Anti-Leukemic Properties of Curcumin on Acute Lymphoblastic Leukemia: A Systematic Review
Teck Chee Soh, Ying Hui Tan, Pen Han Heng, Faizatul Isyraqiah, Rakesh Naidu, Kok-Lun Pang

TL;DR
Curcumin, a compound in turmeric, shows potential as a treatment for childhood leukemia by targeting cancer cells in multiple ways, but more research is needed to test it in humans.
Contribution
This review systematically summarizes curcumin's anti-leukemic mechanisms and highlights the need for improved bioavailability and clinical trials.
Findings
Curcumin inhibits ALL cell proliferation and induces apoptosis via DNA damage and mitochondrial dysfunction.
It targets key signaling pathways like JAK/STAT and PI3K/AKT and reduces drug resistance markers like MDR1.
In vivo evidence is limited, and no human trials have been conducted due to curcumin's poor bioavailability.
Abstract
Acute lymphoblastic leukemia (ALL) is the most common cancer in children, where immature white blood cells grow uncontrollably. While chemotherapy can treat it, side effects and resistance are common. Curcumin, a natural substance found in turmeric, has shown promising anticancer effects. This systematic review scientifically summarises the anti-leukemic effects of curcumin from cell lines, animal and human. Curcumin works through multiple ways, including damaging cancer cell DNA, causing stress inside the cells, blocking growth signals, and some additional mechanisms in causing suicidal cell death in ALL cells. However, studies in animals are very limited, and there is no solid evidence from human trials yet. One major challenge is that curcumin is not easily absorbed in the body, which makes it harder to use as a medicine. More research is needed to improve how curcumin is delivered…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
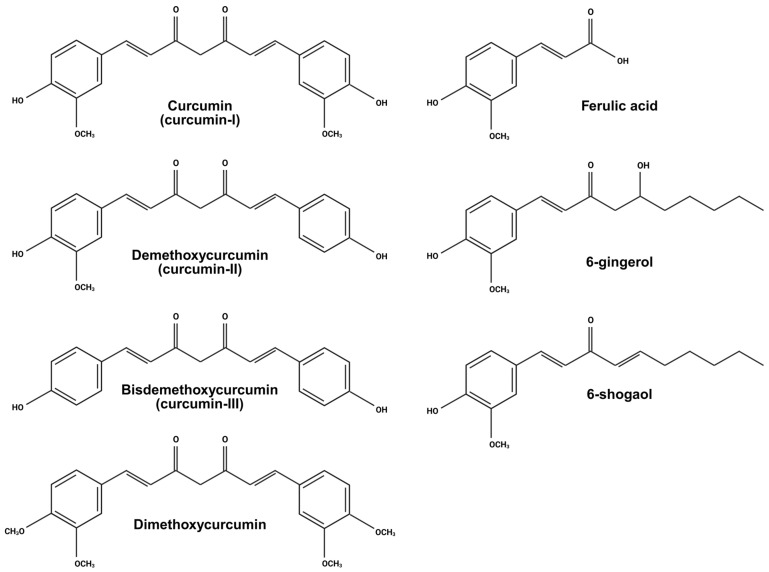 Figure 1
Figure 1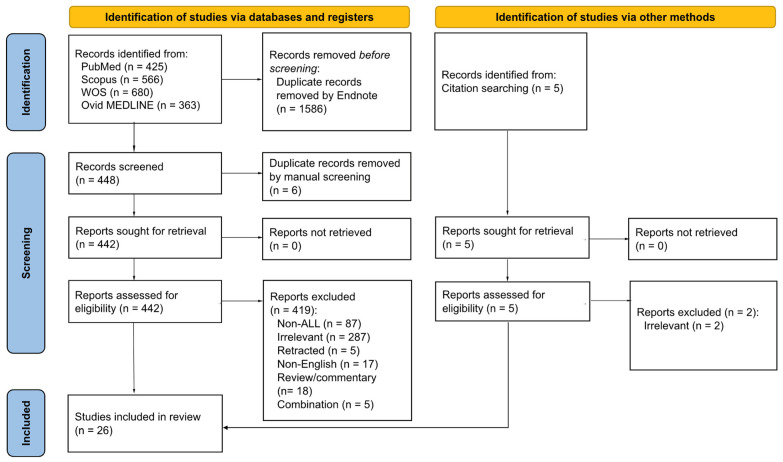 Figure 2
Figure 2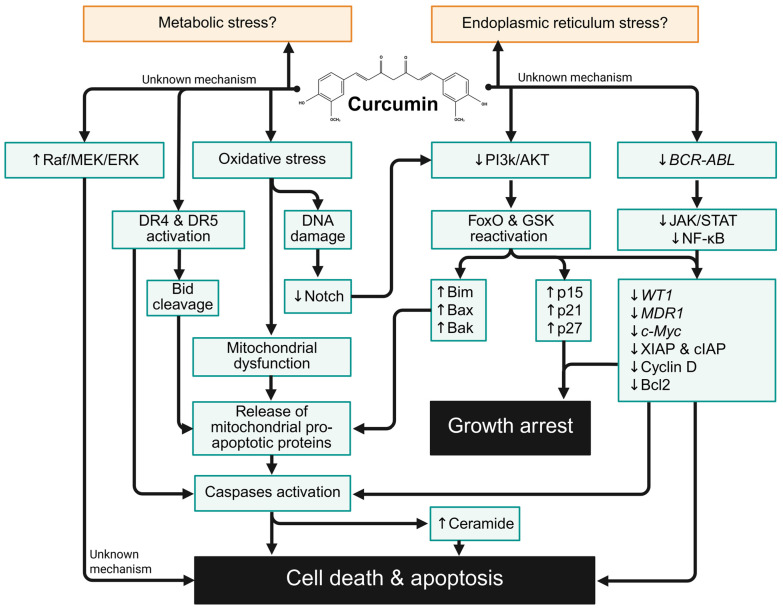 Figure 3
Figure 3- —School Seed Grant 2024
- —Jeffrey Cheah School of Medicine and Health Science and Monash University Malaysia Early Career Researcher Funding Scheme 2025
- —Monash University Malaysia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCurcumin's Biomedical Applications · Acute Lymphoblastic Leukemia research · Retinoids in leukemia and cellular processes
1. Introduction
Acute lymphoblastic leukemia (ALL), also known as acute lymphocytic leukemia, is a haematological malignancy that arises from the lymphoid cell lineage [1]. This disease results from genetic aberrations or mutations in the differentiation pathway of lymphoid progenitor cells, leading to uncontrolled proliferation and diminished apoptosis of immature lymphoblasts in the bone marrow [1]. Consequently, the high number of circulating blast cells replaces normal bone marrow cells and may infiltrate the peripheral blood circulation and other organs. According to the National Comprehensive Cancer Network, a diagnosis of ALL is made when lymphoblasts constitute ≥ 20% of nucleated bone marrow cells [2]. ALL exhibits a bimodal age distribution, with the first incidence peak in childhood and a second peak among adults older than 50 years [3,4]. According to the Global Burden of Disease 2023 [5], the age-standardised incidence of ALL was 1.71 per 100,000 population worldwide. In the United States (US), approximately 6100 new cases and 1400 deaths are projected for 2025 [3]. Based on the Surveillance, Epidemiology and End Results Program (SEER) data, the overall 5-year survival rate in the US is around 72.6% [3]. Children with ALL usually respond well to the standard treatment, with a 5-year survival rate of more than 90% [6]. However, the prognosis worsens with age, with 5-year survival rates of 74% for adolescents, 43–59% for adults, and below 30% for elderly patients [7].
