Identification, Characterization and Expression Profiles of Xylogen-like Gene Family in Kiwifruit in Different Developmental Tissues and Under Various Abiotic Stresses
Caili Wang, Chen Li, Xiaoli Tang, Chunhua Li

TL;DR
This study identifies and analyzes 28 XYLP genes in kiwifruit, revealing their roles in growth, development, and stress responses.
Contribution
Systematic identification and characterization of the XYLP gene family in kiwifruit, including their expression and stress response roles.
Findings
28 AcXYLP genes were identified and grouped into five clades through phylogenetic analysis.
AcXYLP genes are involved in growth, development, and abiotic stress responses, with three genes (AcXYLP13, AcXYLP15, AcXYLP27) playing central roles.
Segmental duplication was the main driver of AcXYLP gene family expansion.
Abstract
This study investigates the XYLP gene family in kiwifruit, focusing on its roles in growth, development, phytohormone response, and abiotic stress. A total of 28 AcXYLP gene family members were identified and categorized into five clades. Most AcXYLPs are alkaline, predominantly hydrophobic, and tend to be structurally unstable. All the AcXYLPs are in silico predicted to be localized to the cell membrane. The 28 AcXYLP genes exhibit uneven distribution, with segmental duplication as the major driving force during gene family expansion. Transcriptome analysis, complemented by qRT-PCR validation, confirmed the involvement of XYLP genes in organ development, fruit ripening, and responses to phytohormones and abiotic stresses. This study lays a foundation for future investigations into the biological functions of XYLP genes in kiwifruit. The XYLP family is reported to be essential for the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
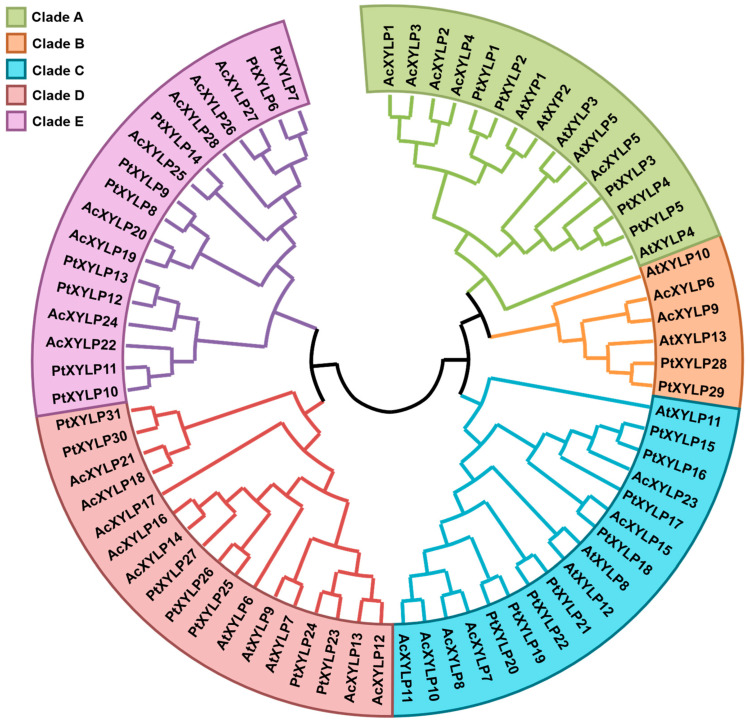 Figure 1
Figure 1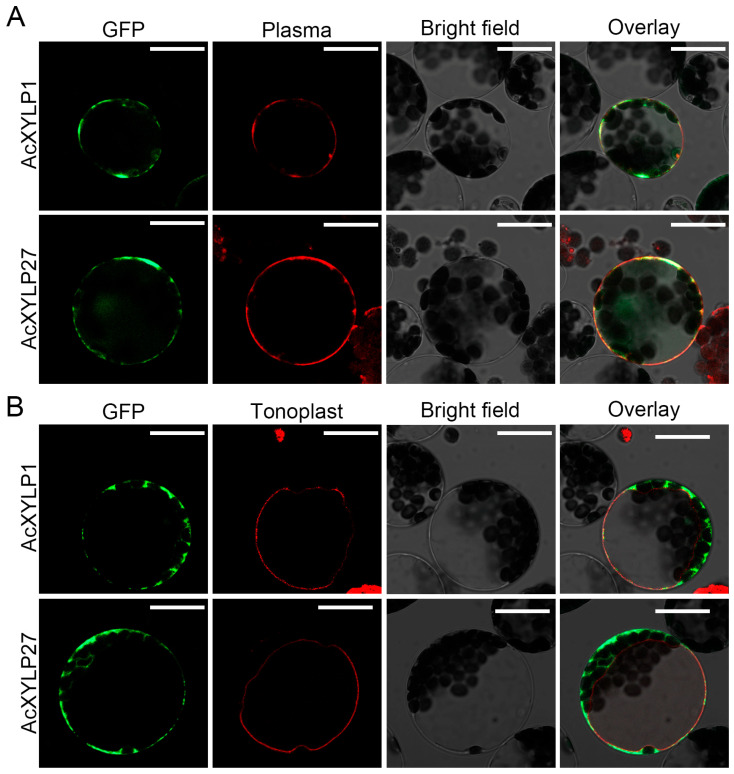 Figure 2
Figure 2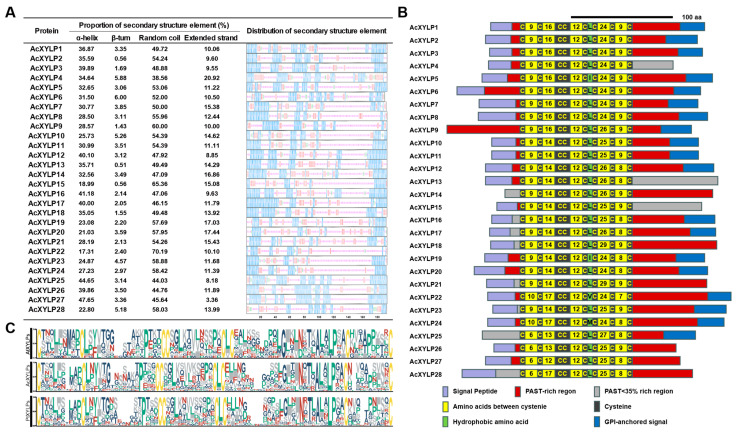 Figure 3
Figure 3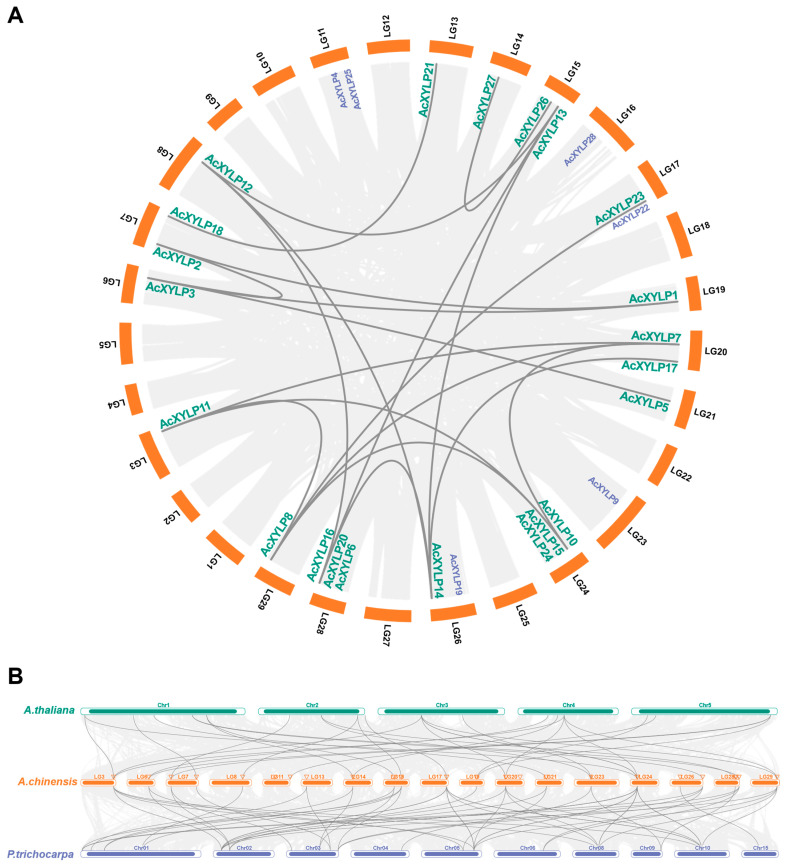 Figure 4
Figure 4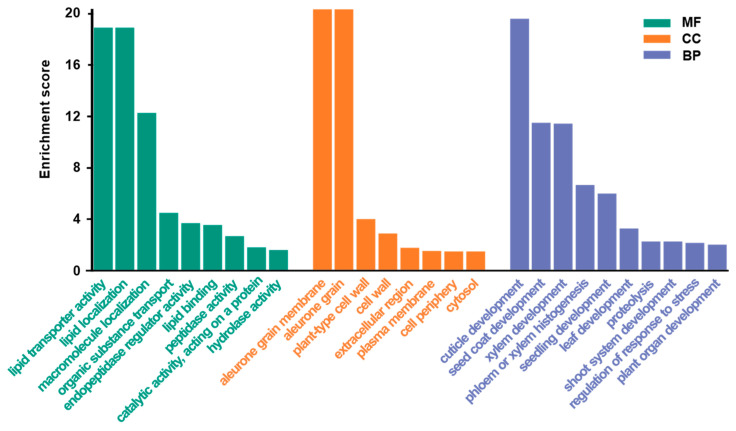 Figure 5
Figure 5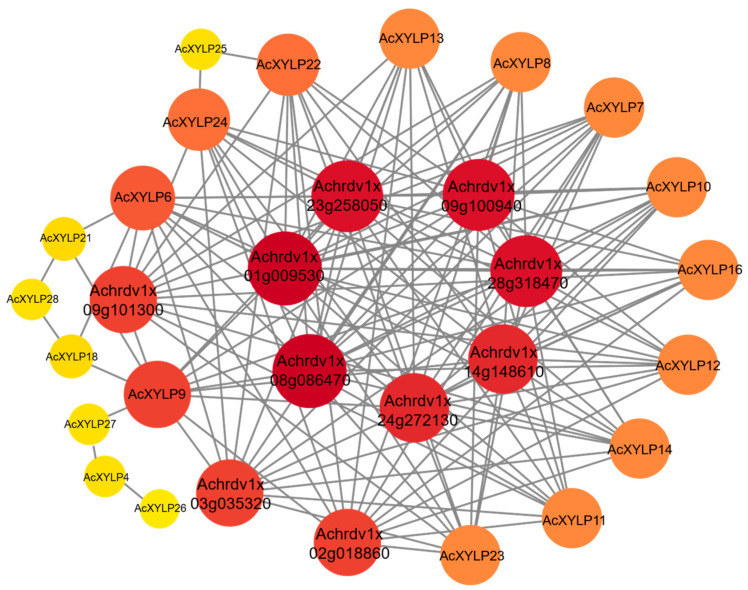 Figure 6
Figure 6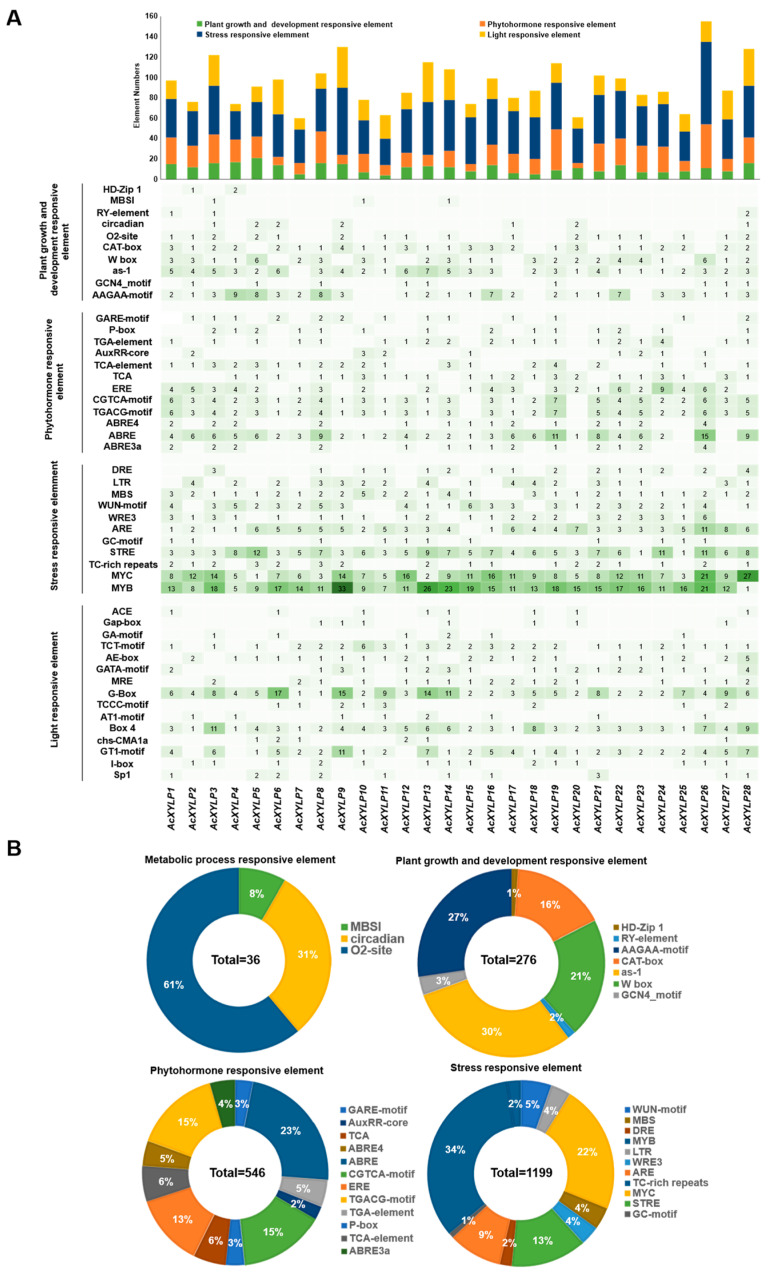 Figure 7
Figure 7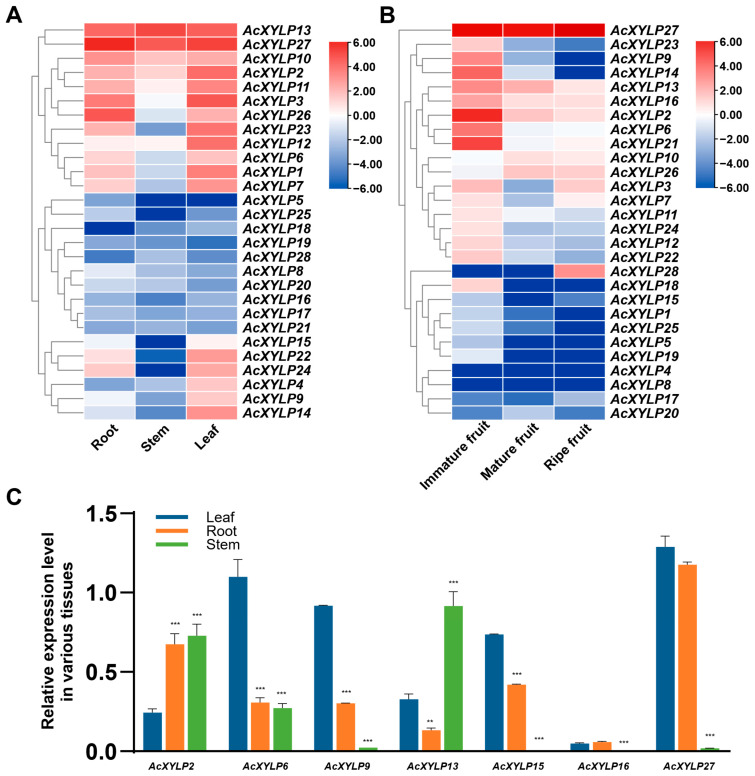 Figure 8
Figure 8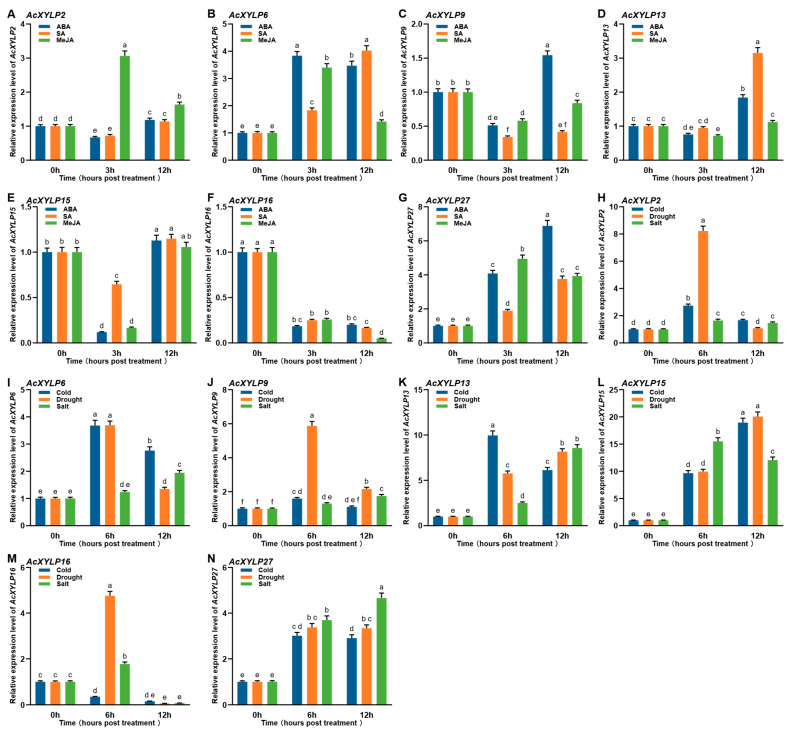 Figure 9
Figure 9- —Shandong Province Natural Science Foundation for Youths of China
- —Scientific Research Foundation of Dezhou University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Polysaccharides and Plant Cell Walls · Postharvest Quality and Shelf Life Management
1. Introduction
