Growth, Health and Physiological Responses of Freshwater-Reared Atlantic Salmon (Salmo salar) Fed Graded Dietary Lipid Levels
Byoungyoon Lee, Junoh Lee, Saeyeon Lim, Gwanghyeok Kim, Minjae Seong, Dahyun Jeong, Sijun Han, Byung-Hwa Min, Kang-Woong Kim, Seong-Mok Jeong, Mun Chang Park, Woo Seok Hong, Se Ryun Kwon, Youngjin Park

TL;DR
This study finds that Atlantic salmon raised in freshwater don't need high-lipid diets, which can lower costs and improve aquaculture sustainability.
Contribution
The study identifies 16% dietary lipid as optimal for freshwater Atlantic salmon, challenging the need for high-lipid diets in this rearing context.
Findings
Salmon fed 16% lipid diets showed the best growth and feed efficiency.
Lower lipid diets (14%) led to reduced antioxidant activity and higher immune-related gene expression.
Transcriptomic analysis revealed 2117 differentially expressed genes between 14% and 16% lipid diets.
Abstract
Atlantic salmon require large amounts of energy during smoltification and seawater transfer, processes that involve major physiological and metabolic adjustments. To meet these energy demands, high-lipid diets are widely used in Atlantic salmon aquaculture. However, excessive dietary lipid intake has been associated with negative effects, including metabolic stress, abnormal lipid accumulation, and impaired immune responses. In recent years, Atlantic salmon have been increasingly reared entirely in freshwater without undergoing seawater transfer, raising questions about whether high-lipid diets are still necessary under these conditions. In this study, we evaluated the optimal dietary lipid level for Atlantic salmon reared exclusively in freshwater by assessing growth performance, antioxidant capacity, metabolic responses, and liver gene expression. Salmon were fed diets containing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
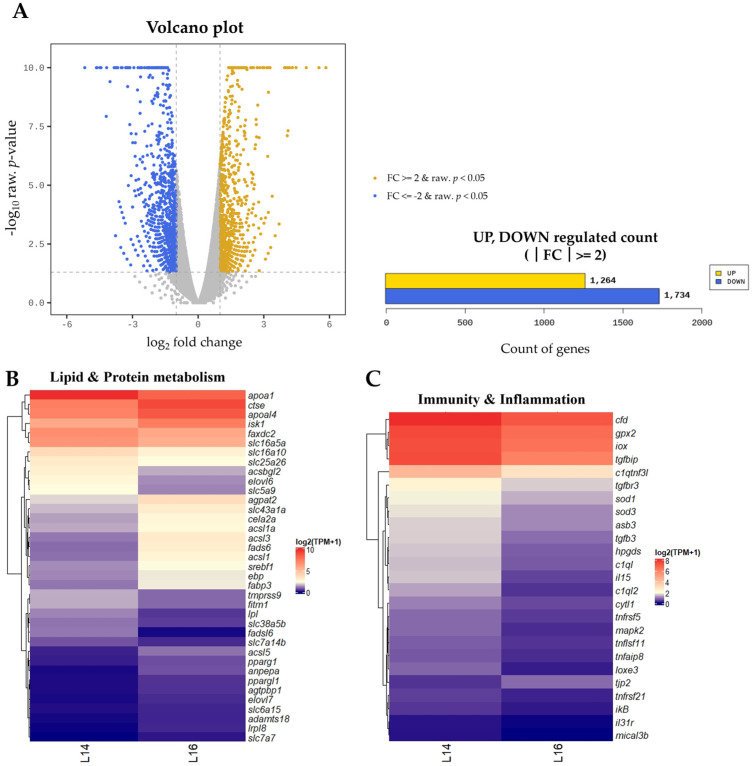 Figure 1
Figure 1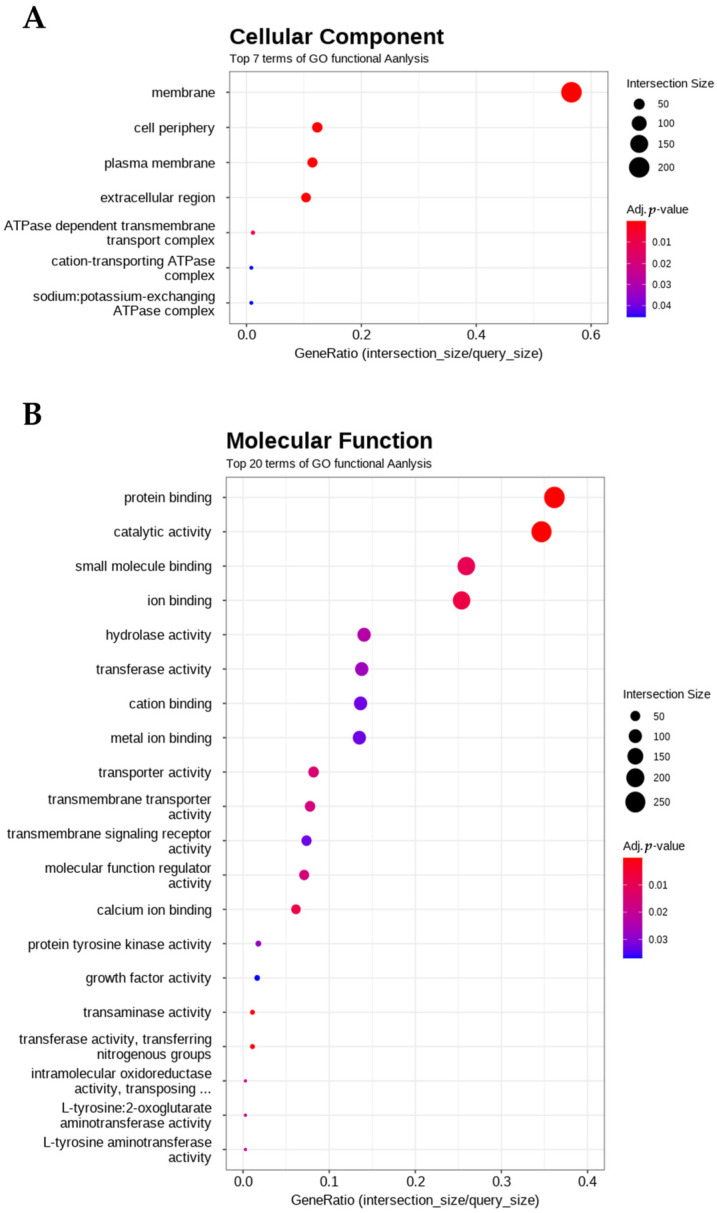 Figure 2
Figure 2- —National Institute of Fisheries Science, Republic of Korea
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture Nutrition and Growth · Aquaculture disease management and microbiota · Innovations in Aquaponics and Hydroponics Systems
1. Introduction
Aquaculture is a critical component of global food resources and continues to grow [1,2,3]. In parallel with this growth, the production and market demand for aquafeeds are also increasing. Aquafeeds serve to supply fish with essential nutrients and are directly linked to their nutritional physiology [4]. Therefore, the use of appropriate feeds is essential to ensure efficient growth and optimal health in aquaculture species [5]. Aquafeeds are formulated with dietary proteins and lipids to provide indispensable amino acids and fatty acids required for normal physiological functions [6]. In recent years, high-energy diets with elevated levels of dietary lipids have become increasingly prevalent, aiming to support faster and more efficient growth performance in cultured fish species [7,8].
