Natural Genetic Variation Impacts Stress-Induced Quiescence and Regeneration in Response to Rapamycin
Sahiti Peddibhotla, Miriam Gonzaga, Tricia Zhang, Yasha Goel, Jun Sun, Benjamin R. Harrison, Daniel E. L. Promislow, Hannele Ruohola-Baker

TL;DR
This study shows that natural genetic differences affect how rapamycin influences cell recovery after radiation, with mitochondrial function playing a key role.
Contribution
The study identifies mitochondrial dynamics as a key factor in genetic variation affecting rapamycin response.
Findings
Genetic background significantly influences rapamycin's effect on quiescence and regeneration after radiation.
Mitochondrial dynamics are more strongly linked to rapamycin sensitivity than DNA damage repair processes.
Natural genetic variation in DGRP reveals critical differences in therapeutic response to rapamycin.
Abstract
In response to ionizing radiation (IR), both adult and cancer stem cells enter reversible cell cycle arrest at the G1/S transition to evade apoptosis and subsequently re-enter the cell cycle to regenerate damaged tissue. Entry into and exit from this arrest, known as “quiescence,” is governed by the inhibition of mTORC1. The pharmacological suppression of mTORC1 with rapamycin prevents quiescent stem cells from re-entering the cell cycle and impairs tissue regeneration. Rapamycin holds great therapeutic promise in preventing tumor regrowth from dormant cancer stem cells. Yet the extent to which genetic background impacts the known variation in the pharmacological