The Physiological Significance of TRP and Piezo Channels as Physical Stimulus Sensors in Brown Adipocytes
Kunitoshi Uchida, Mari Iwase

TL;DR
This review explains how TRPV2 and Piezo1 ion channels help regulate brown fat cell development and heat production through calcium signaling.
Contribution
The paper introduces the concept of a 'modal shift' in TRP/Piezo channels during brown adipocyte development.
Findings
TRPV2 and Piezo1 are key regulators of brown adipocyte differentiation and thermogenesis.
Ion channel function changes during different stages of brown adipocyte development.
Cation-channel-mediated calcium signaling is crucial for thermogenesis and metabolic balance.
Abstract
What are the main findings? This review highlights Ca2+-permeable ion channels, particularly TRPV2 and Piezo1, as important regulators of brown adipocyte differentiation and thermogenesis.Recent evidence supports the concept of a stage-dependent shift in ion channel function during brown adipocyte differentiation, which may involve changes in channel sensitivity and regulatory mechanisms. This review highlights Ca2+-permeable ion channels, particularly TRPV2 and Piezo1, as important regulators of brown adipocyte differentiation and thermogenesis. Recent evidence supports the concept of a stage-dependent shift in ion channel function during brown adipocyte differentiation, which may involve changes in channel sensitivity and regulatory mechanisms. What are the implications of the main findings? These findings clarify the role of Ca2+ signaling in brown adipocyte responses to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
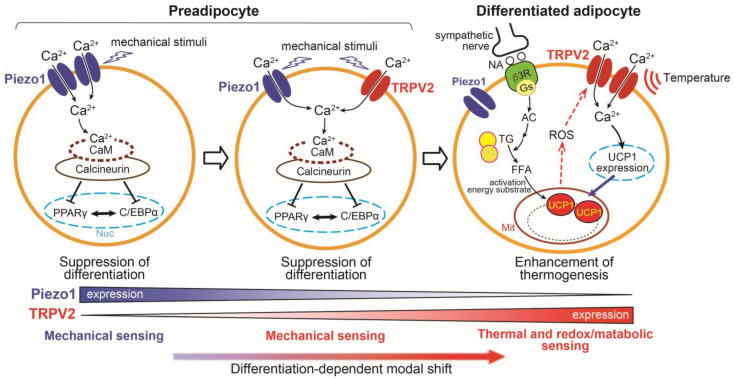 Figure 1
Figure 1- —Takeda Science Foundation
- —JSPS KAKENHI
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsIon Channels and Receptors · Calcium signaling and nucleotide metabolism · Erythrocyte Function and Pathophysiology
1. Introduction
1.1. Brown and White Adipose Tissues
Adipose tissue is a highly plastic organ that dynamically adapts to environmental, nutritional, and neural cues to maintain systemic energy homeostasis [1]. In mammals, adipose tissue is broadly categorized into white adipose tissue (WAT) and brown adipose tissue (BAT), with an additional population of recruitable brown-like adipocytes, called beige (or brite) adipocytes, emerging within subcutaneous WAT depots under specific physiological conditions [2]. While WAT’s primary function is to store energy, BAT is specialized for energy expenditure. A defining functional property of BAT is Uncoupling protein 1 (UCP1)-dependent non-shivering thermogenesis, which dissipates the proton gradient generated by oxidative phosphorylation as heat [3,4]. This thermogenesis is initiated by sympathetic nervous system activation, typically in response to cold exposure or gastrointestinal chemical stimuli such as pungent compounds, leading to noradrenaline release and subsequent β-adrenergic receptor signaling in brown adipocytes [5]. The high mitochondrial density and dense sympathetic innervation of BAT make this tissue particularly well suited for stimulus-responsive energy dissipation. Beige adipocytes share key functional features with classical brown adipocytes, including inducible UCP1 expression and thermogenic capacity, although their developmental origin and transcriptional profiles are different from brown adipocytes [6,7]. Recruitment of beige adipocytes is triggered by sustained cold exposure or pharmacological β3-adrenergic receptor activation, highlighting the importance of environmental and neural inputs in shaping and determining adipose tissue phenotypes.