Treatment for ALL involves a range of therapies, including systemic chemotherapy, intrathecal therapy, tyrosine kinase inhibitors (TKIs), allogeneic hematopoietic stem cell transplantation, targeted immunotherapy drugs, radiation therapy, and supportive care, tailored to the patient’s clinical condition [8]. Despite their efficacy, these treatments are commonly associated with undesirable and potentially life-threatening adverse effects, including diarrhoea, nausea and vomiting, neutropenia, anaemia, and neurotoxicity [2]. For instance, rapid destruction of leukemic cells may trigger tumour lysis syndrome [9]. In addition to gastrointestinal toxicities, chemotherapy and immunotherapy significantly increase the risk of infections, thrombosis, neuropathies, hepatotoxicity, anaphylaxis, cytokine release syndrome, and hypothalamic–pituitary–adrenal axis dysregulation [10]. Furthermore, radiotherapy is associated with long-term neurocognitive deficits, endocrinopathies, impaired growth, and secondary central nervous system malignancies [10]. Vulnerable populations, such as infants, adolescents, the elderly, and patients with Down Syndrome, experience higher risks of treatment-related toxicity and mortality [10]. Managing older ALL patients remains particularly challenging, as conventional treatments are often poorly tolerated and less effective [11]. Although treatment outcomes are favourable in younger patients, treatment-related toxicity frequently leads to noncompliance, contributing to relapse and the development of multidrug resistance (MDR). Relapse occurs in approximately 10% of children and up to 50% of adults who achieved complete remission [12]. While stem cell transplantation can provide durable remission or cure, it is associated with other risks, and identification of a suitable donor may be time-consuming [10]. These limitations highlight the urgent need for safer and more effective therapeutic strategies for ALL.
Natural compounds have long been investigated for their therapeutic potential against malignant diseases due to their broad spectrum of pharmacological activities. Curcumin or curcumin-I, scientifically known as diferuloylmethane, is a well-recognised bioactive compound derived from turmeric and exhibits diverse medicinal properties (Figure 1). Isolated from the rhizomes of the Curcuma longa plant, curcumin is a natural yellow pigment traditionally used in cooking, as well as for colouring, antiseptic, and cosmetic purposes. As a polyphenolic molecule, curcumin has been reported to possess anti-inflammatory, antioxidative, hypoglycaemic, antimicrobial, neuroprotective, and anticancer effects [13]. Accumulated evidence has demonstrated that it has potential efficacy against a variety of cancers, including gastric, skin, prostate, breast, colon, lung, ovarian, and liver cancers [14]. Curcumin is well-tolerated and generally safe for consumption at doses as high as 12 g/day [14]. Despite several related reviews on the anti-leukemic effects of curcumin being reported [15,16,17], to our best knowledge, a systematic review that summarises its anti-leukemic effects, especially on ALL, is not available. Therefore, this systematic review aims to summarise the anti-leukemic properties of curcumin based on in vitro, in vivo, and human studies.
2. Methodology
2.1. Literature Search
This systematic review was conducted in accordance with the Preferred Reporting Items for Systematic Review and Meta-analyses (PRISMA) 2020 guidelines and checklist (Supplementary Table S1) [18]. We searched the literature using four electronic databases, including PubMed, Web of Science (WoS), Scopus, and Ovid MEDLINE until August 2025 (Supplementary Table S2). The following keywords and Boolean operators were used: ((curcumin) OR (“natural yellow 3”) OR (diferuloylmethane) OR (“turmeric yellow”)) AND ((leukaem*) OR (leukem*)). The curcumin-related terms were selected based on PubMed Medical Subject Heading (MeSH) terms. Additionally, a manual literature search was conducted to identify additional records from the reference lists of included articles. The literature searching protocol was registered and publicly available on the Open Science Framework at https://osf.io/jdp64/ (accessed on 25 December 2025).
The records from all databases were first imported into EndNote 21 software as a library. Duplicates were removed using the software’s tool and manually reconfirmed. The remaining records were first screened by three reviewers independently for eligibility based on their titles and abstracts, followed by a full text review. Any discrepancies were resolved through discussions among all reviewers. Research articles reporting the anti-leukemic effects of curcumin, specifically on ALL, were included, regardless of whether the studies were in vitro, in vivo, or human studies. There were no restrictions on the year of publication. However, we excluded studies that were: (1) written in a non-English language; (2) lacked primary or full data (e.g., review articles, meta-analyses, reports, books, book chapters, commentary articles, and conference/proceeding papers); (3) published as a preprint; (4) retracted; or (5) focused on curcumin analogues, Curcuma extracts, or combinations with other non-specific bioactive compounds, without curcumin data. After finalising the articles, three reviewers extracted essential information from the included articles and summarised it in Table 1 independently, which includes authors’ names, publication years, ALL models, curcumin details, treatment conditions, findings, and critical appraisal score. A PRISMA flow chart summarising our procedures for article identification, screening, and final inclusion of articles is presented in Figure 2.
