogaster Proc. Natl. Acad. Sci. USA 1971682112211610.1073/pnas.68.9.21125002428 PMC 389363 · doi ↗ · pubmed ↗
- 2Reddy P. Zehring W.A. Wheeler D.A. Pirrotta V. Hadfield C. Hall J.C. Rosbash M. Molecular analysis of the period locus in Drosophila melanogaster and identification of a transcript involved in biological rhythms Cell 19843870171010.1016/0092-8674(84)90265-46435882 · doi ↗ · pubmed ↗
- 3Gu Z. Liu Y. Dong Y. Zhang W. Yao H. Unravelling the neuroendocrine system of nocturnal spawning regulated by circadian clock in the razor clam, Sinonovacula constricta Aquac. Rep.20243810227810.1016/j.aqrep.2024.102278 · doi ↗
- 4Honma S. The mammalian circadian system: A hierarchical multi-oscillator structure for generating circadian rhythm J. Physiol. Sci.20186820721910.1007/s 12576-018-0597-529460036 PMC 10717972 · doi ↗ · pubmed ↗
- 5Yang M. Zhu X. Chen C. Guo X. Xu S. Xu Y. Du S. Xiao S. Mueller-Roeber B. Huang W. The plant circadian clock regulates autophagy rhythm through transcription factor LUX ARRHYTHMOJ. Integr. Plant Biol.2022642135214910.1111/jipb.1334335962716 · doi ↗ · pubmed ↗
- 6Fu Z. Han F. Huang K. Zhang J. Qin J.G. Chen L. Li E. Impact of imidacloprid exposure on the biochemical responses, transcriptome, gut microbiota and growth performance of the Pacific white shrimp Litopenaeus vannamei J. Hazard. Mater.202242412751310.1016/j.jhazmat.2021.12751334687996 · doi ↗ · pubmed ↗
- 7Lin C. Feng S. Deoliveira C.C. Crane B.R. Cryptochrome-Timeless structure reveals circadian clock timing mechanisms Nature 202361719419910.1038/s 41586-023-06009-437100907 PMC 11034853 · doi ↗ · pubmed ↗
- 8Zheng X.Z. Zhang Y.P. Zhu D.L. Geng Z.C. Molecular biology of the biological clock gene period Zool. Res.199865866