The presence of metabolically active BAT in adult humans, once considered minimal, has been firmly established by fluorodeoxyglucose positron emission tomography–computed tomography (FDG-PET/CT) imaging studies [8,9,10]. Histological and transcriptomic analyses have identified UCP1-positive adipocytes and heterogeneous adipocyte populations with brown- and beige-like characteristics in human adipose depots [2,11]. Importantly, BAT abundance and activity are inversely correlated with adiposity-related parameters, including body fat mass and body mass index (BMI) [8,12], highlighting the contribution of thermogenic adipocytes to whole-body energy balance. Consistent with these observations, experimental models demonstrate that sustained activation of BAT thermogenesis through prolonged cold exposure or β3-adrenergic stimulation increases BAT mass and reduces adiposity [13], whereas genetic impairment of thermogenic capacity, such as in UCP1-deficient mice, leads to obesity in animals under chronic high-fat feeding conditions [14]. Collectively, these findings indicate that BAT functions as a highly stimulus-responsive metabolic tissue, and that disruption of its adaptive thermogenic capacity can contribute to metabolic imbalance. The general features of white, brown, and beige adipocytes have been described elsewhere; here, attention is directed to stimulus-responsive mechanisms in brown and beige adipocytes, particularly those involving Ca^2+^ signaling and ion channels.
1.2. The Mechanism of Thermogenesis in Brown and Beige Adipocytes
Intracellular Ca^2+^ signaling has been implicated in the regulation of adipocyte differentiation and thermogenic activation. In brown adipocytes, β3-adrenergic receptor signaling promotes mitochondrial uncoupling via UCP1, resulting in dissipation of the proton gradient across the inner mitochondrial membrane and the conversion of chemical energy into heat [15]. Long-chain fatty acids generated by lipolysis and redox-dependent modifications of UCP1 have been proposed as important contributors to this process [16,17]. Along with these processes, Ca^2+^ signaling has been shown to influence adipocyte differentiation. In preadipocytes, increases in intracellular Ca^2+^ concentration ([Ca^2+^]i) suppress differentiation through calcineurin-dependent pathways and inhibition of insulin signaling, as demonstrated in 3T3-L1 cells [18,19,20]. These findings indicate that Ca^2+^ signaling may exert distinct effects depending on the differentiation states of adipocytes.
In differentiated brown and beige adipocytes, Ca^2+^ dynamics appear to participate in the regulation of thermogenesis. Changes in [Ca^2+^]i have been linked to UCP1 expression and whole-body energy expenditure [21]. In addition, ATP-dependent Ca^2+^ cycling mediated by Sarco/endoplasmic reticulum Ca^2+^-ATPase 2b (SERCA2b) and Ryanodine receptor 2 generates UCP1-independent heat in beige adipocytes [22], whereas membrane hyperpolarization that limits Ca^2+^ influx impairs adrenergic thermogenesis in brown adipocytes [23]. More recently, adrenergic stimulation has been shown to promote the formation of a mitochondrial complex containing the Mitochondrial Ca^2+^ uniporter (MCU), its regulatory component (Essential MCU regulatory element; EMRE), and UCP1, thereby enhancing mitochondrial Ca^2+^ uptake and supporting uncoupled respiration [24]. Despite these advances, how Ca^2+^ influx is initiated, coordinated, and tuned in adipocytes under physiological conditions remains poorly understood. This suggests that Ca^2+^-permeable ion channels, including transient receptor potential (TRP) and Piezo channels, may act as upstream regulators connecting physiological stimuli that induce Ca^2+^ signaling to adipocyte differentiation and thermogenesis.