2.2. Critical Appraisal
Two reviewers independently assessed the quality of the included articles using a modified OHAT risk of bias tool for in vitro studies and the OHAT risk of bias tool for animal studies [47,48,49]. We evaluated each domain accordingly, with one question per domain. Full compliance and direct evidence of a low risk of bias in one domain were recorded as “Definitely low risk of bias (+ +)”. Mild deviations or indirect evidence of a low risk were marked as “Probably low risk of bias (+)”. Insufficient information on relevant risk of bias or indirect evidence of a high risk of bias was recorded as “Probably high risk (−)”, and lastly, “Definitely high risk (− −)” was used for direct evidence of high-risk bias practices. Overall appraisal utilised a 3-level tiering of quality according to previous studies [48,50]. Studies classified as Tier 1 were considered high-quality articles with >50% of Definitely and/or Probably Low Risk; Tier 2 as moderate quality; while Tier 3 were poor-quality articles with >50% of Definitely and/or Probably High Risk. Any disagreement between authors was resolved by discussion. The OHAT tier is included in Table 1, and detailed information is shown in Supplementary Tables S3 and S4.
3. Results
3.1. Characteristics of Included Articles
Our literature search across four databases retrieved 2034 articles: 425 records from PubMed, 566 records from Scopus, 680 records from WoS, and 363 records from Ovid MEDLINE. After removing duplicates using EndNote software and manual check, 442 records were selected for further screening and full-text eligibility assessment. Of these, 419 records were excluded from the final analysis for the following reasons: irrelevant content (n = 287), non-ALL studies (n = 87), retracted articles (n = 5), non-English articles (n = 17), review or commentary articles (n = 18), and combinational studies lacking anti-leukemic data specific to curcumin (n = 5). A total of 26 articles were included to be relevant in this systematic review, including 3 records identified through reference searching.
In this systematic review, most of the included studies used immortalised T lymphoid ALL-derived cell lines, including Jurkat cells [19,20,21,24,30,31,32,40], CCRF-CEM cells [20,24,26,29,30,34,38,45], MOLT-4 cells [24,27,32,41,44], HSB2 cells [24], KOPT-K1 [42], DND-41 [42], and TALL-1 cells [42]. Despite B lymphoid ALL being the major subtype of ALL in both children and adults [51], relevant studies on B-ALL in this systematic review were far fewer than T-ALL. The immortalised B-ALL cell lines include human childhood REH cells [32,35,36,39], childhood SUP-B15 cells [37,39,45], childhood 697 cells [39], and adult RS4;11 cells [35,39]. Doxorubicin-resistant CCRF-CEM cells with overexpressed P-glycoprotein (P-gp; also known as MDR1) were also being studied [38]. Virus-transformed models such as human EBV-related Burkitt lymphoma lymphoblast-like Raji cells [33] and HTLV-1 transformed T cell leukemia cell lines (MT-2, HuT-102 and SLB-1 cells) [25] were also studied. Only four studies used primary patient-derived ALL cells isolated from the peripheral blood [41] or bone marrow [22,23,45] for ex vivo short-term exposure to curcumin. Nevertheless, the cell concentration used in treatment varied within the same studies and across studies. The majority of the included studies were inconsistent in treatment conditions [19,29,32,33,41] or did not disclose this information for some or all experiments [21,22,23,24,26,27,34,35,37,39,40,42,45]. Some studies used cell number instead of cell concentration [20,24,26,29,30,32,33,35,36,38,41,45]. Rajasingh et al. and Koszałka et al. consistently used 2.5 and 3 × 10^5^ cells/mL in their studies [25,44]. Only two studies used ALL xenograft animal models, which included BALB/c null mice with intravenous injection of 10^7^ human B-ALL SUP-B15 cells [45] and NOD/SCID mice with intravenous injection of 5 × 10^6^ human B-ALL SEM cells [46]. To date, there have been no reports of curcumin treatment in patients with ALL.
Most studies sourced curcumin from Sigma [20,21,22,24,27,29,30,32,34,35,36,37,39,40,44,45], though some were obtained from Merck [19], Invitrogen^TM^ Life Technology [23], Calbiochem [25], Cayman Chemical [31,46], Acros Organics [33], and Fluka AG [38]. Two studies purified [27] or synthesised curcumin by themselves [42]. Two studies did not disclose the source or manufacturer of curcumin [26,41]. Anuchapreeda et al. [22,23,27], Kong et al. [29], Olivas-Aguirre et al. [40], Koszałka et al. [44], and Zunino et al. [46] stated the purity of curcumin used, which ranged from ≥65% to 99%. Anuchapreeda reported that commercial-grade and self-extracted curcumin from turmeric powder contain 77% curcumin (or curcumin-I), 17% demethoxycurcumin (or curcumin-II), and 3% bisdemethoxycurcumin (or curcumin-III) [22,23,27]. Piwocka et al. and Khar et al. dissolved curcumin in ethanol [19,20], and most studies used dimethyl sulfoxide to dissolve curcumin [29,30,31,34,35,37,39,41,44,45,46]. The others did not disclose how they dissolved curcumin. In addition, some studies disclosed the maximum concentration of vehicle used in treatment [20,29,31,39,40,44], the concentration of stock solution [29,30,34,37,40,41,44,45], and/or storage conditions for stock solution [29,30,37,41,45,46]. For in vitro studies, the tested concentrations of curcumin were up to 100 μM for treatment durations ranging from 1 h to 7 days. Some studies did not disclose the treatment duration or concentrations [25,26,36,38]. For animal studies, 0.5% w/w curcumin in the diet for 3 weeks was used with an oral route of administration [46], while intraperitoneal injection of curcumin (either 5 [46] or 50 mg/kg body weight/day [45]) for 2 to 4 weeks was employed.
Based on the OHAT critical appraisal tool, in vitro studies are inherently low risk in selection bias, performance bias, attrition/exclusion bias, and detection bias [50]. Nevertheless, all the included studies did not disclose blinding, even though it is not a common practice in reporting. The major issue is exposure characterisation, where most of the studies did not disclose the source [26,42], purity of curcumin [19,20,21,24,25,30,31,32,33,34,35,36,37,38,39,45], curcumin treatment duration, and tested concentrations [25,26,36,38,40] or leukemic cell concentration in treatment [25,26,36,38,40]. More than half of the included studies were not consistent with the treatment conditions [19,29,32,33,41] or did not disclose this information for some or all parameters [21,22,23,24,26,27,34,35,37,39,40,42,45]. Despite some quality issues, based on the tier grouping, all the included studies were ranked as Tier 1 because more than 50% of domains were classified as Definitely and/or Probably low risk of bias [20,22].