In higher plants, many extracellular proteins are involved in various biological processes, including intercellular communication, growth and developmental processes, as well as stress responses. Arabinogalactan proteins (AGPs), as extracellular proteins, are widely distributed in the plant kingdom and algae species, and are among the most complex macromolecule families in plants [1,2]. AGPs are a class of glycoproteins composed of large type II arabinogalactan (AG) glycan chains and a protein backbone rich in proline/hydroxyproline (P), alanine (A), serine (S), and threonine (T) [3,4,5,6]. The AG glycan chain and the protein backbone are linked together through O-glycosylation of hydroxyproline [6]. AGPs are present in almost all plant organs and cell types, and are involved in multiple processes such as cell expansion [7,8], cell proliferation [9], embryonic development [10,11], pollen tube growth [12], pollen development [13], cell wall remodeling [14], xylem differentiation [15], root development [16], responses to brassinosteroids [17] and salt stress [18].
The high functional diversity of AGPs can be attributed to the high complexity and specificity of their carbohydrate moieties and protein backbone composition [2,3]. In addition to the AGP domain, chimeric AGPs also possess other conserved domains. Based on the composition of conserved domains, chimeric AGPs can be further classified into plastocyanin-like AGPs (PLAs) [19,20], fasciclin-like AGPs (FLAs) [21], and xylogen-like proteins (XYLPs) [22]. As chimeric AGPs, the XYLPs have a unique structure containing an AGP domain and a conserved non-specific lipid-transfer protein (nsLTP) domain [23]. nsLTPs are a class of small, soluble and basic proteins belonging to a complex, multigene family, primarily defined for their ability to transport lipid molecules in vitro [24]. The nsLTP exhibits high stability, which is attributed to its unique structure: it contains eight conserved cysteine residues that form four disulfide bonds, thereby stabilizing the protein’s tertiary structure [25]. Xylogen is first isolated and identified as an intercellular signaling molecule from suspension-cultured Zinnia leaf cells [26,27]. Previous studies have shown that xylogen, secreted by differentiating vascular cells, can be transported to adjacent undifferentiated cells and promote their differentiation into tracheary elements [22].
The fine regulation of vascular differentiation is triggered by various signaling regulators, including phytohormones (such as auxin, cytokinin and brassinolide), xylogen and plant small peptides, among which xylogen plays a crucial role [28]. In Arabidopsis, the double knockouts of xyp1 xyp2 exhibit obvious defects in vascular structure, with discontinuous veins and vessels in leaves and roots [22]. The xylp7 T-DNA insertion mutant in rice demonstrates a shorter length of internodes except for the basal internode [29]. The overexpression of PtXYLP1 results in the defective venation pattern in Arabidopsis cotyledons [30]. Meanwhile, there may be a close interaction between phytohormones and xylogen, as the expression of xylogen is upregulated by auxin, cytokinin and gibberellin. In addition to plant growth, xylogen may also be involved in plants’ response to abiotic stress, with XYLP genes being almost regulated by salt, drought and cold treatment [29,31].