1.3. Transient Receptor Potential Channels
TRP channels comprise a large family of Ca^2+^-permeable ion channels, most of which are non-selective [25]. Structurally, TRP channels share a conserved architecture consisting of six transmembrane domains (TM1-TM6) and a pore-forming loop between TM5 and TM6, with both N- and C-termini located in the cytosol [26]. In mammals, the TRP superfamily comprises six subfamilies: TRPV (Vanilloid), TRPC (Canonical), TRPM (Melastatin), TRPML (Mucolipin), TRPP (Polysystin), and TRPA (Ankyrin), encompassing a total of 28 channels [27,28]. Functionally, TRP channels act as polymodal sensors that are activated by diverse physical and chemical stimuli, including temperature, mechanical force, endogenous lipids, oxidants, ions, and bioactive compounds [29,30]. Among these, several TRP channels exhibit temperature-dependent activation within physiologically relevant ranges; they are collectively referred to as thermo-sensitive TRP channels (thermo-TRPs). Eleven thermo-TRPs have been identified in mammals, primarily belonging to the TRPV, TRPM, TRPA, and TRPC subfamilies. TRPV1, TRPV2, and TRPM3 respond to elevated temperatures, whereas TRPM8, TRPA1, and TRPC5 are activated by cool or cold stimuli, although the cold sensitivity of mammalian TRPA1 remains controversial. TRPV3, TRPV4, TRPM2, TRPM4, and TRPM5 are activated by warm temperatures [31].
Importantly, TRP channels are expressed not only in sensory neurons and skin but also in a wide range of non-sensory tissues that are not directly exposed to rapid environmental temperature changes. In such contexts, TRP channels are thought to function as integrators of cellular signaling downstream of physiological stimuli. Notably, multiple TRP channels are expressed in tissues involved in energy intake and expenditure, including the hypothalamus, gastrointestinal tract, peripheral sensory neurons, liver, and adipose tissue [32,33,34]. Growing evidence indicates that TRP channels in adipocytes contribute to the regulation of differentiation, thermogenesis, and metabolic function, largely through Ca^2+^-dependent mechanisms [35,36].
1.4. Piezo Channels
Piezo channels were identified in 2010 as mechanically activated non-selective cation channels through an RNAinterference screen targeting stretch-evoked currents in mammalian cells [37]. Structural analyses have revealed that Piezo proteins assemble as large trimeric complexes forming a three-bladed propeller-like architecture with a peripheral curved membrane footprint that is well suited for sensing mechanical forces [38]. Two members of the Piezo family are present in mammals: Piezo1 and Piezo2. Although both channels act as mechanotransducers, their expression patterns and physiological roles differ. Piezo1 is mainly expressed in non-sensory tissues and functions as a shear-stress sensor in vascular endothelium and a detector of pressure changes in organs such as the bladder [39,40]. In contrast, Piezo2 is predominantly expressed in sensory neurons, including dorsal root ganglion neurons, and Merkel cells, and plays essential roles in touch sensation, proprioception, and mechanical pain [41,42].
Piezo channels are increasingly recognized as regulators of diverse biological processes, such as vascular development, bone formation, immune function, and respiratory control [43]. Piezo1, in particular, integrates extracellular mechanical cues with Ca^2+^ signaling to influence cell fate, proliferation, and differentiation. Emerging evidence indicates that Piezo1 is expressed in metabolic tissues, including adipose tissue, and may participate in adipocyte differentiation, adipose remodeling, and systemic metabolic regulation, suggesting additional roles for Piezo channels beyond somatosensation.
While our previous reviews summarized the roles of TRP channels in systemic energy metabolism and thermoregulation, less attention has been paid to how the functional stimuli and signaling roles of these channels change across adipocyte developmental stages [35,44]. In this review, we focus on the concept of A developmental-stage-dependent “modal shift” in TRPV2 and Piezo1 functions in brown adipocytes, highlighting how identical Ca^2+^-permeable channels can exert distinct effects on differentiation versus thermogenic activation depending on expression levels, activation stimuli, and cellular expression levels.