3.2. Curcumin Induces Cytotoxicity, Apoptosis, Autophagy, and Growth Arrest in ALL Cells While Sparing Non-Cancerous Cells
Curcumin was consistently reported to induce time- and/or concentration-dependent cytotoxicity and apoptosis in ALL cells [19,20,21,24,25,26,27,30,31,32,35,36,37,38,39,40,42,44,45]. Curcumin-treated cells demonstrated typical apoptotic features, including morphological alterations, DNA fragmentation, poly(ADP-ribose) polymerase (PARP) cleavage, mitochondrial dysfunction, cytochrome c release, BH3-interacting domain death agonist (Bid) cleavage, and downstream executioner caspases activation [19,20,21,24,25,30,32,35,36,39,40,42]. It also suppressed the colony-forming capacity of Bcr-Abl-positive SUP-B15 and RS4;11 cells [39], induced autophagy and autophagy-related cell death in SUP-B15 cells [37], and caused antiproliferation and growth arrest [25]. Curcumin induced G2/M arrest in CCRF-CEM, RS4;11 cells, REH cells [34,35], Jurkat T, and MOLT-4 cells [30] but G1 arrest in human EBV-related Burkitt lymphoma lymphoblast-like Raji cells [33] via upregulation of CDKN2B gene and downregulation of cyclin D1 and c-Myc. Despite these findings, the precise mechanisms underlying curcumin-mediated cell cycle arrest in ALL cells remain unclear.
Overall, the current evidence suggests that curcumin is more cytotoxic against the T-lineage of ALL cells than the B-lineage, with Jurkat T cells showing the greatest sensitivity. The reported IC_50_ values of 24 h curcumin treatment ranged from 4.22 μM and 36.5 μM in Jurkat T cells [32,40], decreasing to 2.89 μM upon 48 h treatment [32]. Similar time-dependent cytotoxicity of curcumin was observed in B-ALL REH (24 h IC_50_ of 21.81 μM; 48 h IC_50_ of 18.62 μM) and T-ALL MOLT-4 cells (24 h IC_50_ of 37.27 μM; 48 h IC_50_ of 23.72 μM) [32]. In T-ALL CCRF-CEM cells, the IC_50_ values were 6.49 μM [38], 8.68 μM [29], and 9.84 μM (unconfirmed treatment time) [26] upon 48 h treatment and reached 32.78 μM upon 72 h treatment [45]. Curcumin also induced cytotoxicity to other T-lineage ALL cells including doxorubicin-resistant CCRF-CEM cells (IC_50_: 21.04 µM) [38], B-ALL Raji cells (IC_50_: 20 µM) [33], T-ALL KOPT-K1 cells (IC_50_: 8.22 µM), T-ALL DND-41 cells (IC_50_: 13.255 µM), and TALL-1 cells (IC_50_: 6.33 µM) [42] upon 48 h treatment. It was also cytotoxic, but to a lesser degree, in B-ALL SUP-B15 cells, with IC_50_ values of 27.59–30.59 µM upon 72 h treatment [37,45].
Consistent with other cancer models, curcumin was selectively targeting ALL cells while sparing non-cancerous cells at the same tested concentrations. It was non-cytotoxic to non-cancerous cells, including rat skin fibroblasts, Chinese Hamster Ovarian CHO cells, mouse fibroblast L-929 cells, rat embryo fibroblast F111 cells, primary lymphocytes and hepatocytes (unknown origin, potentially rodent) [20], African green monkey kidney epithelial Vero cells [29], human corneal epithelial cells [20], and primary PBMCs from healthy donors [32,45]. However, Korwek et al. reported that curcumin induced apoptosis equally in both Jurkat T leukemic cells and primary T cells isolated from healthy donors [31].
3.3. Curcumin Activates Both Intrinsic and Extrinsic Apoptosis Pathways and Inhibits Oncogenic Signalling in ALL Cells
In in vitro ALL models, curcumin activates both intrinsic and extrinsic pathways of apoptosis, depending on cell type and experimental conditions. It was demonstrated to activate both caspase-8 and -9 in ALL cells with the decrease in procaspase and/or increase in cleaved caspases [21,24,39]. Specifically, curcumin induced the intrinsic pathway of apoptosis with the involvement of mitochondrial dysfunction, cytochrome c release, altered B cell lymphoma-2 (Bcl2)-associated X (Bax) to Bcl2 ratio, upregulation of mitochondrial pro-apoptotic proteins, and caspase-9 activation [19,21,24,32,39,45]. It also downregulated the inhibitors of caspases, including X-linked inhibitor of apoptosis protein (XIAP), cellular inhibitor of apoptosis protein (cIAP), and survivin [24,30,39], thus enhancing the susceptibility of leukemic cells to the intrinsic pathway of apoptosis. Mitochondrial membrane potential loss was observed in CCRF-CEM, HSB2, Jurkat T, and MOLT-4 cells upon 24 h treatment of 40 µM curcumin [24]. A higher concentration of curcumin (70 µM) caused mitochondrial membrane potential loss in Jurkat T cells as early as 4 h [40], while a lower concentration of 10 µM caused similar effects by 6 h [32]. Olicas-Aguirre et al. demonstrated that curcumin-induced mitochondrial dysfunction may result from mitochondrial uncoupling rather than mitochondrial calcium overload [40]. On the other hand, curcumin also activates the extrinsic pathway of apoptosis in ALL cells. It upregulated death receptors DR4 and DR5, induced Bid cleavage, and activated caspase-8 [21,24,31,41]. A non-toxic concentration of curcumin enhanced tumour necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL)-induced cytotoxicity and apoptosis in MOLT-4 cells and primary ALL cells from patients with B- and T-ALL [41]. The precise mechanism by which curcumin triggers mitochondrial dysfunction, releases mitochondrial proapoptotic proteins, and activates caspase-8 and 9 in ALL cells remains unclear.
Caspase activation in curcumin-treated ALL cells was found to be essential by using the pan-caspase inhibitor, z-VAD-FMK [24,30,32,39]. Pretreatment of z-VAD-FMK completely abrogated curcumin-mediated apoptosis, caspase-3 activation, and/or PARP cleavage in CCRF-CEM, Jurkat T, MOLT-4 cells [24,30], as well as in RS4;11 and SUP-B15 cells [39]. Gopal et al. demonstrated that caspase inhibition significantly but not completely protected Jurkat T cells from curcumin-induced cytotoxicity [32]. The IC_50_ of curcumin increased from 4.22 µM to 15 µM with z-VAD-FMK pretreatment [32]. Piwocka et al. demonstrated that a lower concentration and shorter pretreatment period of z-VAD-FMK (10 µM; 30 min) prevented ultraviolet-induced karyorrhexis but not curcumin-induced chromatin condensation in Jurkat T cells [19], suggesting suboptimal blocking of caspases. Curcumin-induced chromatin condensation occurred without caspase-3 activation, DNA fragmentation, and mitochondrial membrane potential loss [19], indicating caspase-independent cell death under short-term exposure to curcumin.