The identification at a genome-wide scale of the xylogen-encoding gene family has been achieved as an effective strategy for gene function analysis in species such as Arabidopsis [23], rice [29], poplar [30], and moso bamboo [31]. However, no relevant studies on members of XYLPs have been reported in kiwifruit or other wine plants to date. As an economically important fruit crop, kiwifruit is rich in vitamins and minerals, yet the cultivation is often challenged by abiotic stresses, resulting in reduced yield [32,33]. Prompted by these unanswered questions, we performed a comprehensive bioinformatics analysis of the XYLP gene family in the kiwifruit genome. In Actinidia chinensis, 28 AcXYLP genes in total were identified. The physicochemical properties, phylogenetic relationships, gene and protein structures, chromosomal locations, duplication events, gene ontology (GO) annotations, protein–protein interaction networks, and promoter cis-acting elements were systematically characterized. The expression patterns of seven representative XYLP genes were also investigated in response to various abiotic stresses and phytohormone treatments. The results presented here serve as a resource for the XYLP family in kiwifruit and will facilitate targeted functional investigations of its members for further studies.
2. Materials and Methods
2.1. Sequence Identification and Characterization
Protein sequences of 13 AtXYLP were retrieved from The Arabidopsis Information Resource (https://www.arabidopsis.org/). Those sequences served as queries for a BLASTP (v2.13.0+) search against the Kiwifruit Genome Database 3.0 (https://kiwifruitgenome.atcgn.com) [34] with an E-value threshold < 1 × 10^−7^. Iterative BLASTP searches were performed using newly identified candidates as queries until no further homologs were found. Sequences were identified as AcXYLPs [29] based on the presence of both predicted arabinogalactan (AG) glycomodules, identified manually according to established criteria, by analyzing the presence of the Pro/Ala/Ser/Thr (PAST)-rich motifs as AGP markers in the sequence which contained at least one region with ≥35% PAST residues [30,35], and an nsLTP-like domain (PF00234). Domain prediction was performed via the Pfam hidden Markov model (HMM) (http://pfam.xfam.org/) and subsequently validated using InterProScan 108.0 (https://www.ebi.ac.uk/interpro/, accessed on 29 January 2026) [36] with default settings. For comparative analysis, 31 poplar XYLP (PtXYLP) sequences were obtained from Phytozome 14 (https://phytozome-next.jgi.doe.gov/), as described previously [30].