response of rapamycin remains unknown. Here, we show that natural genetic variation across the Drosophila Genetics Reference Panel (DGRP) drives substantial differences in the rapamycin-mediated suppression of post-IR…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
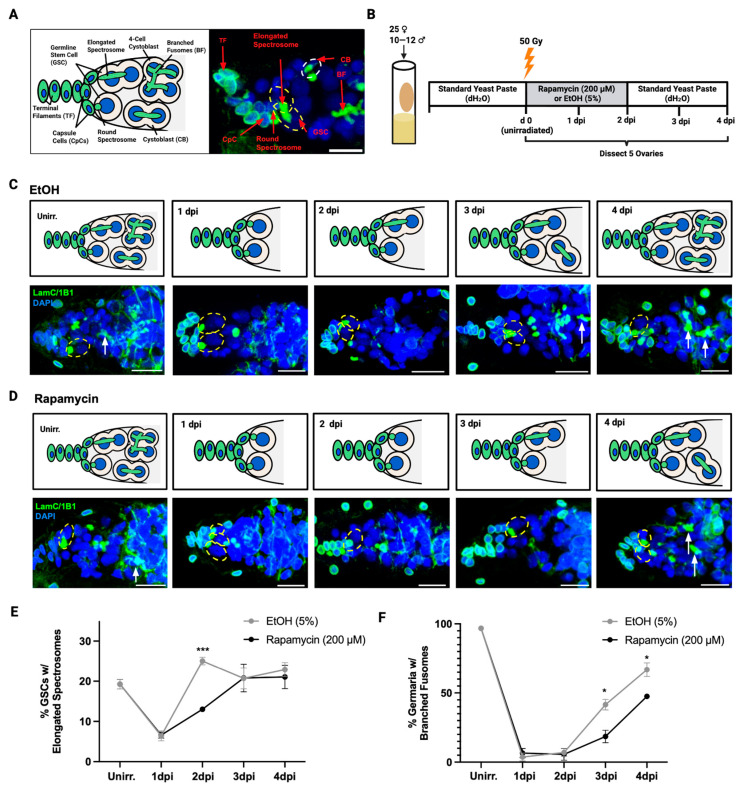 Figure 1
Figure 1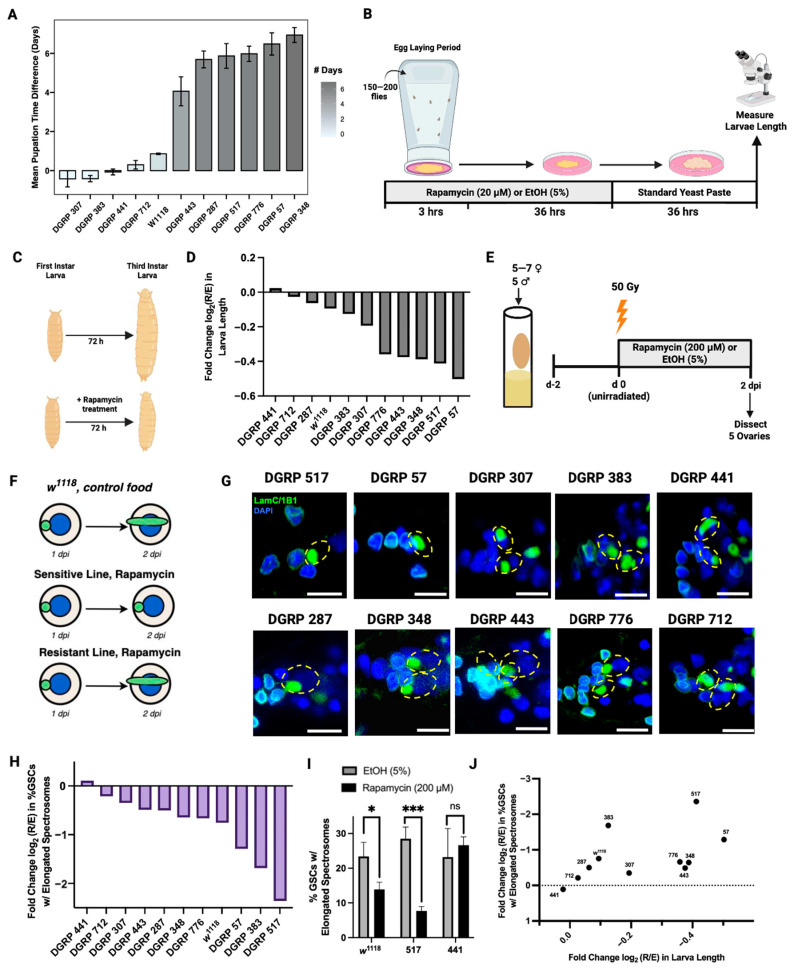 Figure 2
Figure 2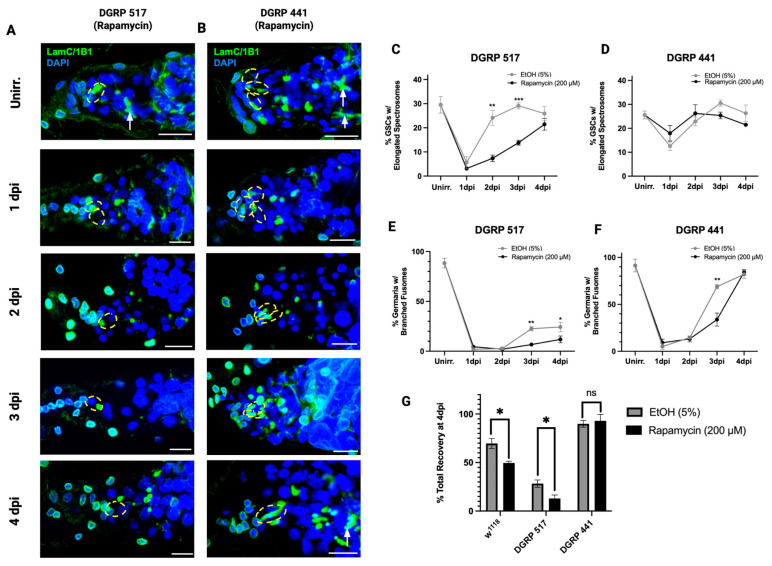 Figure 3
Figure 3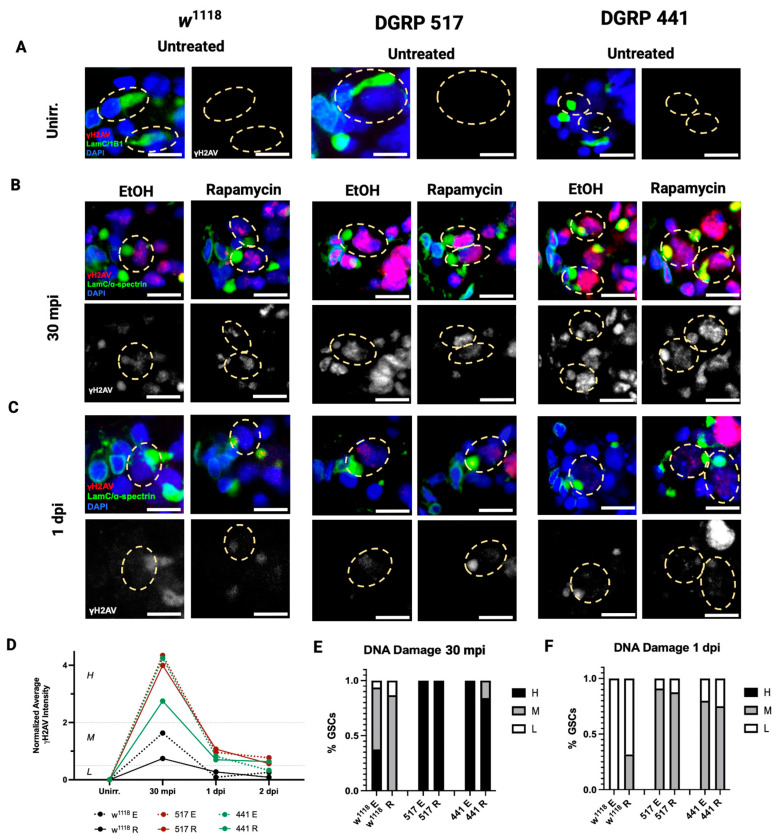 Figure 4
Figure 4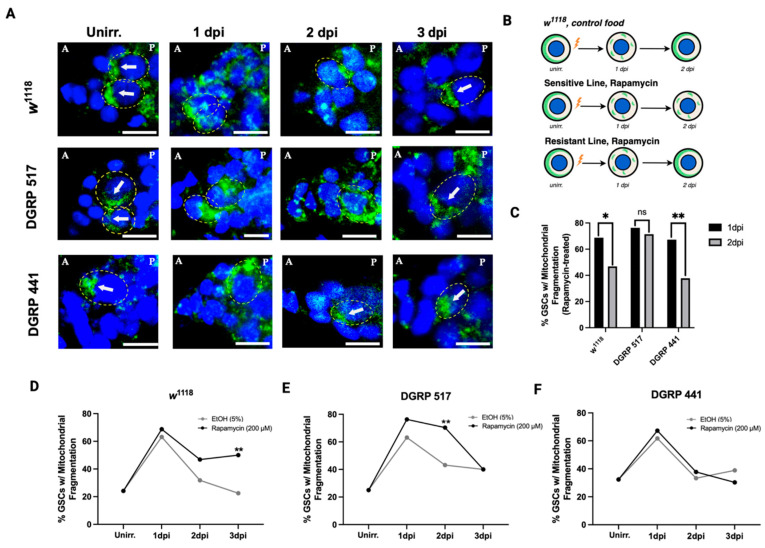 Figure 5
Figure 5- —USDA cooperative agreement
- —NIH National Institute on Aging
- —HRB Fellows Program (Sahiti Peddibhotla)
- —National Institutes of Health
- —NHLBI Progenitor Cell Biology Consortium
- —SCGE
- —DOD
- —Stem Cell Gift Funds
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHippo pathway signaling and YAP/TAZ · PI3K/AKT/mTOR signaling in cancer · Cancer-related Molecular Pathways
1. Introduction
Diverse types of adult stem cells undergo asymmetric divisions to maintain tissue homeostasis and renew the stem cell pool. Under stress conditions, protecting the stem cell pool is critical for effective post-injury regeneration [1,2]. One of the most consequential stressors to cells is genotoxic stress, which can inflict severe cellular damage. Failure to repair these damages leads to the accumulation of dysfunctional organelles, misfolded proteins, and genomic instability, which can either trigger apoptosis, impair regeneration, or lead to tumorigenesis [3,4,5,6,7,8].