2. Functional TRP/Piezo Channels in Brown Adipocytes
Although TRP channels have been well studied in primary afferent neurons, particularly TRPV1, in the vagal sensory pathways involved in dietary sensing, increasing evidence indicates that several TRP and Piezo channels are also expressed in brown adipose tissue and brown adipocytes, and their physiological functions have been elucidated (Table 1). In the following sections, we focus on the physiological roles of TRPV2 and Piezo1 in brown adipocytes, which have been examined in detail by our group and others, and we also summarize recent findings on other TRP channels expressed in brown and beige adipocytes.
2.1. TRPV2
TRPV2 is a Ca^2+^-permeable non-selective cation channel that is activated by noxious heat (>52 °C) and mechanical stimuli such as membrane stretch. TRPV2 is expressed in neurons, the heart, and immune cells and shows particularly high expression in brown adipose tissue [51]. In undifferentiated brown adipocytes, functional TRPV2 expression is low but increases during differentiation, suggesting that TRPV2 plays important roles in differentiated brown adipocytes.
In BAT from systemic TRPV2 knockout (TRPV2KO) mice, brown adipocytes and lipid droplets are enlarged, and whitening-like morphological changes are observed. Consistent with these phenotypes, thermogenic genes including UCP1 are downregulated, whereas genes associated with lipid storage tend to be upregulated, suggesting an imbalance in BAT energy metabolism. Physiologically, TRPV2KO mice show a greater reduction in core body temperature during 4 °C cold exposure, without changes in locomotor activity or sympathetic nerve activity, compared with wild-type mice. Moreover, the β3-adrenergic agonist–induced increase in interscapular BAT temperature is blunted in TRPV2KO mice under anesthesia [51].
In contrast, whole-body oxygen consumption, locomotor activity, and food intake do not differ between TRPV2KO and wild-type mice under standard housing conditions. This observation suggests that alterations in TRPV2-dependent thermogenic phenotypes may not be reflected in basal energy expenditure under standard housing conditions [51].
In isolated mouse primary brown adipocytes, the induction of UCP1 gene expression by treatment with β-adrenergic agonists is also almost abolished in brown adipocytes from TRPV2KO mice. Similarly, induction of UCP1 expression and non-esterified fatty acid release by an adenylyl cyclase (AC) activator, forskolin, is impaired in TRPV2-deficient cells, and chelation of intracellular Ca^2+^ by BAPTA-AM suppresses the β-adrenergic receptor activation-induced upregulation of UCP1. These observations support the idea that TRPV2-mediated Ca^2+^ influx is required for full β-adrenergic induction of thermogenic gene expression, although the precise position of TRPV2 relative to AC/cAMP signaling remains to be determined and could be influenced by additional factors such as changes in redox state or local temperature, as discussed below. In addition, other TRPV2-activating inputs, including mechanical stimulation (membrane stretch), bioactive lipids such as lysophospholipids and endocannabinoids [64,65,66,67], or growth factor–dependent membrane recruitment of TRPV2, cannot be excluded as contributing mechanisms [68,69]. These findings suggest that TRPV2 in differentiated brown adipocytes contributes to non-shivering thermogenesis (Figure 1) [51].
When obesity is induced by feeding a high-fat diet (HFD), TRPV2KO mice develop more severe obesity than wild-type mice. While systemic deletion of TRPV2 does not exclude the contribution of other tissues, this phenotype suggests that TRPV2-dependent thermogenic mechanisms contribute to whole-body energy expenditure. Human brown adipocytes are thought to resemble beige adipocytes, which are induced within white adipose depots upon cold exposure, rather than classical rodent brown adipocytes [70]. Cold-induced recruitment of beige adipocytes is reduced in TRPV2KO mice [51], implying that TRPV2 may also play important roles in human brown/beige adipocytes.