2.2. Analysis of Physicochemical Properties and Protein Structure
The physicochemical properties, including theoretical isoelectric point (pI) and molecular weight (MW) of AcXYLPs were predicted using the ExPASy ProtParam tool (https://web.expasy.org/protparam/, accessed on 29 January 2026) [37]. The generation of the hydropathicity profiles was applied with ProtScale (https://web.expasy.org/protscale/, accessed on 29 January 2026) [37], using the Kyte–Doolittle scale with a window size of 9. SignalP 6.0 (https://services.healthtech.dtu.dk/services/SignalP-6.0/, accessed on 29 January 2026) was introduced in the prediction of the signal peptides of the AcXYLPs [38] with the default organism setting “Eukarya”. Potential GPI-anchor modification signals were predicted using NetGPI 1.1 (https://services.healthtech.dtu.dk/services/NetGPI-1.1/, accessed on 29 January 2026) [39]. The online tool NetNGlyc 1.0 (https://services.healthtech.dtu.dk/services/NetNGlyc-1.0/, accessed on 29 January 2026) [40] was used in the N-linked glycosylation site prediction, applying a potential threshold of >0.5. Secondary structure composition (alpha-helix, extended strand, random coil) was predicted using the SOPMA method via the NPS@ server (https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma_f.html, accessed on 29 January 2026) [41] by using the following parameters: similarity threshold = 8, window width = 17, and the default parameters of the tool. Tertiary structures were modeled using the automated mode of SWISS-MODEL (http://swissmodel.expasy.org/) [42], and model quality was assessed based on GMQE scores.
2.3. Phylogenetic, Genomic, and Gene Structure Analysis
Multiple sequence alignment of AtXYLP, AcXYLP, and PtXYLP was conducted with the built-in ClustalW tool using default parameters using MEGA 11. Subsequently, a phylogenetic tree was constructed from this alignment using the neighbor-joining (NJ) method in MEGA 11 [43,44], applying the Poisson model, pairwise deletion, and 1000 bootstrap replicates. Chromosomal locations, segmental duplication events, and genomic collinearity among A. thaliana, A. chinensis, and P. trichocarpa were analyzed using MCScanX v1.0.0 [45] with the following key parameters: match score = 50, match size = 5, e-value = 1 × 10^−10^, max gaps = 25. The resulting collinear blocks were visualized using the “Dual Synteny Plot” function in TBtools v2.311 [46]. The ratios of non-synonymous (Ka) to synonymous (Ks) substitutions for duplicated gene pairs were calculated with the “Simple Ka/Ks Calculator” in TBtools [46]. Gene structures (exon-intron organization) were visualized with the Gene Structure Display Server 2.0 (http://gsds.cbi.pku.edu.cn/) [47] using genomic and CDS sequences as input. Conserved motifs within the nsLTP domain were aligned using MUSCLE (v3.8.31, https://www.ebi.ac.uk/jdispatcher/, accessed on 29 January 2026) [48] with default settings and rendered as a sequence logo with WebLogo 3 (https://weblogo.threeplusone.com/) [49].
2.4. Promoter and Functional Annotation Analysis
Sequences spanning 2000 bp upstream of the translational start site of each AcXYLP gene were retrieved and screened for cis-acting elements via the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 29 January 2026) [50] with default settings. Gene ontology (GO) annotation was performed by submitting AcXYLP sequences, using the Arabidopsis thaliana GO information as a reference, with the “Go Annotation” module built in TBtools. Enrichment analysis results were visualized using the “ggplot2” module within TBtools [46]. Protein–protein interaction (PPI) networks were predicted using the STRING database (v12.0, https://cn.string-db.org/) with the following parameters: organism “Actinidia chinensis”, meaning of network edges set to “confidence”, and a minimum required interaction score of 0.400 (medium confidence). Functional enrichment in the network was ranked based on the “node degree” parameter, as provided in the “string_node_degrees.tsv” file downloaded from the STRING Exports page. The resulting network was downloaded and visualized using Cytoscape (v3.10.4, https://cytoscape.org/).
2.5. Plant Materials and Treatments
Seeds of A. chinensis “Hong Yang” were surface-sterilized and germinated on half-strength Murashige and Skoog (MS) medium supplemented with 0.8% agar at 24 °C under a 12 h light (8000 lux)/12 h dark photoperiod. For tissue-specific expression analysis, samples from shoot apices (AP), young leaves (YL, the 1st and 2nd leaves from the apex), mature leaves (ML, fully expanded leaves), young stems (YS, internodes of freshly growth), mature stems (MS, lignified stems), and roots (RT) were collected from 3-month-old soil-grown plants. For phytohormone treatments, the aerial parts of 4-week-old seedlings were evenly sprayed with 100 μM solutions of abscisic acid (ABA), salicylic acid (SA), or methyl jasmonate (MeJA), and collected at 3 and 12 hours post-treatment. For abiotic stress treatments, 4-week-old plants were subjected to (i) low temperature (4 °C in a growth chamber), (ii) drought stress (supplied with 10% (w/v) PEG6000 solution), or (iii) salt stress (supplied with 200 mM NaCl solution), and collected at 6 and 12 hours post-treatment. Upon collection, all tissue samples were immediately frozen in liquid nitrogen and subsequently maintained at −80 °C. For each treatment, three biological replicates were established, each constituted by pooling tissues from a minimum of five plants.
2.6. Expression Analysis
The in silico expression analysis was performed on a laboratory server running Ubuntu 22.04.3. Expression profiles were derived from publicly available RNA-seq datasets in the NCBI SRA database, specifically under accessions PRJNA328414 (various tissues) and PRJNA187369 (fruit development stages: DAP20, DAP120, DAP127). The NCBI SRA Toolkit (https://github.com/ncbi/sra-tools/wiki/01.-Downloading-SRA-Toolkit, accessed on 29 January 2026) was used to retrieve sequencing data. SRA records listed in index.txt were downloaded using the command prefetch --option-file ./index.txt. FASTQ files were then extracted from the downloaded SRA files with the command fastq-dump --split-3 ./SRR.sra. Raw sequencing reads were pseudoaligned and quantified using Kallisto (v0.50.0) [51] to obtain Transcripts Per Million (TPM) values. A kiwifruit transcriptome index was first constructed with the command kallisto index ./Ac.cds.fa -i ./index. Quantification was then performed using kallisto quant -i ./index -o output_dir -b 100 ./SRR_1.fastq ./SRR_2.fastq. The “Heatmap” module in TBtools [46] was employed to generate clustered heatmaps with the following parameters: clustering method = “Average” and distance metric = “Euclidean”.
For the validation of the expression profile, total RNA was extracted using the Polysaccharide Polyphenol Plant Total RNA Kit (Tiangen, Beijing, China, DP441) following the manufacturer’s protocol, including an on-column DNase I digestion step. RNA concentration and purity were assessed by NanoDrop (Wilmington, DE, USA) and agarose gel electrophoresis. For cDNA synthesis, 1 µg of total RNA was used in the reverse-transcription, by using the PrimeScript™ RT Master Mix (Takara, Beijing, China, RR036A) according to the manufacturer’s instructions, in a final volume of 20 µL. Gene-specific primers used in quantitative real-time (qRT)-PCR are listed in Table S1. qRT-PCR analyses were conducted on an Agilent Mx3000P system (Agilent Technologies, Santa Clara, CA, USA), employing TB Green^®^ Premix Ex Taq™ II (Takara, RR820A), in a final volume of 20 µL. The thermal cycling conditions were: 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s, 58 °C for 30 s, and 72 °C for 15 s, with a subsequent melting curve analysis (95 °C for 15 s, 60 °C for 60 s, 95 °C for 15 s) to confirm primer specificity. For normalization purposes, AcActin was used as the reference gene, and relative expression was calculated via the 2^−ΔΔCt^ method. All the samples and qRT-PCR reactions were run in triplicate, and each sample consists of 3–5 kiwifruit plants.