When exposed to ionizing radiation (IR), stem cells evade death and permanent damage by entering a transient state of proliferative dormancy at the G1/S transition, or quiescence (G0), while their differentiated progeny undergo apoptosis [2,9,10]. Quiescence preserves the cellular integrity of stem cells by allowing the repair and replacement of damaged DNA and organelles, before resuming the cell cycle. The DNA damage response post irradiation, across multiple organisms, is directed by the FOXO transcription factor, which upregulates DNA repair proteins [11,12]. In Drosophila GSCs, among the downstream factors of FOXO activation that induce cell cycle arrest, is the inhibition of the mechanistic target of rapamycin complex I (mTORC1) [13,14,15]. The inhibition of mTORC1 alleviates the repression of autophagy [16,17,18]. The removal of damaged organelles through autophagy, which includes mitochondrial autophagy or mitophagy, is required for stem cell quiescence. Following autophagy, organelle biogenesis is required to exit quiescence and resume stem cell division [14,19,20]. Once damage is amended, mTORC1 reactivation promotes the switch from quiescence to post-injury proliferation and tissue regeneration.
The inhibition of mTORC1 is a general requirement of stem cell dormancy. Reduced mTORC1 signaling characterizes quiescence across multiple stem cell types in mammals and flies, including hematopoietic, neural, and satellite cells, and is observed in mammalian embryonic diapause and cancer stem cell dormancy [21,22,23,24,25,26]. By preventing mTORC1 inhibition, with a knockdown of the negative regulator Tsc1, irradiated Drosophila GSCs of the w^1118^ line do not enter quiescence [14]. In quiescent GSCs of the w^1118^ line, treatment with the mTORC1 inhibitor rapamycin prolongs quiescence and prevents cell cycle re-entry [14]. In cancer stem cells (CSCs), a subset of tumor cells possessing similar capabilities of self-renewal and maintenance as adult stem cells, rapamycin prevents exit from proliferative dormancy, preventing unwanted tumor growth post chemoradiation therapy [27,28,29]. However, tumor cell populations display high intra and inter tumoral heterogeneity and variability across individuals, leading to inconsistent drug responses [30,31]. Resistance to rapamycin has previously been observed in cancer cells [32,33]. Little is currently understood about how natural genetic variation can modulate the pharmacology of rapamycin and its effects in post-radiation regeneration.
A few studies have demonstrated that the effects of rapamycin can vary among individuals across populations [34,35,36]. For instance, mice display sex differences in lifespan extension with rapamycin [34]. Two fly studies have demonstrated variation in multiple responses to rapamycin, including fecundity and development, using the Drosophila Genetics Reference Panel (DGRP) [35,36]. The DGRP is a collection of ~200 genetically diverse wild-derived lab-inbred strains which represent natural genetic variation within a population [37]. The DGRP therefore provides a powerful tool to assess the impact of natural genetic variation on drug response [38,39].
Rapamycin delays development in Drosophila, such that larvae treated with rapamycin take longer to develop to the pupal stage [40,41]. However, among the DGRP, there is extensive variation in the sensitivity of development time to rapamycin, including some lines showing greater sensitivity than the common lab strain w^1118^, and others remaining resistant to doses over two orders of magnitude higher than the screening dosage [36]. Variation in sensitivity to rapamycin during pupation delay is a strong indication that Drosophila populations maintain standing variation that influences the response of rapamycin on mTORC1 effector pathways.
We have previously shown that rapamycin extends irradiation-induced quiescence in the w^1118^ background [13]. Given recent studies that demonstrate genetic variation in response to rapamycin, we postulate that the sensitivity of IR-induced quiescence to rapamycin might depend on genetic background. Here we survey 10 DGRP lines, and w^1118^, for sensitivity to rapamycin in prolonging quiescence after irradiation. We first find that the effect of rapamycin on GSC quiescence depends strongly on genetic background. We then hypothesize that the variation in the quiescence response might be the same pharmacogenetic variation in the effect of rapamycin on developmental delay. If this is true, we predict that the degree of sensitivity in quiescence across these lines would correlate to the variation in their sensitivity during development. We instead find that sensitivity in pupation delay does not correspond to sensitivity in quiescence, suggesting that genetic variation in mTORC1 effector pathways is specific to cellular processes that are involved in development or in GSC quiescence. We then investigated the mechanism underlying variation in rapamycin-mediated quiescence extension by examining two pathways required for quiescence exit: DNA repair or the removal and replacement of damaged mitochondria. We find that lines at extreme ends of sensitivity to rapamycin in quiescence show no differences in DNA damage repair following IR induction. However, we find that they differ substantially in the time to resolve damaged mitochondria. By using lines that vary in sensitivity to rapamycin, our study suggests that genetic variation in the regulation of mitochondrial removal and replacement may contribute to the variation in sensitivity to rapamycin in quiescence.