As noted above, TRPV2 is already detectable in preadipocytes, although its expression is low and markedly increases during adipocyte differentiation. When a TRPV2 agonist is applied from the onset of differentiation, adipocyte differentiation is suppressed. TRPV2 activation reduces the expression of Peroxisome proliferator-activated receptor γ (PPARγ), a key transcription factor for adipocyte differentiation, without suppressing brown adipocyte-specific genes such as PPARγ coactivator 1 α (PGC-1α) and PR domain containing 16 (PRDM16). Mechanical stimulation likewise inhibits brown adipocyte differentiation, and pharmacological inhibition of TRPV2 suggests that this effect is mediated, at least in part, by TRPV2 activation. Ca^2+^ influx is known to inhibit white adipocyte differentiation through Ca^2+^/calmodulin-dependent activation of calcineurin, and differentiation of brown adipocytes is similarly suppressed by TRPV2 activation. This inhibition is partially reversed by calcineurin inhibitors [52]. Consistent with this idea, recent work showed that selective TRPV2 inhibitors derived from natural coumarin enantiomers reverse TRPV2 agonist-induced suppression of brown adipocyte differentiation, providing pharmacological support for TRPV2-mediated Ca^2+^ entry as a negative regulator of adipogenesis [71]. Thus, TRPV2 exerts stage-dependent functions during the brown adipocyte differentiation process (Figure 1).
Although endogenous activators of TRPV2 in brown adipocytes remain incompletely defined, several candidate stimuli have been proposed, including membrane stretch, bioactive lipid mediators, and Insulin growth factor 1 (IGF-1)–induced membrane recruitment of TRPV2. Recent work has further shown that oxidation of methionine residues at positions 528 and 607 markedly lowers the temperature threshold for TRPV2 activation toward physiological ranges [72]. Consistent with these findings, treatment with a methionine oxidant (chloramine-T) activates TRPV2 at ~30 °C in TRPV2-expressing HEK293T cells and in differentiated brown adipocytes, and potentiates the expression of thermogenesis-related genes [53]. These observations raise the possibility that post-translational modifications, such as methionine oxidation, contribute to TRPV2 sensitivity to intracellular and/or tissue temperature and redox conditions. Given that mitochondrial reactive oxygen species increase during elevated thermogenic activity, such redox changes could be linked to metabolic demand and participate in the modulation of TRPV2-dependent thermogenic responses (Figure 1). However, it should be noted that chloramine-T has low specificity, and its effects on thermogenic gene expression may reflect oxidative stress signaling beyond TRPV2-specific activation. Further clarification is required to determine whether changes in thermogenic gene expression are attributable to TRPV2-specific signaling or secondary responses associated with oxidative stress.
2.2. Piezo1 Channel
Mechanical stimulation during the differentiation of brown adipocytes suppresses differentiation, and functional TRPV2 expression is low in brown preadipocytes. These observations suggested that other mechano-sensitive ion channels may be involved. In mouse brown preadipocytes, Piezo1 is strongly expressed, whereas its functional expression decreases during the progressive differentiation, in contrast to TRPV2. Activation of Piezo1 suppresses brown adipocyte differentiation, similar to the effect of TRPV2 activation. Piezo1 activation also reduces the expression of PPARγ, without suppressing brown adipocyte–specific genes such as PGC-1α and PRDM16. This suppression was completely reversed by a calcineurin inhibitor (FK506), and the enhancement of calcineurin activity by the Piezo1 agonist Yoda-1 is completely abolished by silencing Piezo1 in preadipocytes. Together, these findings suggest that Piezo1-mediated increase in intracellular Ca^2+^ activates calcineurin, thereby inhibiting adipocyte differentiation (Figure 1) [61]. Conversely, the knockdown of Piezo1 via siRNA promoted differentiation under static culture conditions without the application of external mechanical stimulation or a Piezo1 agonist. Although these in vitro conditions do not quantitatively mimic the mechanical environment of adipose tissue in vivo, this observation supports the notion that Piezo1 acts as a sensor for basal or low-level mechanical inputs, including cell movement, during adipocyte differentiation.