2.7. Subcellular Localization
Subcellular localization was predicted in silico using Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/, accessed on 29 January 2026) [52]. For experimental validation, the full-length coding sequences of AcXYLP1 and AcXYLP27 (without the stop codon) were amplified and cloned into the pSAT6-EYFP-N1 vector (via restriction enzymes BamHI and SpeI) to generate C-terminal fusions with GFP under the control of a double CaMV 35S promoter. The recombinant plasmids (35Spro::AcXYLP-GFP) and control vectors (35Spro::GFP) were purified using a Plasmid Miniprep Kit (Axygen, Union City, CA, USA). Arabidopsis leaf mesophyll protoplasts were isolated from 4-week-old Col-0 plants using an enzyme solution (1.5% cellulase R10, 0.4% macerozyme R10, 0.4 M mannitol, 20 mM KCl, 20 mM MES pH 5.7, 10 mM CaCl_2_, 0.1% BSA). Approximately 2 × 10^4^ protoplasts were transfected with 20 µg plasmid DNA via the PEG-calcium method [53]. For co-localization studies, protoplasts were co-transfected with the GFP-fusion constructs and the plasma membrane marker SCAMP1-RFP [54] or the tonoplast marker TPK1-RFP [55] at a 1:1 molar ratio. After incubation in the dark at 22 °C for 16–20 h, fluorescence signals were observed using a confocal laser scanning microscope (Zeiss LSM 880, Jena, Germany) with excitation/emission settings of 488 nm/505–530 nm for GFP and 561 nm/575–620 nm for RFP. Co-localization was assessed using the Pearson correlation coefficient. For each condition, images were acquired from 5 to 10 cells per field of view, and quantification was performed on 20–30 cells per replicate. All experiments were independently repeated at least three times.
2.8. Statistical Analysis
Data were presented as the mean ± SD from a minimum of three independent biological replicates, with all statistical analyses and graphs generated using GraphPad Prism software (v 10.6). For qRT-PCR-derived tissue-specific expression profiles, statistical significance was determined by one-way ANOVA with Tukey’s HSD post hoc test for pairwise comparisons against controls at each time point; significance was denoted as p < 0.05 (), p < 0.01 (), and p < 0.001 (). For the expression profile analyses under phytohormone and abiotic stress treatments, a two-way ANOVA was conducted, and where overall effects were significant, pairwise comparisons were further examined using Tukey’s HSD test, with statistically significant differences (p < 0.05) indicated by different superscript letters in the corresponding figure.
3. Results
3.1. Identification, Physicochemical Properties, and Evolutionary Relationship of the AcXYLP Family Members
To identify putative XYLP gene family members in kiwifruit, a BLASTP search was conducted against the kiwifruit genome database, with the protein sequences of ZeXYP1 and AtXYLPs used as queries, followed by a self-BLASTP search with the candidate kiwifruit XYLPs [22]. The potential XYLP members were verified for the presence of AGP and conserved nsLTP domain, ultimately identifying a total of 28 XYLP genes in the Actinidia chinensis genome, which share significant sequence similarity (E-value < 1 × 10^−7^). For convenience, the abbreviation “Ac” for Actinidia chinensis was placed before the gene family name (XYLP). Based on the BLASTP results, they were named as AcXYLP1-AcXYLP28, and the basic AcXYLP physicochemical properties are shown in Table 1. A phylogenetic tree was generated from 72 XYLPs from Arabidopsis (dicotyledonous herb model plant), poplar (dicotyledonous woody model plant) and kiwifruit. In the unrooted phylogenetic tree, AcXYLPs were classified into five clades (Clade A–E). Further analysis of the phylogenetic tree revealed Clade E comprised poplar and kiwifruit XYLPs. The absence of Arabidopsis corresponding genes suggests that this branch may evolved independently in woody plants (Figure 1).
The physicochemical property analysis of the AcXYLPs revealed that the amino acid lengths of the identified proteins range from 143 (AcXYLP26) to 210 (AcXYLP9), with the least (AcXYLP26, 14.90 kDa) and maximum (AcXYLP9, 21.87 kDa) molecular weights observed among the AcXYLPs. The theoretical pI of AcXYLPs ranges from 3.79 to 9.11, with half of the proteins being alkaline (pI > 7.0). The hydropathicity AcXYLPs were mainly positive (except AcXYLP14), indicating that they were dominantly hydrophobic. The instability index analysis found that most AcXYLPs were structurally unstable (instability index > 40), except AcXYLP16 and AcXYLP18 (instability index < 40). The prediction of subcellular localization indicated that all AcXYLPs were located on the cell membrane (Table 1). To verify the in silico prediction of AcXYLP subcellular localizations, AcXYLP1 and AcXYLP27 were randomly selected and transiently expressed in Arabidopsis mesophyll protoplasts, using SCAMP1-RFP [54] and TPK1-RFP [55] as markers for plasma membrane and tonoplast, respectively. The recombinant vectors of 35Spro::AcXYLP1-GFP and 35Spro::AcXYLP27-GFP were constructed and transiently transformed into Arabidopsis mesophyll protoplasts via PEG-calcium transfection, respectively. As predicted, the signals of AcXYLP1-GFP and AcXYLP27-GFP fusion proteins were clearly observed in the plasma membrane of Arabidopsis mesophyll protoplasts, and no GFP fluorescence signal was present in the tonoplasts (Figure 2).
3.2. Gene Structure, Conserved Motif and Protein Structure of the AcXYLP Family Members
To further understand the evolutionary relationship of XYLP gene family in kiwifruit, the gene structures of Arabidopsis, poplar and kiwifruit were analyzed with the Gene Structure Display Server, displayed in Figure S1. Most AcXYLP genes contain two introns and three exons, and homologous genes within the same clade exhibit similar exon–intron structures (Figure S1). In Clade E, the AcXYLP24 gene, along with its homologous genes PtXYLP10 and PtXYLP13, contains three introns and four exons. Notably, the AcXYLP4 and AcXYLP25 genes were exceptions, as they lack introns and were composed of a single exon (Figure S1). Secondary-structure analysis was performed on the AcXYLPs, and the results showed that all proteins comprised an α-helix, β-turn, random coil and extended strand, with various proportions and orders of these elements. Among all the AcXYLPs, the random coil occupies the highest proportion, ranging from 38.56% to 70.19%, followed by the α-helix and extended strand. The β-turn occupies the lowest proportion, ranging from 0.56% to 6.0% (Figure 3A). Among the 28 AcXYLPs, 25 were predicted to contain an N-terminal signal peptide, indicating that they could be classified into the secretory pathway as extracellular proteins. Most AcXYLPs were found to have a GPI (glycosylphosphatidylinositol) anchor at the C-terminus, which is a crucial domain for anchoring proteins to the plasma membrane (Figure 3B, Table S2). This result was highly consistent with the subcellular prediction of AcXYLPs (Table 1).
Similarly to other XYLPs, the AcXYLP also contains an nsLTP domain with eight well-conserved cysteine residues arranged in the order of C_1_-X-C_2_-X-P-X-C_3_C_4_-X-C_5_XC_6_-X-C_7_-X-C_8_ (Figure 3C). The hydrophobic residue between C_5_ and C_6_ is typically leucine (L), with only a small proportion of exceptions being isoleucine (I), valine (V), alanine (A), phenylalanine (F) and methionine (M) (Figure 3C). The hydrophobic residues and the conserved eight cysteine residues form a hydrophobic core in the tertiary structure, which is crucial for the lipid binding and transport processes. The glycosylation modification of XYLP is crucial for its catalytic function. All AcXYLPs contain at least one region rich in Pro/Ala/Ser/Thr (PAST) (PAST residues ≥ 35%), which provides moieties for potential glycosylation modifications. Additionally, there was at least one putative AG glycomodule within the PAST-rich region, where the amino acids were regularly arranged as Ala-Pro, Pro-Ala, Ser-Pro, Pro-Ser, Thr-Pro, and Pro-Thr (Table S2). The tertiary structures of the AcXYLPs were further predicted by SWISS-MODEL, with significant differences observed in the aggregation angle and structural adjacency of amino acids among each AcXYLP, which may be attributed to variations in the arrangement and proportion of secondary structural elements within the proteins (Figure 3A and Figure S2, Table S9). As expected, the high sequence similarity and tertiary structure of the nsLTP domain indicate its functional conservation (Figure S3, Table S10). Therefore, the analysis of gene structures, conserved motifs, and protein structures indicated high evolutionary conservation within the XYLP gene family.