2. Materials and Methods
2.1. Fly Stocks and Culture Conditions
w^1118^ and DGRP stocks were obtained from the Bloomington Drosophila Stock Center at Indiana University. The following stocks were used: w^1118^ (RRID:BDSC_3605), DGRP-57 (RRID:BDSC_29652), DGRP-307 (RRID:BDSC_25179), DGRP-383 (RRID:BDSC_28190), DGRP-441 (RRID:BDSC_28198), DGRP-517 (RRID:BDSC_25197), DGRP-287 (RRID:BDSC_28165), DGRP-348 (RRID:BDSC_55019), DGRP-443 (RRID:BDSC_28199), DGRP-776 (RRID:BDSC_28229), and DGRP-712 (RRID:BDSC_25201).
2.2. Gamma Irradiation Treatment and Dissection
Two days prior to ionizing radiation treatment, flies (0–3 days old) were cultured in empty vials with standard yeast paste (active dry yeast in H_2_O) for 48 h at 25 °C. On the day of irradiation, five unirradiated females from each genotype were dissected (within one hour of irradiation), and the remainder were transferred into empty vials without yeast paste and treated with 50 Gy of gamma irradiation. A Cs-137 Mark I Irradiator (Foss Therapy Services Inc., Pacoima, CA, USA) was used to administer the proper irradiation dosage as instructed by the dosage chart. At every timepoint, five of the irradiated females per treatment and genotype were dissected, while the remaining irradiated females were flipped into new vials with the respective treatment in yeast paste with young unirradiated males added at a ~1:1 ratio to females.
2.3. Development Assay
For each DGRP line assayed, 150–200 parents aged 3–5 days laid eggs for 3 h at 25 °C in egg chambers containing grape agar plates with yeast paste dissolved with either 20 uM rapamycin in 5% EtOH or solvent control (5% EtOH). Plates with embryos were collected after 3 h. Yeast paste was changed to standard yeast paste at 36 h post oviposition, and larvae were measured at 72 h post oviposition.
2.4. Rapamycin Treatment
A 4 mM stock was prepared in 200-Proof ethanol and stored at −20 °C. From 4 mM rapamycin, 1 mL of 200 µM in 5% EtOH was prepared and mixed with 0.5 g of active dry yeast. To prepare control food, 1 mL of 5% EtOH was prepared in H_2_O and mixed in 0.5 g of active dry yeast. For irradiated flies, treatment or control was administered to the side of vials for 48 h. For larvae, 20 µM rapamycin treatment or solvent control in yeast paste was administered at the center of grape agar plates in the egg collection chamber for the duration of oviposition (3 h) and 36 h post oviposition. Standard and supplementary yeast paste was prepared with 0.5 g of active dry yeast for every 1 mL of H_2_O.
2.5. Immunocytochemistry
Fly ovaries were dissected in cold PBS, immediately fixed in 4% paraformaldehyde for 15 min, washed in PBT (PBS containing 0.2% Triton X-100) three times for 10 min each, and stored in PBS at 4 °C for 24–96 h. Dissected samples from all timepoints were simultaneously blocked in PBTB (PBT containing 0.2% BSA, 5% normal goat serum) for one hour at room temperature. Ovaries were incubated overnight at 4 °C with the following primary antibodies: mouse anti-1B1 (RRID: AB_528070 1:30), mouse anti-ɑ-spectrin (RRID mouse anti-Lamin C (RRID: AB_528339 1:30), and rabbit anti-γH2AV (RRID: AB_828383 1:200). After three 10 min washes with PBT, secondary fluorescence antibodies were utilized including anti-rabbit Alexa 488 (RRID: AB_221544 1:250) and anti-mouse 568 (RRID: AB_2535773 1:250) for 2 h at room temperature. Samples were washed once with PBT for 10 min, incubated with DAPI (diluted with PBS to 2 μg/mL) for 15 min, and washed two times with PBS for 10 min each. The samples were mounted in glycerol and analyzed and imaged on a Leica SPE5 confocal laser-scanning microscope. Image processing and editing were performed using ImageJ v2.16.0/1.54p. For all images, intensity was uniformly adjusted, and the background was removed using ImageJ. Unedited images were used for γH2AV intensity quantifications.
2.6. Intensity Quantifications
For every GSC for each treatment, line, and timepoint, the mean intensity was calculated from ROIs drawn over γH2AV puncta within nuclei using ImageJ. Intensities of GSCs (n = 10–19) were averaged for each timepoint and treatment, then normalized to the intensity of the untreated, unirradiated timepoint for each respective line. Normalized intensities were categorized into low damage (>0.5), moderate damage (0.5–2), and high damage (2<).
2.7. Statistical Analysis
Student’s t-test (two-way, unpaired), two-way ANOVA, and Spearman’s correlations were computed using R v4.2.1. Graphs and diagrams were created using R, GraphPad Prism 10, and BioRender.
3. Results
3.1. Rapamycin Delays Post-IR Regeneration of the Drosophila Germline
We first aimed to determine the extent to which the post-IR regeneration of the Drosophila germline is affected by rapamycin treatment. The Drosophila ovary is composed of approximately 16 developing follicles, or ovarioles, with six to seven developing egg chambers [42]. The germarium, typically housing two to three GSCs, sits at the anterior tip of the ovariole. The GSCs are adjacent to the stem cell niche, consisting of capsule cells (CpCs). Both the niche CpCs and apical terminal filaments (TFs) can be identified with lamin C (LamC) to stain their nuclear lamina (Figure 1A). GSCs undergo asymmetric divisions to produce both a self-renewing stem cell and a daughter cystoblast (CB) [43].