Piezo1 is also expressed in brown adipose tissue, and Piezo1 protein is detectable in brown adipocytes containing lipid droplets [61], indicating the possibility that Piezo1 also exerts functional roles in differentiated brown adipocytes. Its expression is elevated in the adipose tissue of obese mice. Adipocyte-specific Piezo1KO mice exhibit insulin resistance and impaired adipose remodeling when fed with a HFD, together with adipocyte hypertrophy and increased inflammatory tone in perigonadal white adipose tissue (pgWAT) [62]. In addition, analyses of adipocyte-specific Piezo1 knockout mice show that activation of Piezo1 in differentiated white adipocytes induces secretion of fibroblast growth factors (FGFs), which act on progenitor cells to promote their differentiation [63]. Lack of Piezo1 in differentiated white adipocytes impaired preadipocyte-to-adipocyte differentiation under HFD feeding, a defect accompanied by adipocyte hypertrophy, elevated white adipose tissue inflammation, and reduced insulin sensitivity [63]. Although direct evidence in brown adipocytes is completely lacking, it is possible that a similar Piezo1-dependent FGF signaling mechanism, from differentiated brown adipocytes to preadipocytes, could also operate in BAT.
2.3. Other TRP Channels
Recent studies implicated TRPV1 in adipocyte biology. TRPV1 expression, activated by dietary capsaicin and heat, has been reported in the 3T3-L1 adipocyte cell line, and its activation induces a Ca^2+^ influx that suppresses adipogenesis, likely via calcineurin-dependent mechanisms [45]. TRPV1 activation also enhances thermogenic gene expression and promotes browning during adipocyte differentiation [48], although TRPV1 inhibition has been reported to increase PGC-1α expression, indicating context-dependent regulation. Beyond effects on differentiation, TRPV1 appears linked to thermogenic capacity. Capsaicin treatment elevates intracellular and mitochondrial Ca^2+^ handling and stimulates both UCP1-dependent and ATP-dependent thermogenesis through β-adrenergic and Ca^2+^-dependent pathways [49]. TRPV1 has also been detected in a subset of brown preadipocytes in mice. A population of vascular smooth muscle–derived adipocyte progenitors expressing TRPV1 expands in response to cold and differentiates into thermogenic adipocytes [46]. Moreover, combined loss of TRPV1 and UCP1 results in severe impairment of BAT mitochondrial respiration and thermogenesis, suggesting that TRPV1 may contribute to mitochondrial Ca^2+^ regulation in brown adipocytes. Collectively, these findings support the role of TRPV1 in the regulation of adipocyte differentiation and thermogenic function, although the physiological modes of TRPV1 activation in adipocytes require further clarification.
TRPV4 is expressed in both brown and white adipose tissues [54]. In the white adipocyte cell line 3T3-F442A, knockdown of TRPV4 induces the expression of thermogenic genes such as UCP1, whereas TRPV4 activation leads to phosphorylation of ERK and JNK and suppresses thermogenic gene expression. Consistent with these findings, TRPV4 knockout mice are resistant to HFD-induced obesity, and pharmacological inhibition of TRPV4 promotes browning of white adipose tissue [54]. In contrast, adipose-specific overexpression of TRPV4 has also been reported to protect mice against diet-induced obesity and to promote white fat browning via activation of the AKT pathway. This phenotype was accompanied by reduced adipose inflammation and improved insulin sensitivity [55]. Together, these observations suggest that the metabolic impact of TRPV4 signaling depends on cellular context and signaling state. Notably, TRPV2KO and TRPV4KO mice display opposite phenotypes in response to high-fat feeding, indicating that the role of Ca^2+^ signaling in adipocytes is highly channel- and context-dependent, including the timing of channel activation during differentiation or metabolic challenge.
TRPM8 has been reported to be expressed in mouse brown adipose tissue. TRPM8 activation by menthol increases UCP1 expression in brown adipose tissue via protein kinase A activation [58]. In addition, the activation of TRPM8 in human white adipocyte cell lines induces the browning of white adipocytes [60]. More recently, menthol pretreatment of the skin is shown to enhance cold tolerance and mitigate cold injury in mice via TRPM8-dependent activation of BAT thermogenesis, further supporting the physiological role of TRPM8 in adaptive heat production [59]. These findings raise the possibility that, at least in vivo, TRPM8 expressed in sensory neurons may contribute to the modulation of BAT thermogenesis.