Curcumin exerts anti-leukemic effects in ALL cells by suppressing Janus kinase (JAK)/signal transducer and activator of transcription (STAT) [25,37,45] and phosphoinositol-3 kinase (PI3K)/AKT, followed by functional reactivation of Forkhead box O (FoxO)/glycogen synthase kinase 3β (GSK3β) [24,30,37,39,45] while activating the RAF/mitogen-activated protein kinase kinase (MEK)/extracellular signal-regulated kinase (ERK) pathway [37]. The PI3K/AKT and JAK/STAT pathway signalling cascades are highly associated with the leukemogenesis of ALL [52,53]. Curcumin induced dephosphorylation of AKT (inactivation), reducing downstream phosphorylation of FoxO and GSK3 (resulting as functional activation) in CCRF-CEM, Jurkat T and MOLT-4 cells [24,30,45], with similar effects observed in SUP-B15 and RS4;11 cells [39,45]. Guo et al. reported that curcumin suppressed the activation of AKT/mechanistic target of rapamycin (mTOR) but not phosphatase and Tensin homolog (pTEN) and 3-phosphoinositide-dependent protein kinase 1 (PDK1) in SUP-B15 cells and/or primary ALL cells isolated from the bone marrow of patients [37,45]. Curcumin also suppressed STAT5 activation in SUP-B15 cells in a time and concentration-dependent manner [45]. Concurrently, early and prolonged phosphorylation of RAF, MEK1/2, and ERK 1/2 was observed in curcumin-treated SUP-B15 cells, contributing to cell death rather than proliferation [37]. Further, chemical inhibition of MEK (by U0126) protected SUP-B15 cells from curcumin-mediated cytotoxicity and autophagy [37]. Similarly, it inhibited phosphorylation of JAK3, non-receptor tyrosine kinase 2 (TYK2), STAT3, and STAT5 in HTLV-1 transformed leukemic MT-2, HuT-102, and SLB-1 cells [25]. Curcumin also downregulated BCR-ABL mRNA in SUP-B15 cells [45] and inhibited nuclear factor κB (NF-κB) activation in REH cell apoptosis [36].
3.4. Curcumin Induces Oxidative Stress, DNA Damage, and Ceramide Accumulation as Upstream Apoptotic Signals
Curcumin exhibits mixed findings of oxidative stress and DNA damage induction in ALL cells, potentially depending on cell type and treatment conditions. Gopal et al. reported that curcumin induced reactive oxygen species (ROS) production and intracellular glutathione (GSH) depletion in REH, Jurkat T, and MOLT-4 cells, which could be prevented by N-acetyl cysteine (NAC) and GSH supplementation [32]. Similarly, curcumin induced ROS generation and chromosomal breaks in Raji cells [33], and it induced DNA damage with impaired DNA repair and Notch pathway suppression in KOPT-K1, DND41, and TALL-1 cells [42]. Parallelly, it caused ROS production, GSH depletion, DNA damage, and/or H2Ax (DNA damage sensing protein) activation in SUP-B15 and RS4;11 cells [39], as well as Jurkat T and MOLT-4 cells [30]. In contrast, Korwek et al. observed that 24 h curcumin exposure (up to 50 µM) did not induce significant DNA damage or DNA repair with no activation of H2Ax, ataxia-telangiectasia mutated protein (ATM), checkpoint kinase 2, and P53 in Jurkat T cells [31]. Similarly, Kong et al. also reported that curcumin (up to 200 µM) was not damaging pBR322 DNA plasmids in a cell-free system, unless in the presence of copper (II) ions (20 µM, a physiologically achievable concentration in serum) [29].
Interestingly, curcumin induced ceramide accumulation in ALL cell apoptosis, which is related to caspase activation and oxidative stress. Curcumin accumulated ceramide with a decrease in sphingomyelin synthase (SMS) activity but not ceramidase, sphingomyelinase, or glucosylceramide synthase [30]. Exogenous GSH prevented curcumin-induced apoptosis, SMS suppression, and ceramide production. Pan-caspase inhibition by z-VAD-FMK, as expected, could not block GSH depletion but surprisingly prevented curcumin-induced ceramide generation [30]. Pretreatment with buthionine sulphoximine (BSO; a GSH synthesis inhibitor) or D-609 (a SMS inhibitor) enhanced curcumin-mediated ceramide production and apoptosis [30].
3.5. Curcumin Downregulates WT1 and MDR1 and Upregulates CDK2NB Gene Expression
Curcumin downregulates Wilms’ tumour 1 (WT1) and multidrug resistance 1 (MDR1) gene expressions, which may contribute to its cytotoxicity in ALL cells. It reduced WT1 mRNA expression in MOLT-4 cells [27] and primary ALL cells isolated from patient bone marrow [22], with greater effects in cells exhibiting high to moderate expression of WT1 [22]. Interestingly, pure curcumin (95 to 99% purity), but not self-prepared or commercial-grade curcuminoid mixture (77% purity), significantly decreased WT1 mRNA and protein levels in MOLT-4 cells [27]. Similarly, curcumin also downregulated MDR1 in primary ALL cells isolated from patient bone marrow [23], with the strongest effects in cells with high to moderate MDR1 expression. In other words, it was most effective in relapsed patients (60%), followed by newly diagnosed (56%), drug maintenance (50%), and completed treatment cases (43%) [23]. Parallel to this, it also significantly inhibited MDR1 (also known as P-gp) activity in doxorubicin-resistant, MDR1-overexpressed CCRF-CEM cells [38]. Nevertheless, the molecular mechanisms by which curcumin regulates these gene expression remain unclear.
Curcumin upregulated tumour suppressor gene in ALL cells, especially p15INK4b, a cyclin-dependent kinase inhibitor, partly through epigenetic regulation. CDKN2B (gene name for p15INK4b) is commonly hypermethylated in ALL [54,55]. A 72 h treatment of curcumin (5 and 10 µM) significantly induced G1 arrest in Raji cells via CDKN2B mRNA upregulation by reversing promoter hypermethylation and downregulating DNA methyltransferase 1 (DNMT1) mRNA [33]. At lower concentrations of 1 and 2 µM, it did not significantly upregulate CDKN2B and CDH1 genes in CCRF-CEM cells with no apoptosis induction, despite an increasing trend in CDKN2B mRNA being observed [34]. Similarly, DNA pyrosequencing confirmed that curcumin did not significantly reverse DNA methylation at CDKN2B CpG sites under these low-concentration treatments [34]. Despite the lack of DNA hypomethylating activity, curcumin significantly upregulated Ten Eleven Translocation (TET1/2/3) enzymes concurrently, but not DNMT1/3a/3b [34]. Further studies are needed to confirm the mechanisms by which curcumin regulates DNA hypomethylation and gene expression in higher concentrations.