3.3. Chromosomal Localization, Duplication and Collinear Relationship of the AcXYLP Family Members
The 28 AcXYLP genes were unevenly distributed on the 18 kiwifruit chromosomes. Specifically, there were three genes on chromosomes 24 and 28; two genes on chromosomes 7, 11, 15, 17, 20 and 26; and the remaining 10 genes were uniformly distributed, with one gene on chromosomes 3, 6, 8, 13, 14, 16, 19, 21, 23 and 29 (Figure 4A). As a fundamental evolutionary mechanism, gene duplication is common to all species and plays a pivotal role in the generation of new genes and species evolution [56]. The result showed that the AcXYLP genes had undergone 19 segmental duplication events, with no tandem duplication events observed in the kiwifruit genome, indicating that segmental duplication events might be the major force in the expansion of the AcXYLP gene family (Figure 4A). Furthermore, Ka/Ks ratios were calculated to evaluate the evolutionary selection pressure acting on the AcXYLP gene pairs generated by segmental duplication events. Among the 19 AcXYLP pairs analyzed, 18 exhibited Ka/Ks ratios below one. The pair AcXYLP14/AcXYLP17 was excluded from this calculation (denoted as N/A) due to high sequence divergence (pS ≥ 0.75), which precludes reliable Ka/Ks estimation. The highest ratio was observed for AcXYLP7/AcXYLP8 (0.51), while the lowest was found for AcXYLP10/AcXYLP11 (0.18) (Table S3), implying the XYLP genes in kiwifruit evolved under a strong purifying selective pressure. The evolutionary relationships of orthologous XYLP genes were analyzed through comparative synteny among kiwifruit, Arabidopsis, and poplar genomes. The collinear analysis revealed that most AcXYLP genes had a high synteny with XYLP genes in other species (Figure 4B, Table S4). Compared to Arabidopsis, one additional AcXYLP orthologous gene was identified in poplar (Figure 4B), which may indicate a closer phylogenetic relationship with poplar.
3.4. GO Annotation and Protein–Protein Interaction Network Prediction of the AcXYLP Family Members
To gain a preliminary understanding of the biological functions of the 28 AcXYLPs, gene ontology (GO) was employed for functional annotation. GO enrichment analysis revealed that these differentially expressed AcXYLPs were associated with multiple biological processes, cellular components, and molecular functions (Figure 5, Table S5). At the molecular-function level, the AcXYLPs were mainly involved in lipid transporter activity (GO:0005319), lipid localization (GO:0010876), lipid binding (GO:0008289) and catalytic activity on a protein (GO:0140096). At the biological-process level, the AcXYLPs were mainly involved in cuticle development (GO:0042335), xylem development (GO:0010089), seedling development (GO:0090351), leaf development (GO:0048366), shoot system development (GO:0048367) and regulation of response to stresses (GO:0080134). The most significant enrichment was observed at the cellular-component level, mainly enriched in the aleurone grain membrane (GO:0032578), plant-type cell wall (GO:0009505) and extracellular regions (GO:0005576) (Figure 5). These results indicated that the AcXYLPs were primarily involved in lipid transport, localization and binding, and may play significant roles in plant organ development and plant response to stresses.
To better understand the potential biological functions of XYLPs in kiwifruit, the potential proteins interacting with AcXYLPs were analyzed using the STRING database V 12.0. The protein–protein interaction network was predicted and analyzed based on the related proteins in A. chinensis. According to the prediction, 20 of the 28 AcXYLPs established connections with its potential interacting proteins (Figure 6, Table S6). The results indicated that the XYLP family members, namely AcXYLP6, AcXYLP7, AcXYLP8, AcXYLP9, AcXYLP10, AcXYLP11, AcXYLP12, AcXYLP13, AcXYLP14, AcXYLP16, AcXYLP22, AcXYLP23 and AcXYLP24, play crucial roles in plant growth and development (Figure 6). Specifically, these genes were predicted to be involved in the regulation of the transition of meristems from the vegetative to the reproductive phase, and also participated in the plant’s responses to auxin and ethylene (Figure 6, Table S6).
3.5. Cis-Acting Elements Present in the AcXYLP Gene Family Promoters
In order to investigate the putative roles of the AcXYLP genes in kiwifruit, the cis-acting elements in their promoter sequences were extracted and analyzed with plantCARE. Promoter analysis of the AcXYLP genes identified 2620 putative cis-acting elements, which were classified into 48 functional types (Table S7). According to the predicted biological functions, these cis-acting elements fall into four major groups: plant growth and development, phytohormone signaling, stress responses, and light-responsive elements. Each AcXYLP gene promoter contained various types of cis-acting elements, and the numbers differed (Figure 7). Among all the cis-acting elements, the stress-responsive elements accounted for the highest percentage at 45.8%, followed by light-responsive elements (21.5%), phytohormone-responsive elements (20.8%), and plant growth and development responsive element (11.9%). The methyl jasmonate (MeJA)-responsive elements and abscisic acid (ABA)-responsive elements constituted the largest proportions within the phytohormone-related category, accounting for 32% and 30%, respectively, with 175 and 164 identified in the 26 AcXYLP genes (Figure 7A). Furthermore, all of these genes contained salicylic acid (SA)-responsive elements (Figure 7A). The promoters of all AcXYLP genes contained MYB and MYC binding sites, which constituted 34% and 22% of the stress response elements, respectively (Figure 7A).
The plant growth and development category contained ten cis-acting elements. Six were implicated in key processes: differentiation of palisade mesophyll cells (HD-Zip 1), meristem expression (CAT-box), senescence expression (W box), shoot expression (as-1) and endosperm expression (GCN4_motif and AAGAA-motif) (Figure 7B), and the shoot expression (as-1) were extensively distributed in the promoters of 26 AcXYLP genes, with 82 occurrences (Figure 7A). Multiple cis-acting elements related to the phytohormone-responsive element category were found in AcXYLP gene promoters, including gibberellin (GARE-motif and P-box), auxin (TGA-element and AuxRR-core), salicylic acid (TCA-element and TCA), ethylene (ERE), MeJA (CGTCA-motif and TGACG-motif) and ABA (ABRE4, ABRE and ABRE3a) (Figure 7B). These phytohormone-responsive elements were widely present and can be found in almost every AcXYLP gene, especially the ABA-, MeJA-, and SA-responsive elements (Figure 7A). Among the stress-responsive elements, there are not only MYB- and MYC-responsive elements but also dehydration-responsive elements (DRE), low-temperature-responsive elements (LTR), drought-responsive elements (MBS), and wound-responsive elements (WUN-motif) (Figure 7B). The MYB- and MYC-responsive elements were closely associated with drought stress, salt stress, and cold stress [57,58,59]. The results above suggested that members of the AcXYLP gene family play important roles in plant responses to phytohormones and various stress conditions.
3.6. Expression Patterns of the AcXYLP Family Members in Different Tissues and Developmental Stages
Analysis of the gene expression patterns can provide important clues for elucidation of gene potential functions. Based on the RNA-seq datasets publicly available at NCBI Sequence Read Archive (SRA), the expression patterns of all 28 AcXYLP genes in different kiwifruit tissues were analyzed (Table S8). RNA-seq transcriptomic data for tissue-specific expression patterns was obtained under the accession of PRJNA328414 and PRJNA187369, encompassing vegetative organs such as roots, stems, and leaves, as well as three stages of fruit ripening: immature fruit, mature fruit, and ripe fruit (Figure 8A,B). A hierarchical cluster analysis was performed using the expression levels of AcXYLP genes. This analysis revealed that all 28 AcXYLP genes were expressed in at least one vegetative organ or reproductive developmental stage. Based on the transcriptome profiling data revealed by the PRJNA328414, AcXYLP13 and AcXYLP27 exhibited high expression levels across all vegetative organs (Figure 8A). AcXYLP2, AcXYLP10 and AcXYLP11 were abundantly expressed in roots and leaves but showed low expression levels in stems (Figure 8A). Nine genes (AcXYLP1, AcXYLP3, AcXYLP6, AcXYLP7, AcXYLP12, AcXYLP22, AcXYLP23, AcXYLP24 and AcXYLP26) were primarily expressed in roots and leaves (Figure 8A). The expression levels of ten genes (AcXYLP5, AcXYLP8, AcXYLP16, AcXYLP17, AcXYLP18, AcXYLP19, AcXYLP20, AcXYLP21, AcXYLP25, and AcXYLP28) were relatively low in all vegetative organs, with some being undetected in specific tissues. Additionally, AcXYLP4, AcXYLP9, AcXYLP14 and AcXYLP15 were predominantly expressed in leaves (Figure 8A).