The cell cycle of GSCs can be tracked by examining spectrosome morphology, identified with spectrosome-specific antigens, such as 1B1 and spectrins. The S-phase of the cell cycle is characterized by a fused or elongated spectrosome, which bridges a GSC and a CB. In Drosophila GSCs, injury-induced quiescence occurs at the G1/S transition. Elongated spectrosomes indicate that GSCs have re-entered the S-phase of the cell cycle and are no longer quiescent [14,43]. The recovery of the germline follows GSC division and is accomplished when the daughter CB undergoes several rounds of incomplete divisions to produce a 16-cell cyst. One of the sixteen cells then differentiates as the oocyte, while the remaining cells differentiate as nurse cells to support the oocyte by synthesizing proteins and nutrients [44,45]. As these incomplete divisions occur, CB spectrosomes branch into fusome networks, also marked by 1B1 and spectrins, whose presence indicates the regeneration of the germline after IR. Following IR, the CB population is temporarily depleted but typically recovers fully by four days post IR (dpi) [12,13].
To assess the impact of rapamycin on post-IR regeneration, wild-type (w^1118^) flies (0–3 days old) were conditioned in food vials, supplemented with standard yeast paste, for 48 h. Flies were then irradiated with 50 Gy to induce quiescence. Following IR, we treated flies with either 200 µM rapamycin in 5% EtOH or the solvent control (5% EtOH), dissolved in yeast paste, and analyzed GSC division rates by assessing GSC spectrosome elongation once every 24 h. After 2 dpi, the treatment yeast paste (rapamycin or solvent control) was replaced with standard yeast paste in order to measure germline recovery by 4 dpi (Figure 1B) (Methods). In untreated flies, GSCs enter quiescence at approximately 1 dpi and then exit quiescence at approximately 2 dpi [13,14]. In the germaria of both rapamycin-treated and control-treated flies, the percentage of GSCs with elongated spectrosomes significantly decreased at 1 dpi compared with the unirradiated timepoint, indicating the successful induction of quiescence by IR (Figure 1C–E). At 2 dpi, the GSCs of rapamycin-treated flies continued to have a significantly lower percentage of elongated spectrosomes, compared with the GSCs of control flies at 2 dpi (Figure 1C–E). At 3 dpi, the percentage of GSCs with elongated spectrosomes in the rapamycin-treated flies had returned to a frequency similar to the unirradiated control samples (Figure 1E). Consistent with previous reports [13], we show that rapamycin delays IR-induced quiescence in GSCs, preventing their re-entry into the cell cycle in the wild-type strain, w^1118^ (Figure 1E).
Next, we quantified the percentage of germaria containing branched fusomes (BFs) as an indication of cystoblast division and germline regeneration. The percentage of germaria containing BFs drastically decreases from unirradiated to 1 dpi, indicating the death of somatic cells in the germline as a result of IR (Figure 1C,D,F). After 1 dpi, and until 4 dpi, BF frequency remained low in rapamycin-treated flies. In comparison, the BF frequency of control-treated flies returned to the value prior to irradiation more quickly (Figure 1C,F). At 4 dpi, 66.8% of germaria in control-treated flies contained BFs, while a significantly lower percentage of germaria (47.5%) from rapamycin-treated flies contained BFs (p = 0.02). These data indicate that rapamycin significantly delays the post-IR regeneration of the germline.
3.2. The Effect of Rapamycin Varies Across Genetic Backgrounds
While rapamycin can delay post-IR germline regeneration in the w^1118^ genetic background, which is commonly used in Drosophila melanogaster studies, it is unknown whether this effect is conserved among genetically diverse Drosophila strains. To test the initial hypothesis that rapamycin sensitivity in development corresponds to sensitivity in quiescence, we selected DGRP lines that demonstrated extreme phenotypes of sensitivity or resistance to rapamycin in development as reported in Harrison et al. (2024) (Figure 2A) [36]. The common lab strain, w^1118^, shows moderate sensitivity to rapamycin, delaying pupation by an average of 0.86 days, whereas more sensitive strains show up to a 6-day delay by rapamycin, and resistant strains show little-to-no detectable delay (Figure 2A) [36].
We reproduced the developmental phenotype of DGRP lines screened in Harrison et al. (2024) [36] by measuring the length of 72 h-old larvae, treated with 20 µM rapamycin in 5% EtOH or solvent control (5% EtOH) (Figure 2B, Methods). We recorded the phenotype of w^1118^ and six DGRP lines whose development was highly sensitive to rapamycin (DGRP 443, 287, 517, 776, 57, and 348), and four DGRP lines whose development was relatively resistant (DGRP 307, 383, 441, and 712). Drosophila larvae enter the early third instar stage at about 72 h post fertilization [46]. The early administration of rapamycin in Drosophila leads to a reduced body size of third instar-stage larvae (Figure 2C) [36,47]. We find that the sensitivity to rapamycin greatly varies across the DGRP lines, where rapamycin reduces length by up to half in sensitive strains, and by almost none in resistant strains (Figure 2D). Larval length varies significantly with genotype and treatment, and we detect a significant genotype × treatment interaction (two-way ANOVA, p < 0.001 for all factors). Across these DGRP lines, the reduction in larval size by rapamycin shows a moderately strong correlation with the previously published pupation delay (r_s_ = −0.655, p = 0.029, Supplementary Figure S1B) [30].