Collectively, these findings suggest that multiple TRP channels contribute to the regulation of the adipocyte phenotype and thermogenesis. These seemingly opposing phenotypes likely reflect differences in cell type, differentiation stage, stimulus intensity and duration, and systemic and adipocyte-autonomous effects.
3. Conclusions and Perspectives
Most TRP and Piezo channels are Ca^2+^-permeable, non-selective cation channels that regulate cellular excitability and elicit downstream signaling responses through Ca^2+^ influx. In brown adipocytes, the expression and functional contributions of these channels change during development. Piezo1 is highly expressed in preadipocytes, where it is thought to sense mechanical inputs arising from cell movement, volume changes, or lipid accumulation, thereby acting as a negative regulator of differentiation through Ca^2+^/calcineurin signaling. As differentiation proceeds, Piezo1 expression declines, whereas TRPV2 expression increases, and TRPV2 similarly suppresses differentiation when activated during the early stages, at least in part via the calcineurin-dependent pathway. In mature brown adipocytes, upregulated TRPV2 functionally contributes to β3-adrenergic induction of thermogenic gene expression and non-shivering thermogenesis. Although TRPV2 likely detects mechanical input during early differentiation in a manner analogous to Piezo1, accumulating evidence suggests that TRPV2 in differentiated adipocytes may become responsive to changes in the redox state and tissue temperature, conditions that intensify when metabolic demand and heat production increase. Together, these observations support the idea that brown adipocyte function is regulated by sequential shifts in the dominant activation cues and signaling roles of mechanically sensitive channels, such as Piezo1 and TRPV2, rather than by static channel functions across all developmental stages.
Emerging evidence further suggests that TRPV2 activity in differentiated brown adipocytes may be modulated by post-translational mechanisms, such as methionine oxidation, potentially enabling TRPV2 to integrate metabolic or oxidative signals with thermogenic responses during sympathetic activation. However, it remains unclear whether such modifications occur in vivo during non-shivering thermogenesis and whether they are required for TRPV2-dependent Ca^2+^ signaling. In contrast, Piezo1 appears to act primarily during the early lineage stages and could exert additional indirect effects on adipose tissue remodeling. The distinct stimuli detected by Piezo1 and TRPV2, together with their different periods of activity during adipocyte development, form the basis for a developmental-stage-dependent functional “modal shift” model for TRP and Piezo channel signaling in brown adipocytes (Figure 1).
In addition to TRPV2 and Piezo1 channels, several TRP channels, including TRPV1, TRPV4, and TRPM8, have been reported to be involved in adipocyte differentiation, browning, and thermogenic regulation. TRPV1 has been linked to adipocyte browning and intracellular Ca^2+^-related thermogenic signaling, whereas TRPV4 and TRPM8 differentially modulate metabolic and thermogenic responses. Other TRP channels, such as TRPV3, TRPM4, TRPA1 and TRPC5, are likewise expressed in white and/or brown adipocytes and have been implicated in the regulation of processes including adipocyte differentiation. Along with stage-dependent changes in Piezo1 and TRPV2 activity, additional TRP channels likely act cooperatively to regulate brown adipocyte function. Together, these channels likely form a broader regulatory network that supports thermogenic and metabolic responses in brown adipose tissue.