3.6. Curcumin Exhibits Limited Efficacy in ALL Xenograft Models
Preclinical in vivo studies of curcumin on ALL are scarce and show limited efficacy. Zunino et al. reported that oral supplementation (0.5% in diet for 3 weeks) and intraperitoneal injection (5 mg/kg body weight/day for 4 weeks) of curcumin did not reduce SEM leukemia cell growth in the blood of NOD/SCID mice [46]. Higher doses of curcumin injection (25–100 mg/kg body weight) were toxic and reduced the lifespan of mice; thus, Zunino et al. used a dose of 5 mg/kg that was not toxic. Nevertheless, curcumin, whether administered orally or intraperitoneally, did not improve the survival of mice compared to untreated controls [46]. In contrast, Guo et al. reported that a high dose of intraperitoneal curcumin (50 mg/kg body weight/day for 14 days) reduced SUP-B15 leukemic cell infiltration in the spleen and BCR-ABL mRNA in the bone marrow of immunosuppressed female BALB/c null mice [45]. Nevertheless, the safety profile of curcumin was not reported [45]. There is no report on the therapeutic effects of curcumin in patients with ALL.
4. Discussion
This systematic review highlights the anti-leukemic effects of curcumin in ALL, based on 26 selected in vitro and in vivo studies. Across studies, curcumin reduced ALL cell viability in a concentration- and time-dependent manner, mainly through intrinsic and extrinsic pathways of apoptosis with suppression of proliferative signalling pathways including PI3K/AKT and JAK/STAT, although some experimental variations lead to conflicting findings. For instance, Piwocka et al. reported that curcumin induced Jurkat T cell death, which is independent of DNA fragmentation, mitochondrial depolarisation, and caspase-3 activation [19]. This discrepancy might be due to the experimental design of transient exposure to curcumin (1 h treatment) followed by a 2 to 5 h curcumin-free recovery period. Most other studies assessed the cell death outcomes after 24 to 48 h of treatment. In addition, Piwocka et al. also defined cell death based on chromatin condensation from morphology observation [19], which is not always indicative of apoptosis [56]. Short and low concentration of z-VAD-FMK (10 µM; 30 min) in their study did not prevent curcumin-induced chromatin [19], while other studies used ≥50 µM and ≥1 h of z-VAD-FMK pretreatment [24,30,32,39]. Incomplete or suboptimal inhibition of caspases could account for these observed differences. In another study, Kong et al. demonstrated that curcumin did not cause DNA damage in a cell-free system unless in the presence of copper ions [32]. It is noted that copper ions can be present in cell culture media upon the enrichment of fetal bovine serum. On the other hand, Korwek et al. observed selective caspase-8 activation without detectable caspase-2 and -9 cleavage in curcumin-treated Jurkat T cells [31]. It is known that a variety of antibodies are available to evaluate caspase expression, either pro-, cleaved, or both forms [57,58]. The authors did not disclose the catalogue number of the antibodies used; hence, we were unable to confirm this. Band intensity analysis, however, indicated activation of procaspase-2 and -9, where the intensities of procaspase-2 and -9 were decreased along with curcumin treatment [31]. They also reported non-selective apoptosis induction in both Jurkat T cells and primary T lymphocytes from healthy donors, which may reflect biological and culture-related differences rather than experimental variation. Culture media [59,60,61] and serum batch-to-batch variation [62] are known to influence T lymphocyte metabolism and function. Nevertheless, the culture conditions for the isolated T lymphocytes were not reported.
Curcumin induces apoptosis in ALL cells primarily through oxidative stress and DNA damage, although the upstream molecular mechanisms remain unclear. Curcumin induced ROS generation and intracellular GSH depletion, leading to oxidative DNA damage and mitochondrial dysfunction [30,33,39,42]. This is consistent with previous findings where curcumin activated caspase-2 and/or -9 in ALL cells, which are known to be activated in DNA damage-mediated cell death [63,64]. At low concentrations, curcumin acts as an antioxidant, whereas at high concentrations, it behaves as a pro-oxidant in cancer cells [65]. In non-ALL models, curcumin has been shown to inhibit thioredoxin reductase and convert it to a NADPH oxidase to generate ROS [66,67,68]. Curcumin also induced oxidative stress through GSH depletion, which diminishes cellular defence against ROS. However, it is uncertain whether GSH depletion is mainly due to ROS scavenging or direct Michael addition with curcumin itself. GSH-conjugated curcumin has been detected in cell-free systems [69,70] and in human colorectal cancer Caco-2 cells [70]. In addition, NAC, which is membrane-permeable, can reduce intracellular radicals, whereas GSH cannot readily enter cells without the aid of specific membrane transporters [71,72,73]. Radicals like superoxide anion and hydroxyl radicals are highly reactive and are more likely to cause damage in situ rather than diffuse across the membrane to be neutralised by extracellular GSH. This explains why NAC and GSH supplementation, but not superoxide anion dismutase and catalase, prevented curcumin-induced GSH depletion in ALL cells [32]. We postulate that curcumin directly conjugates with thiol- or cysteine-rich molecules, reducing its availability to act on target pathways. Parallelly, curcumin binding to cysteamine in a cell-free thiol conjugation assay supports this hypothesis [43]. On the other hand, Kuttikrishnan et al. demonstrated that pan-caspase inhibition by z-VAD-FMK significantly reduced 24 h curcumin-mediated DNA damage in SUP-B15 and RS4;11 cells [39]. Thus, DNA damage can be secondary to cell death due to cell death-activated endo- and exonucleases rather than a primary initiating event. DNA integrity should therefore be assessed at earlier time points or in less toxic concentrations to clarify whether DNA damage precedes apoptosis.