Next, we asked whether genetic variation impacts the delay in post-IR germline regeneration by rapamycin. The length of the GSC quiescent period influences the recovery of the germline from injury. Therefore, we observed the extent to which exit from GSC quiescence was delayed. GSCs from w^1118^ flies re-enter the cell cycle at 2 dpi, while GSCs in rapamycin-treated w^1118^ flies do not fully re-enter the cell cycle until 3 dpi (Figure 1). Therefore, we recorded the percentage of GSCs with elongating spectrosomes at 2 dpi in each line treated with rapamycin or solvent control to gauge how genetic variation affects cell cycle re-entry delay by rapamycin. We compared the response of w^1118^ to the responses of the six DGRP lines whose development was highly sensitive to rapamycin and to the four DGRP lines whose development was relatively resistant (Figure 2A). Flies were conditioned in vials with standard food and supplementary yeast paste for 2 days prior to IR. Following IR and treatment with rapamycin or solvent control until 2 dpi, ovaries were dissected to observe spectrosome morphology (Figure 2E).
We observed that rapamycin treatment until 2 dpi prevents cell cycle re-entry at 2 dpi in most lines. However, the magnitude of this delay varies greatly amongst the lines (Figure 2G,H). w^1118^ showed moderate sensitivity, with a log2 fold change in spectrosome elongation of −0.76, with a significant decrease in elongation at 2 dpi in rapamycin-treated flies compared with control-treated flies (p = 0.023). Several other DGRP lines exhibited fold changes similar to w^1118^, such as DGRP 776 (−0.66) and 348 (−0.65). DGRP 517, whose developmental timing is highly sensitive to rapamycin, had a low fold change of −2.36 (Figure 2H), with a highly significant decrease in elongation at 2 dpi in rapamycin-treated flies compared with the control (p = 0.001) (Figure 2I). DGRP 441, whose developmental timing is highly resistant to rapamycin, had a positive fold change of 0.11 (Figure 2H) with no significant difference in elongation between treatments at 2 dpi (p = 0.529) (Figure 2I).
Sensitivity to rapamycin in GSC quiescence shows a moderately strong positive correlation with the effect on larvae length (r_s_ = 0.682, p = 0.021) (Figure 2J). However, comparing the sensitivity of GSC quiescence to pupation delay reported in Harrison et al. (2024) [36], the degree of sensitivity to rapamycin in pupation delay does not significantly correlate with sensitivity in quiescence exit (r_s_ = 0.4, p = 0.223, Supplementary Figure S2B). Among the lines that were the most sensitive in pupation delay (Figure 2A), the lines 287 (−0.5), 443 (−0.49), and 776 (−0.66) appeared relatively resistant in quiescence. DGRP 383, on the other hand, was resistant in pupation delay (Figure 2A), but was among the more sensitive lines to rapamycin in the GSC quiescence response (fold change = −1.68, Figure 2H). We show that there is a significant, moderately strong correlation between the previously published pupation delay and larval length (Supplementary Figure S1B). However, the lack of correlation between pupation delay and quiescence exit suggests that the effect of rapamycin not only depends on genetic background, but also on the biological context.
3.3. Germline Regeneration Varies with Rapamycin Across Genetic Backgrounds
We have shown that genetic variation influences the extension of quiescence by rapamycin in GSCs. Next, we asked if the variation in sensitivity to rapamycin in quiescence is also associated with variation in the capacity for post-IR regeneration. Among the DGRP lines screened for rapamycin sensitivity in IR-induced quiescence previously, DGRP 517 was the most sensitive to rapamycin, while DGRP 441 was the most resistant (Figure 2H).
We first assayed how rapamycin impacts the capacity for post-injury regeneration in these two DGRP lines, along with w^1118^ for comparison, by tracking spectrosome elongation rates throughout the duration of recovery (until 4 dpi). Conditioning, irradiation, treatment, and dissection were performed, and spectrosome elongation rates were quantified. GSCs from both lines, regardless of rapamycin or control treatment, entered quiescence as evidenced by the drop in spectrosome elongation at 1 dpi (Figure 3A–D). However, a higher percentage of GSCs had elongated spectrosomes in DGRP 441 at 1 dpi (12.9% and 17.9% for control and rapamycin, respectively) compared with DGRP 517 (6.0% and 3.1% for control and rapamycin, respectively), suggesting that IR is less likely to induce quiescence in the GSCs of DGRP 441 than DGRP 517.
GSCs from DGRP 441 flies in both treatments exited quiescence by 2 dpi, as evidenced by the increase in spectrosome elongation (22.85% and 26.24% for control-treated and rapamycin-treated flies, respectively). In DGRP 441, there was no significant difference in percent GSCs with elongated spectrosomes between the treatments across all timepoints (Figure 3D). As observed formerly (Figure 2H,I), GSCs from DGRP 517 flies treated with rapamycin stayed in quiescence at 2 dpi while GSCs from those treated with the solvent control re-entered the cell cycle (Figure 3C). Unlike DGRP 441 and w^1118^, spectrosome elongation rates in rapamycin-treated DGRP 517 flies did not equate to those of the control-treated flies until 4 dpi (p = 0.308). We therefore affirm that the recovery of GSC division post IR is compromised by rapamycin in the sensitive line.
Next, we assessed how the regeneration of the germline varied in response to rapamycin by quantifying BF rates. BF rates in germaria from rapamycin-treated DGRP 441 flies were significantly lower at 3 dpi compared with control-treated flies, despite there being no significant difference in spectrosome elongation rates across all timepoints. However, by 4 dpi, the germline of DGRP 441 was fully regenerated, with 81.9% and 82.4% of germaria having BF for control-treated and rapamycin-treated flies, respectively, with no significant difference between the percent total recovery rates between treatments (p = 0.705). DGRP 517, on the other hand, showed a significant decrease in BF rates at 3 dpi and 4 dpi in the germaria of rapamycin-treated flies, compared with control-treated flies at those timepoints, with a significant difference in percent of total recovery at 4 dpi between the treatments (p = 0.041). Surprisingly, in both treatments of DGRP 517, the germaria do not fully recover despite there being high spectrosome elongation rates, suggesting that the cystoblast division does not fully recover after irradiation. Here we find that rapamycin significantly delays the post-IR germline regeneration of a sensitive line, but not a resistant one.