During the kiwifruit ripening process described in PRJNA187369, the genes AcXYLP10, AcXYLP26 and AcXYLP28 were found to be upregulated. The expression levels of 15 genes, including AcXYLP2, AcXYLP3, AcXYLP6, AcXYLP7, AcXYLP9, AcXYLP11, AcXYLP12, AcXYLP13, AcXYLP14, AcXYLP16, AcXYLP18, AcXYLP21, AcXYLP22 AcXYLP23, and AcXYLP24, were significantly reduced during fruit ripening (Figure 8B). It was worth noting that three genes (AcXYLP2, AcXYLP13 and AcXYLP27) were expressed at high levels in both vegetative and reproductive tissues (Figure 8A,B). This showed that these AcXYLP genes might play crucial roles in all tissues throughout the developmental stages of kiwifruit. To confirm the expression profiles of AcXYLP genes in different tissues, primers were designed for qRT-PCR amplification of the 28 AcXYLP genes. Due to non-specific amplification, with the trial of three primer pairs for each gene, the expression patterns of only seven genes were successfully obtained: AcXYLP2, AcXYLP6, AcXYLP9, AcXYLP13, AcXYLP15, AcXYLP16, and AcXYLP27 (Figure 8C). Fortunately, these seven genes were distributed across five clades, and these genes were further used in subsequent experiments. Gene-specific primers used in qRT-PCR are listed in Table S1 and melting curves are shown in Figure S4. Samples were collected from 3-month-old ‘Hong Yang’ soil-grown plants, and the expression levels of these seven genes during the vegetative growth stage were detected, which were highly consistent with results previously described in RNA-seq (Figure 8A,C).
3.7. Expression Profiles of the AcXYLP Family Members in Response to Abiotic Stresses and Phytohormone Treatments
Previous studies have shown that XYLP genes play roles in plant responses to multiple phytohormone treatments and abiotic stresses [30,31]. To determine whether AcXYLP genes also function in abiotic stresses and phytohormones, the expression levels of seven genes (AcXYLP2, AcXYLP6, AcXYLP9, AcXYLP13, AcXYLP15, AcXYLP16, and AcXYLP27) were examined in kiwifruit plants subjected to cold, drought, salt, ABA, SA, and MeJA treatments (Figure 9). Expression pattern analysis revealed that most AcXYLP genes were significantly upregulated in response to cold, drought, and salt stress, compared to untreated controls, except AcXYLP2 and AcXYLP16 (Figure 9A–G). Under salt stress, the expression of AcXYLP2 showed an initial downregulation followed by upregulation (Figure 9A). The expression of AcXYLP16 was downregulated under cold stress, and exhibited an initial upregulation followed by downregulation under drought and salt stress (Figure 9B). Notably, the expression levels of AcXYLP13, AcXYLP15 and AcXYLP27 genes were significantly upregulated under three stress conditions: cold, drought, and salt stress (Figure 9D,E,G).
Most AcXYLP genes showed similar expression profiles after exposure to phytohormone treatments (Figure 9H–N). The expression of AcXYLP6 and AcXYLP27 was upregulated under ABA, SA and MeJA treatments, and the expression of AcXYLP16 was downregulated (Figure 9I,M,N). The expression of AcXYLP13 and AcXYLP15 was first downregulated and then upregulated by ABA, SA and MeJA treatments (Figure 9K,L). AcXYLP2 was initially downregulated by ABA and SA treatments, then upregulated, while it was upregulated by MeJA treatment (Figure 9H). AcXYLP9 was initially downregulated by ABA and MeJA treatments, then upregulated, while it was downregulated by SA treatment (Figure 9J). All these results indicated that all seven AcXYLP genes participated in plant response to abiotic stresses and phytohormone treatments, with AcXYLP13, AcXYLP15 and AcXYLP27 potentially playing significant roles in enhancing plant tolerance to abiotic stresses.
4. Discussion
4.1. Molecular Characteristics and Evolutionary Analysis of the AcXYLP Genes
In this study, 28 AcXYLP genes were identified in the genome of Actinidia chinensis. These AcXYLPs contained at least one PAST-rich (PAST residues ≥ 35%) region and a highly conserved nsLTP domain (C_1_-X-C_2_-X-P-X-C_3_C_4_-X-C_5_XC_6_-X-C_7_-X-C_8_) (Figure 3B), which was in agreement with the characteristics of XYLPs in Arabidopsis [23] and poplar [30] plants. Notably, there were significant differences in the physicochemical properties of AcXYLPs, including variations in the number of amino acids, isoelectric points, and instability indices (Figure 3A). Despite these differences, all XYLPs were in silico-predicted to be hydrophobic, consistent with their roles in transmembrane signaling transduction (Table 1). In addition, the differences in the physicochemical parameters of AcXYLP might be attributed to the structural diversity of the XYLP genes. In agreement with previous reports, the members of AcXYLP gene family contained one to four exons and zero to three introns, and genes within the same phylogenetic clade exhibited similar gene structures [23,29,30,31] (Figure S1). AcXYLP4, AcXYLP25, and PeXYLP1 exhibited a reduced number of exons; each possessed only a single exon [31] (Figure S1). This reduction might be due to the disruption of splicing sites caused by the insertion of transposable elements. Additionally, the subcellular localization of the AcXYLPs was in silico-predicted to be on the plasma membrane, and this was proved on AcXYLP1 and AcXYLP27 in Arabidopsis (Figure 2), suggesting a potential role in transmitting intercellular signals and inducing differentiation into vascular tissues.
To elucidate the evolutionary relationships of the XYLP gene family, the phylogeny of kiwifruit was compared with that of Arabidopsis and poplar. It was found that, as a woody plant, the genes of the XYLP family in kiwifruit had likely evolved independently, paralleling the findings in poplar [30]. There are 28 AcXYLP genes in the kiwifruit genome, a number higher than the XYLP genes in Arabidopsis (13) [23], rice (21) [29], and moso bamboo (23) [31], but lower than the 31 XYLP genes present in poplar [30]. Gene duplication plays a fundamental role in the expansion and development of plant gene families during evolution, contributing to the generation of novel genes and the diversification of gene functions [56]. The kiwifruit genome contained 29 chromosomes, with 28 AcXYLP genes randomly distributed across 18 of these chromosomes (Figure 4A). Notably, the distribution of AcXYLP genes did not correlate with chromosome size, and 11 chromosomes did not harbor any AcXYLP genes (Figure 4A). These findings suggested that the AcXYLP gene family in kiwifruit may have undergone segmental loss events during evolution. In A. chinensis, all identified duplicated XYLP gene pairs originated from segmental duplication events (Table S4). The absence of tandemly duplicated gene pairs suggested that segmental duplication was the dominant force shaping the kiwifruit XYLP family. While XYLP expansion in other species commonly involves both segmental and tandem events [29,30,31], kiwifruit appeared to have followed a divergent evolutionary path, likely shaped by its specific long-term adaptive pressures.