Current evidence suggests that curcumin can inhibit oncogenic receptors and signalling pathways in ALL. In non-ALL cancer models, curcumin has been shown to inhibit hepatocyte growth factor and its receptor c-MET [74,75] and suppress human epidermal growth factor receptor 2 signalling [76,77]. In ALL, curcumin was reported to downregulate BCR-ABL mRNA expression in Bcr-Abl-positive SUP-B15 cells and corresponding xenograft in null mice [45]. Similar observations were reported in Bcr-Abl-positive chronic myeloid leukemia via competitive inhibition and epigenetic regulation [78,79]. Curcumin further synergised with imatinib (a first-line Bcr-Abl inhibitor) by suppressing AKT/mTOR signalling and downregulating the expression of BCR-ABL gene, an effect not seen with imatinib alone [45]. These findings suggest that curcumin may act through epigenetic or post-transcriptional regulation of Bcr-Abl expression. In addition, curcumin also inhibited JAK/STAT [25,37,45], PI3K/AKT [24,30,37,39,45], and NF-κB pathways [36] in ALL cells, which are known to be downstream canonical effectors of the Bcr-Abl signalling pathway [80,81,82]. Nevertheless, the evidence on Bcr-Abl signalling to date is mainly derived from studies of the P210 Bcr-Abl isoform in CML rather than the P190 isoform predominant in Philadelphia chromosome-positive ALL. Structurally, P190 Bcr-Abl protein lacks the Dbl-homology and Pleckstrin-homology domain with conserved ATP-binding pocket and major signalling pathways [83,84,85]. Consequently, the relevance of curcumin-mediated pathway inhibition in P190-driven ALL has not been directly established. In addition, direct molecular targets of curcumin in Bcr-Abl-negative ALL have yet to be identified, representing a critical gap in current knowledge.
Beyond oxidative stress and DNA-damaging activities, curcumin appears to target drug resistance-related molecular targets in reducing chemoresistance. Preclinical evidence demonstrates that curcumin downregulates MDR1 and WT1 genes and inhibits P-gp activity, highlighting its potential role as a chemosensitising agent in ALL [25,26,40]. It also prevented chemoresistance by inhibiting NF-κB activation, a pathway closely associated with chemoresistance in ALL cells [36]. Consistent with these findings, curcumin-mediated WT1 downregulation has been reported across other leukemia subtypes, including acute and chronic myeloid leukemia [27,86,87,88,89,90,91]. Beyond ALL, it downregulated MDR gene in other leukemias, including mouse lymphocytic leukemia [92], acute myeloid leukemia [23,93], and chronic myeloid leukemia [23,94,95]. Curcumin also retained cytotoxic or chemosensitising activity in P-gp-overexpressed cervical cancer and breast cancer cells [28,96]. Moreover, structurally modified curcuminoids have demonstrated P-gp inhibitory activity as well [94,97,98,99]. Current preclinical evidence suggests that curcumin may function primarily as an adjunct to conventional chemotherapy rather than a single-target chemotherapeutic agent. Figure 3 summarizes the molecular mechanisms of anti-leukemic properties of curcumin on ALL cells.
Despite its potent anticancer activity in vitro, the clinical translation of curcumin is severely limited by its poor bioavailability. Curcumin is limited by poor water solubility, poor intestinal absorption, chemical instability, and rapid metabolism and excretion [100,101]. Consequently, blood curcumin levels are frequently undetectable or extremely low, with the majority of curcumin excreted in the feces [102,103]. In addition, curcumin is highly unstable under neutral aqueous solution, where about 90% of curcumin degraded non-enzymatically within 30 minutes [104]. This degradation is partially mitigated in culture media with serum due to stabilising interactions with biomolecules [105,106]. However, such protection is unlikely in an animal model. Curcumin failed to suppress leukemia cell growth or improve survival in NOD/SCID mice [46], which may be partially due to these limitations. Consistently, curcumin was rapidly metabolised into glucuronidated and sulfated conjugates in the plasma of mice as early as 1 h after administration [46]. From a medicinal chemistry standpoint, the β-diketone bridge of curcumin contributes to both chemical instability and metabolic susceptibility [107]. To overcome these limitations, chemical modification [108,109,110] and pharmaceutical strategies including nanoparticles, liposomes, and exosomes [105,111,112] have been widely used to improve the bioavailability of curcumin in acute myeloid leukemia (AML) treatment [113,114]. Nanoparticles, such as mesoporous silica and gold-based curcumin formulations, have exhibited increased cytotoxicity in cancer cells than free curcumin, while also improving its stability and bioavailability [105,114]. Liposomes, including curcumin modified with hyaluronan to target CD44, show higher binding affinity to CD44-overexpressing AML cells and improved cellular uptake [112,113]. Nevertheless, evidence supporting the efficacy of nanoformulated curcumin in ALL remains limited.
The structural–activity relationship of curcumin explains its biological activity, metabolic stability, and therapeutic potential [115], although it has not been systematically evaluated in the ALL model. The phenolic hydroxyl groups contribute to both antioxidant and anti-inflammatory activities as well as the key conjugation site during Phase II metabolism [116]. Chemical substitution of these hydroxyl groups has been shown to enhance metabolic stability and solubility. On the other hand, phenolic methoxy substituents are more strongly associated with anti-inflammatory and anticancer effects [115]. Consistent with this, curcumin, which contains two phenolic methoxy groups, demonstrates greater anticancer activity than demethoxycurcumin (single phenolic methoxy group) and bisdemethoxycurcumin (no phenolic methoxy group; Figure 1) in suppressing TNF-induced NF-κB activation [117] and inducing cytotoxicity in human glioblastoma LN229 and GBM8401 cells [118]. Similarly, pure curcumin (95–99% purity) significantly suppressed WT1 mRNA and protein levels in MOLT-4 cells compared to a curcumin–demethoxycurcumin–bisdemethoxycurcumin mixture [27]. Synthetic curcumin analogues such as dimethoxycurcumin (Figure 1), which contains four phenolic methoxy groups, exhibited enhanced anticancer activity relative to curcumin, likely due to increased solubility, metabolic stability, cellular uptake, and bioavailability [108,109,119,120]. Supporting this, dimethoxycurcumin was proven less susceptible to liver microsomal enzymatic degradation and more potent in inducing DNA hypomethylation in ALL cells [34]. Similarly, polyphenols such as ferulic acid and ginger-derived compounds, including 6-gingerol, 6-shogaol, 10-gingerol, and 10-shogaol (Figure 1), share a common phenolic hydroxyl and methoxy structural feature and were reported to have similar anticancer, antioxidant, and anti-inflammatory mechanisms overlapping with those of curcumin [121,122,123,124,125,126,127,128,129,130]. These further highlight the importance of phenolic hydrocarbon structures in mediating curcumin’s biological activity.