3.4. Genetic Variation in Mitophagy and Not DNA Repair During Quiescence
We then tested whether genetic variation in the response to rapamycin involves either of two repair mechanisms required to exit quiescence: DNA repair or the removal and replacement of damaged mitochondria. Drosophila GSCs show evidence of DNA damage at 30 min post IR which is fully repaired by 24 h post IR [13]. We tested whether the timing of IR-induced DNA-DSB repair differs between the highly sensitive DGRP 517 line and the highly resistant DGRP 441 line when treated with rapamycin, and in comparison, to the moderately sensitive genotype w^1118^. If rapamycin extends quiescence by impairing DNA damage repair, we should see that the DNA damage accumulation pattern of each line with rapamycin parallels the duration of quiescence.
To estimate the level of DNA damage, we measured the intensity of phospho-H2Av (anti-γH2Av), a marker for DNA-DSB in Drosophila (Methods). All lines and treatments showed an increase in γH2Av intensity from their unirradiated condition to 30 min post IR (30 mpi, Figure 4A,B). In DGRP 517 and 441, both unirradiated and irradiated flies had a higher percentage of GSCs with high-level DNA damage compared with w^1118^, indicating that both DGRP lines have a higher sensitivity to IR (Figure 4B,D,E). In control-treated w^1118^ flies, all GSCs at 1 dpi had low-level γH2Av, suggesting they had already repaired their DNA. In contrast, at 1 dpi in rapamycin-treated w^1118^ flies, 32% of GSCs still had moderate-level DNA damage. At 1 dpi, 91% and 80% of GSCs in control-treated DGRP 517 and 441 flies, respectively, showed moderate DNA damage. In the same lines, GSCs from rapamycin-treated flies at 1 dpi show similar percentages of moderate DNA damage (88% for DGRP 517 and 75% for DGRP 441), suggesting that rapamycin does not have a substantial effect on DNA repair in these lines. The similarity in DNA damage between rapamycin-treated and control flies of DGRP 517 and 441 suggests that DNA damage repair does not contribute to the substantial difference in the duration of quiescence in response to rapamycin in these lines.
Next, we asked whether sensitive and resistant lines differ in their ability to remove and replace damaged mitochondria when treated with rapamycin. In Drosophila GSCs, the entry into quiescence requires mitochondrial fission and mitophagy to remove dysfunctional mitochondria. Re-entry into the cell cycle requires mitochondrial biogenesis and fusion to reconstruct the mitochondrial network [14,48,49,50]. Actively proliferating stem cells often feature a tubular mitochondrial network localized to the cell anterior [12,41,42,43,44,45,46,47,48,49,50,51,52,53]. In contrast, the mitochondrial network of quiescent cells, when mitophagy is active, appears fragmented and is diffused over a larger part of the cell [12,51,54]. We gauged the rate of mitochondria removal and replacement by quantifying the number of GSCs with diffuse localization of the mitochondrial marker ATP β-Synthase (Methods).
Following irradiation, we observed an increase in the percentage of GSCs with fragmented mitochondria at 1 dpi across all genotypes and treatments, signifying active mitophagy (Figure 5A,D–F). However, post 1 dpi fragmentation rates varied drastically across genotypes in response to rapamycin. In DGRP 441, rapamycin did not have a significant impact on mitochondrial fragmentation, with no significant difference observed in fragmentation between rapamycin-treated and control-treated flies across all timepoints (Figure 5F). In both treatments of DGRP 441, the fragmentation rates resolved to the levels prior to irradiation by 2 dpi.
In contrast, rapamycin significantly impacted fragmentation rates past 1 dpi in the lines whose quiescence was sensitive to rapamycin. In DGRP 517 and w^1118^, but not in DGRP 441 flies, GSC mitochondrial fragmentation was significantly prolonged due to rapamycin treatment beyond 1 dpi (DGRP 517, p = 0.008; w^1118^, p = 0.009). These data suggest that mitochondrial fragmentation correlates with rapamycin sensitivity.
Unlike the resolution of DNA damage that followed IR among flies of both the highly sensitive line DGRP 517 and the resistant line DGRP 441 (Figure 4), we find that prolonged mitochondrial fragmentation associates with the sensitivity to rapamycin in GSC quiescence among these lines. These data suggest that genes involved with increased mitophagy or a lack of mitochondrial biosynthesis are plausible culprits for the rapamycin sensitivity differences between genetic backgrounds.