Despite these advances, several key questions remain unresolved. First, the mechanisms that regulate the developmental-stage-dependent expression and functional engagement of TRPV2 and Piezo1 remain largely unknown. The transcriptional, post-transcriptional, and/or epigenetic programs that establish these shifts in channel availability and responsiveness are yet to be defined, and elucidating these processes would substantially strengthen the mechanistic foundation of the modal shift model. Second, the mechanism by which TRPV2 positively regulates the thermogenic functions of mature brown adipocytes remains unclear. While TRPV2-mediated Ca^2+^ influx has been linked to the induction of thermogenic gene expression, it is unclear whether TRPV2 primarily acts as a permissive Ca^2+^ entry pathway or whether its activity is tuned by physiological inputs associated with thermogenesis, such as local temperature elevation or changes in the cellular redox state. In particular, whether temperature-dependent gating and methionine oxidation of TRPV2 occur in vivo during non-shivering thermogenesis and whether these processes are necessary for TRPV2-dependent Ca^2+^ signaling remain poorly understood. Moreover, whether TRPV2 activity in vivo contributes not only to thermogenic activation but also to the regulation of brown adipocyte differentiation in a stage-dependent manner remains to be determined. Finally, it is also unclear whether Piezo1-mediated Ca^2+^ signaling in vivo exerts sustained effects on adipocyte differentiation or adipose tissue remodeling, including early lineage stages.
Future studies clarifying how TRP and Piezo channels are activated under physiological and pathophysiological conditions, and how their functions shift during adipocyte development, will further advance our understanding of thermogenic regulation. Given the widespread expression and diverse physiological roles of TRP and Piezo channels, their potential as therapeutic targets for metabolic diseases requires careful consideration of tissue specificity and safety. In particular, tissue-specific (e.g., Ucp1-Cre or Adipoq-Cre mice) and developmental-stage-dependent (e.g., Myf5-Cre mice) deletions of TRP and Piezo channels in adipocytes are essential to distinguish cell-autonomous effects from systemic or developmental contributions and to test the proposed modal shift model. The relevance of the proposed modal shift framework to human brown or beige adipocytes remains to be established, as functional evidence in human systems is currently limited. Moreover, even in mouse models, the physiological significance and regulatory mechanisms of this framework are not yet fully defined. Addressing these issues is essential for improving our understanding of thermogenic regulation and its potential relevance to metabolic diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Spiegelman B.M. Flier J.S. Adipogenesis and obesity: Rounding out the big picture Cell 19968737738910.1016/S 0092-8674(00)81359-88898192 · doi ↗ · pubmed ↗
- 2Wu J. Bostrom P. Sparks L.M. Ye L. Choi J.H. Giang A.H. Khandekar M. Virtanen K.A. Nuutila P. Schaart G. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human Cell 201215036637610.1016/j.cell.2012.05.01622796012 PMC 3402601 · doi ↗ · pubmed ↗
- 3Argyropoulos G. Harper M.E. Invited Review: Uncoupling proteins and thermoregulation J. Appl. Physiol.2002922187219810.1152/japplphysiol.00994.200111960973 · doi ↗ · pubmed ↗
- 4Nedergaard J. Ricquier D. Kozak L.P. Uncoupling proteins: Current status and therapeutic prospects EMBO Rep.2005691792110.1038/sj.embor.740053216179945 PMC 1369193 · doi ↗ · pubmed ↗
- 5Feldmann H.M. Golozoubova V. Cannon B. Nedergaard J. UCP 1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality Cell Metab.2009920320910.1016/j.cmet.2008.12.01419187776 · doi ↗ · pubmed ↗
- 6Ikeda K. Maretich P. Kajimura S. The Common and Distinct Features of Brown and Beige Adipocytes Trends Endocrinol. Metab.20182919120010.1016/j.tem.2018.01.00129366777 PMC 5826798 · doi ↗ · pubmed ↗
- 7Li Y. Fromme T. Schweizer S. Schottl T. Klingenspor M. Taking control over intracellular fatty acid levels is essential for the analysis of thermogenic function in cultured primary brown and brite/beige adipocytes EMBO Rep.2014151069107610.15252/embr.20143877525135951 PMC 4253847 · doi ↗ · pubmed ↗
- 8Cypess A.M. Lehman S. Williams G. Tal I. Rodman D. Goldfine A.B. Kuo F.C. Palmer E.L. Tseng Y.H. Doria A. Identification and importance of brown adipose tissue in adult humans N. Engl. J. Med.20093601509151710.1056/NEJ Moa 081078019357406 PMC 2859951 · doi ↗ · pubmed ↗