4.2. Prediction of the Potential Biological Functions of the AcXYLP Genes
Cis-acting elements are reported to be pivotal regulators of transcriptional patterns and levels [60]. Analysis of the AcXYLP gene promoters revealed an enrichment of cis-acting elements responsive to plant growth and development, stress, phytohormones, and light, underscoring their potential roles in these key biological processes. For instance, there are abundant elements related to meristem (CAT-box), shoot (as-1), and endosperm (GCN4_motif and AAGAA-motif) development (Figure 7B). Additionally, the AcXYLP genes contained many phytohormone-responsive elements, including those for auxin, ethylene, and gibberellin (Figure 7A), which played crucial roles in various physiological processes, such as seed germination [61,62], root development [63,64], stem elongation [65,66], vascular differentiation [67], and fruit ripening [68].
In addition to elements related to stress responses, such as dehydration, salt stress response (DRE), low-temperature response (LTR), wound response (WUN-motif), and drought response (MBS), the AcXYLP genes also contained a significant number of MYB and MYC elements closely associated with low-temperature, drought, and salt stress [57,58,59] (Figure 7B). It was suggested that XYLPs in kiwifruit played key roles in regulating plant growth, development, and responses to abiotic stresses. GO enrichment analysis identified a substantial number of AcXYLP genes associated with lipid transport and catalytic activity. Furthermore, these AcXYLP genes were also involved in the development of leaves, xylem, and shoots, as well as in the plant’s response to environmental stresses (Figure 5). The analysis of the protein–protein interaction network revealed that many AcXYLPs participated in plant growth and hormone signaling (Figure 6), which aligned with the findings from the GO enrichment analysis (Figure 5).
4.3. Expression Profile Reveals the Potential Roles of the AcXYLP Genes in Abiotic Stresses and Phytohormone Responses
According to relevant studies, multiple XYLPs have been found to be involved in the development of plant vascular systems [22,26,27,29,30]. In Arabidopsis, the XYLP double-knockout mutant (xyp1 xyp2) displayed discontinuous veins and vessels in both leaves and roots, which suggested that the absence of XYLPs significantly impaired the formation of vascular tissues [22]. The defects observed in the internodes of the rice xylp7 mutant and the leaf veins of 35Spro::PtXYLP1 overexpressing plants indicated that XYLPs play a crucial role in the development of the vascular system [29,30]. In this study, two genes, AcXYLP13 and AcXYLP27, exhibited high expression levels in all vegetative tissues (Figure 8A,C). The remaining 16 genes were abundantly expressed in at least one vegetative tissue, with 12 genes being mainly expressed in roots and leaves, while 4 genes were mainly expressed in leaves (Figure 8A). These results indicated that AcXYLPs played a significant role in plant vegetative growth, which was similar to findings described in poplar and moso bamboo [30,31]. As kiwifruit matures, the expression of most AcXYLP genes was downregulated. However, the expression levels of AcXYLP10, AcXYLP26, and AcXYLP28 were significantly upregulated, while AcXYLP27 consistently maintained a high expression level (Figure 8B). These findings suggested that members of the AcXYLP gene family play a crucial role in regulating the fruit ripening process.
Abiotic stress triggers the expression of XYLP genes. For example, OsXYLP7 is induced under drought, salt, and cold stress [29], and PeXYLP2 and PeXYLP9 are activated by PEG and NaCl [31]. The AcXYLP genes exhibited diverse expression patterns in response to abiotic stress and various phytohormone treatments. After exposure to cold, drought, and salt stress, the expression of the seven AcXYLP genes, except for AcXYLP16, was found to be upregulated. Notably, AcXYLP13, AcXYLP15, and AcXYLP27 displayed significant increases in their expression levels (Figure 9A–G). Phytohormone treatments also induce the expression of XYLP genes, with PtXYLP1 and PtXYLP2 being induced by NAA, 6-BA, and GA treatments [30]. The majority of AcXYLP genes showed modulated expression at different time points during phytohormone treatments. It was noteworthy that under ABA stress, the expression levels of genes AcXYLP2, AcXYLP6, AcXYLP9, AcXYLP13, AcXYLP15, and AcXYLP27 were significantly upregulated (Figure 9H–N). This upregulation was also observed in response to cold, drought, and NaCl abiotic stresses (Figure 9A–G). These findings might be attributed to the role of ABA, as a stress hormone, which played a crucial role in mediating various abiotic stress responses in plants [69].
Consistent with cis-acting element predictions, the promoters of AcXYLP genes contained multiple MYB and MYC elements. The results implicated the kiwifruit XYLP gene family in crucial abiotic stress response pathways.
5. Conclusions
This study identified and characterized members of the XYLP gene family in the kiwifruit genome using bioinformatics methods to elucidate the structural and functional roles. A total of 28 AcXYLP gene family members were identified, which were unevenly distributed across 18 chromosomes. The expansion of the AcXYLP gene family was suggested to have primarily originated from segmental duplication events and had undergone strong purifying selection pressure throughout evolution. Based on the phylogenetic analysis, the AcXYLP genes were grouped into five clades, exhibiting similar gene structures and conserved motif compositions within each clade. Subsequently, the potential biological functions of AcXYLP genes were investigated in silico through analysis of promoter cis-acting elements, gene ontology annotations, and protein–protein interaction networks. This analysis indicated that AcXYLP genes might play pivotal roles in the regulation of growth, development, hormone regulation, and abiotic stress responses of A. chinensis. RNA-seq transcriptomic analysis, complemented by qRT-PCR validation, suggested the involvement of XYLP genes in the growth of vegetative tissues and the fruit ripening process in kiwifruit. Moreover, the AcXYLP genes were implicated in the plant’s response to phytohormones (e.g., ABA, SA, and MeJA) and abiotic stresses (e.g., cold, drought, and salt). Overall, this study provided valuable insights into the role of the XYLP gene in the growth and development of kiwifruit, as well as its response to abiotic stress. This research established a robust foundation for future functional validation and applied studies in molecular breeding.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Nguema-Ona E. Coimbra S. Vicré-Gibouin M. Mollet J.-C. Driouich A. Arabinogalactan proteins in root and pollen-tube cells: Distribution and functional aspects Ann. Bot.201211038340410.1093/aob/mcs 14322786747 PMC 3394660 · doi ↗ · pubmed ↗
- 2Du H. Clarke A.E. Bacic A. Arabinogalactan-proteins: A class of extracellular matrix proteoglycans involved in plant growth and development Trends Cell Biol.1996641141410.1016/S 0962-8924(96)20036-415157509 · doi ↗ · pubmed ↗
- 3Ellis M. Egelund J. Schultz C.J. Bacic A. Arabinogalactan-Proteins: Key Regulators at the Cell Surface?Plant Physiol.201015340341910.1104/pp.110.15600020388666 PMC 2879789 · doi ↗ · pubmed ↗
- 4Carpita N.C. Gibeaut D.M. Structural models of primary cell walls in flowering plants: Consistency of molecular structure with the physical properties of the walls during growth Plant J.1993313010.1111/j.1365-313X.1993.tb 00007.x 8401598 · doi ↗ · pubmed ↗
- 5Ma Y. Johnson K. Arabinogalactan proteins—Multifunctional glycoproteins of the plant cell wall Cell Surf.2023910010210.1016/j.tcsw.2023.10010236873729 PMC 9974416 · doi ↗ · pubmed ↗
- 6Seifert G.J. Roberts K. The biology of arabinogalactan proteins Annu. Rev. Plant Biol.20075813716110.1146/annurev.arplant.58.032806.10380117201686 · doi ↗ · pubmed ↗
- 7Kaur D. Moreira D. Coimbra S. Showalter A.M. Hydroxyproline-O-Galactosyltransferases Synthesizing Type II Arabinogalactans Are Essential for Male Gametophytic Development in Arabidopsis Front. Plant Sci.20221393541310.3389/fpls.2022.93541335774810 PMC 9237623 · doi ↗ · pubmed ↗
- 8Lee K.J. Sakata Y. Mau S.L. Pettolino F. Bacic A. Quatrano R.S. Knight C.D. Knox J.P. Arabinogalactan proteins are required for apical cell extension in the moss Physcomitrella patens Plant Cell 2005173051306510.1105/tpc.105.03441316199618 PMC 1276029 · doi ↗ · pubmed ↗