4. Discussion
Adult, embryonic and cancer diapause-like stem cells enter a reversible quiescence upon radiation stress to resolve damage before regenerating lost tissue. Across stem cell types, the repression of mTORC1 characterizes stem cell quiescence, and its activation promotes cell cycle re-entry [21,22,23,24,25,26]. We have shown that the mTORC1-specific inhibitor rapamycin prevents stem cells from exiting quiescence and delays regeneration in the Drosophila female germline. We now show that the effect of rapamycin on quiescence and regeneration varies greatly with genetic background. We analyze this variation on a mechanistic level and show that rapamycin sensitivity in quiescence is closely attributed to differences in mitochondrial dynamics.
mTORC1 signaling is fundamental to growth and development in a wide range of species, and there are a growing number of cellular and developmental mechanisms that are affected by mTORC1 [55,56]. The effects of rapamycin, therefore, may be highly pleiotropic, with a range of downstream responses that may differ with rapamycin across genetic backgrounds. We have revealed that the genetic variation in the response to rapamycin that appears among the wild-derived DGRP is not consistent across the phenotypes affected by rapamycin (Supplementary Figure S2). Rapamycin both reduces larval size and prolongs the time required for larvae to form pupae [41,42], and both effects vary strongly by genetic background [36]. By sampling the DGRP lines from those screened for the effect of rapamycin on development time, we were able to test the hypothesis that genetic variation in the effect of rapamycin affects mTORC1 activity generally across phenotypes, such as the pace of development and stem cell quiescence. Consistent with this hypothesis, we found that the effect of rapamycin on quiescence correlates with the rapamycin sensitivity of larval growth among these lines (Figure 2J). In contrast to this hypothesis, however, we found that the extent of developmental delay induced by rapamycin reported in Harrison et al. (2024) [36] did not strictly correlate with the variation in the duration of quiescence that we measured (Supplementary Figure S2B). The difference in the response to rapamycin between genotypes in development, therefore, appears to affect different mechanisms than those that affect the duration of quiescence. Thus, rapamycin sensitivity can be defined by line-specific variation in mTORC1 effector pathways, driven by genetic diversity.
After establishing that sensitivity to rapamycin in IR-induced quiescence varies across the DGRP, we probed the mechanism that underlies this sensitivity by considering two requirements of quiescence exit: DNA damage repair and the removal and replacement of damaged mitochondria. We compared these processes across w^1118^ and two DGRP lines at the extreme ends of the quiescence sensitivity distribution, DGRP 517 and DGRP 441. In DGRP 517 and w^1118^, whose quiescence was sensitive to rapamycin, we see an increase in mitochondrial fragmentation past 1 dpi, where fragmentation takes longer to resolve in response to rapamycin than in the control. But in DGRP 441, the resistant line with a normal duration of quiescence with rapamycin, mitophagy appeared to resolve more quickly (Figure 5)
In contrast to the variation in the persistence of mitophagy, we found no evidence for a differential effect of rapamycin on the timing of DNA-DSB repair in w^1118^, compared with DGRP 441 and DGRP 517. Therefore, the genetic variation associated with prolonged quiescence does not appear to affect DNA repair. Our results suggest that the repair or replacement of mitochondria is sensitive to genetic background. We propose that the mechanism underlying rapamycin sensitivity in quiescence is mitophagy or mitochondrial biogenesis. The DGRP lines we tested house tens of thousands of SNP differences [37]. Future work will characterize the genetic basis of observed sensitivity in quiescence. In particular, it will be important to dissect whether differences in the resolution of fragmented mitochondria arise from SNPs in genes regulating mitophagy or mitochondrial biogenesis.
Rapamycin and its analogs (rapalogs) hold significant promise as anti-cancer stem cell therapeutics yet pronounced variability in their response limits their efficacy. Our findings uncover a previously unrecognized mechanism underlying this resistance and identify mitochondrial regulation as a tractable vulnerability. These results provide a strong rationale for testing combinatorial strategies that pair rapalogs with agents targeting mitochondrial autophagy or biogenesis to enhance therapeutic response.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fuchs E. Blau H.M. Tissue Stem Cells: Architects of Their Niches Cell Stem Cell 20202753255610.1016/j.stem.2020.09.01133007238 PMC 7861346 · doi ↗ · pubmed ↗
- 2Biteau B. Hochmuth C.E. Jasper H. Maintaining tissue homeostasis: Dynamic control of somatic stem cell activity Cell Stem Cell 2011940241110.1016/j.stem.2011.10.00422056138 PMC 3212030 · doi ↗ · pubmed ↗
- 3Oh J. Lee Y.D. Wagers A.J. Stem Cell Aging: Mechanisms, Regulators and Therapeutic Opportunities Nat. Med.20142087088010.1038/nm.365125100532 PMC 4160113 · doi ↗ · pubmed ↗
- 4Behrens A. van Deursen J.M. Rudolph K.L. Schumacher B. Impact of Genomic Damage and Ageing on Stem Cell Function Nat. Cell Biol.20141620120710.1038/ncb 292824576896 PMC 4214082 · doi ↗ · pubmed ↗
- 5Davalli P. Mitic T. Caporali A. Lauriola A. D’Arca D. ROS, Cell Senescence, and Novel Molecular Mechanisms in Aging and Age-Related Diseases Oxid. Med. Cell. Longev.20162016356512710.1155/2016/356512727247702 PMC 4877482 · doi ↗ · pubmed ↗
- 6Martínez-Reyes I. Chandel N.S. Mitochondrial TCA Cycle Metabolites Control Physiology and Disease Nat. Commun.20201110210.1038/s 41467-019-13668-331900386 PMC 6941980 · doi ↗ · pubmed ↗
- 7Ishibashi J.R. Keshri R. Taslim T.H. Brewer D.K. Chan T.C. Lyons S. Mc Manamen A.M. Chen A. Del Castillo D. Ruohola-Baker H. Chemical Genetic Screen in Drosophila Germline Uncovers Small Molecule Drugs That Sensitize Stem Cells to Insult-Induced Apoptosis Cells 202110277110.3390/cells 1010277134685753 PMC 8534514 · doi ↗ · pubmed ↗
- 8Urbán N. Cheung T.H. Stem Cell Quiescence: The Challenging Path to Activation Development 2021148 dev 16508410.1242/dev.16508433558315 PMC 7888710 · doi ↗ · pubmed ↗