In vivo and clinical evidence supporting the efficacy of curcumin in ALL remains scarce. In this systematic review, only two included articles reported animal data, and neither provided direct evidence that curcumin effectively eliminates ALL cells in animal models. Oral supplementation and intraperitoneal injection of curcumin failed to suppress the growth of SEM leukemia xenografts and did not improve survival outcomes in NOD/SCID mice [46]. A high dose of intraperitoneal injection of curcumin reduced SUP-B15 leukemic cells infiltration in the spleen of null mice [45]; however, the safety profile and reduction of leukemic cells in systemic circulation were not evaluated. To date, no published interventional studies have reported curcumin use in patients with ALL. Major clinical trial registries in the US (clinicaltrial.gov) and Europe (euclinicaltrials.eu) were searched to identify any ongoing or completed clinical trials evaluating curcumin in patients with ALL. We only identified a single registered trial conducted in Egypt between 2021 and 2024. This Phase II study (NCT05045443) [131] enrolled 30 paediatric patients aged 1 to 18 years with ALL and evaluated oral curcumin (500 mg turmeric-derived curcumin capsules, twice daily for one month), initiated during week 1 of the maintenance phase of chemotherapy. The primary outcome was the percentage of patients experiencing adverse events within four weeks. Although the trial status is listed as completed, no results have been published so far, leaving both the safety and therapeutic relevance of curcumin in patients with ALL unresolved. In addition, another related trial (NCT02100423) evaluated curcumin in combination with vitamin D, but it focused on patients with chronic lymphocytic leukemia or small lymphocytic lymphoma [132]. Similarly, it is completed, but the results remain unpublished. Despite these promising effects from in vitro studies, the optimal therapeutic dose of curcumin for ALL remains unclear. Evidence from studies in other cancers and non-cancer human conditions indicates that oral curcumin doses ranging from 20 mg/day up to 12 g/day are generally well tolerated, with minimal adverse effects [14,133,134]. Based on these findings, a similar dosing range could be considered in future ALL studies.
This systematic review, like others, has some limitations. Methodological heterogeneity of included articles was found, including inconsistent treatment procedure, variable curcumin concentrations, differing treatment durations, and variable endpoint measurements. These inconsistencies may lead to errors in data interpretation and analysis. Future studies should be standardised in experimental procedures, including cell seeding and treatment conditions. Another limitation of this systematic review is the purity of the curcumin used. Most commercial-grade or extracted curcumin from turmeric powder contains around 30% of demethoxycurcumin and bisdemethoxycurcumin [22,23,27]. The composition of extracted curcuminoids varies greatly depending on the Curcuma species [108]. Only 3 out of 26 included studies used curcumin that was ≥90% pure [27,29,46]. In addition, demethoxycurcumin and bisdemethoxycurcumin share similar anticancer activities but with weaker potency than curcumin, including cytotoxicity [118,135], P-gp inhibition [28], and PI3K/AKT suppression [77]. This significantly hampers the interpretation of anti-leukemic effects of curcumin, as it is unclear whether the reported biological effects are from curcumin, curcuminoids or a mixture. Future studies should assess the anti-leukemic effects of pure curcumin and explicitly report the purity of the compound used. In terms of literature searching, we only considered primary studies written in English, and this search strategy may have overlooked grey literature and other relevant articles. To minimise this risk, we manually searched for more relevant articles from the reference list of included articles. Lastly, a meta-analysis was not conducted due to the heterogeneity in outcome measures across studies.
5. Conclusions
This systematic review provides an understanding of the selective cytotoxicity and apoptosis-inducing effects of curcumin in ALL cells, with T-lineage cells being more susceptible than B-lineage cells. Curcumin activates both intrinsic and extrinsic pathways of apoptosis, downregulates PI3K/AKT and JAK/STAT signalling pathways, downregulates BCR-ABL, WT1 and MDR1, inhibits P-gp activity, and upregulates p15INK4b and RAF/MEK/ERK signalling. Upstream molecular mechanisms remain incompletely understood, but current evidence suggests the involvement of oxidative stress, DNA damage, and ceramide accumulation upon curcumin treatment. Despite promising in vitro results, in vivo evidence is limited, with only two ALL animal studies and no published clinical trials in patients with ALL. This highlights the need for Phase II/III clinical trials specifically targeting ALL, as most curcumin trials to date have examined other cancer types or non-ALL leukemias in general. Additional challenges for clinical translation include curcumin’s poor bioavailability and variability in curcuminoid composition. Further in-depth preclinical studies using standardised experimental protocols with well-designed clinical trials are essential to clarify curcumin’s therapeutic efficacy, optimal dosing, and safety profile as an anti-leukemic agent.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Puckett Y. Chan O. Acute Lymphocytic Leukemia Stat Pearls Stat Pearls Publishing Treasure Island, FL, USA 2023
- 2National Comprehensive Cancer Network NCCN Guidelines for Patients: Acute Lymphoblastic Leukemia National Comprehensive Cancer Network (NCCN) and NCCN Foundation Plymouth Meeting, PA, USA 2025
- 3National Cancer Institute Cancer Stat Facts: Leukemia—Acute Lymphocytic Leukemia (ALL)Available online: https://seer.cancer.gov/statfacts/html/alyl.html(accessed on 9 November 2025)
- 4Ekpa Q.L. Akahara P.C. Anderson A.M. Adekoya O.O. Ajayi O.O. Alabi P.O. Okobi O.E. Jaiyeola O. Ekanem M.S. A Review of Acute Lymphocytic Leukemia (ALL) in the Pediatric Population: Evaluating Current Trends and Changes in Guidelines in the Past Decade Cureus 202315 e 4993010.7759/cureus.4993038179374 PMC 10766210 · doi ↗ · pubmed ↗
- 5Institute for Health Metrics and Evaluation (IHME) Global Burden of Disease 2023 Results Available online: https://vizhub.healthdata.org/gbd-results/(accessed on 17 January 2026)
- 6Inaba H. Mullighan C.G. Pediatric acute lymphoblastic leukemia Haematologica 20201052524253910.3324/haematol.2020.24703133054110 PMC 7604619 · doi ↗ · pubmed ↗
- 7Sasaki K. Jabbour E. Short N.J. Jain N. Ravandi F. Pui C.H. Kantarjian H. Acute lymphoblastic leukemia: A population-based study of outcome in the United States based on the surveillance, epidemiology, and end results (SEER) database, 1980–2017 Am. J. Hematol.20219665065810.1002/ajh.2615633709456 PMC 9517941 · doi ↗ · pubmed ↗
- 8American Cancer Society Typical Treatment of Acute Lymphocytic Leukemia (ALL)Available online: https://www.cancer.org/cancer/types/acute-lymphocytic-leukemia/treating/typical-treatment.html(accessed on 9 November 2025)