Dietary lipids are essential components of formulated aquafeeds, serving as a primary energy source for fish with a higher energy density than proteins or carbohydrates. Lipids play various critical roles in supporting growth performance, body composition, and overall health in aquaculture species [9,10]. In addition, dietary lipids are the sole source of essential fatty acids, which are crucial for maintaining normal physiological functions and immune competence in fish [7,11,12]. However, excessive lipid intake has been associated with adverse effects in various fish species, including abnormal lipid deposition, impaired lipid metabolism, increased physiological stress, inflammation, and hepatic steatosis [13,14,15,16]. Therefore, determining species-specific optimal lipid requirements remains a critical objective in aquaculture nutrition research and continues to be the focus of extensive investigation.
Among aquaculture species, dietary lipid intake and requirements have been extensively studied in Atlantic salmon (Salmo salar). This is largely due to their unique migratory life cycle, which includes a smoltification phase during which juveniles undergo significant structural, physiological, and functional transformations in preparation for migration from freshwater to seawater. These changes include alterations in osmoregulatory capacity, increased oxygen consumption, and elevated metabolic rates, all of which necessitate a sufficient energy supply during this critical period [17,18]. Lipids and fatty acids play essential roles in these processes, contributing to osmoregulation, metabolic regulation, membrane structure stabilization, and transmembrane transport [19,20]. Consequently, the intake of adequate levels of essential fatty acids has been identified as a key factor in regulating smoltification in salmonids [12,21,22]. For these reasons, dietary lipids have been considered a critical nutritional component in Atlantic salmon aquaculture, and high-lipid diets are increasingly adopted to support efficient growth and energy provision. However, the rising cost of lipid sources continues to pose economic challenges to feed formulation and overall aquaculture profitability.
Atlantic salmon reared in freshwater environments exhibit a lower physiological sensitivity to dietary fatty acids, which has been interpreted as an evolutionary adaptation to natural dietary conditions [23]. In particular, freshwater-adapted Atlantic salmon and sockeye salmon have demonstrated greater capacities for fatty acid deposition in muscle tissues and more efficient lipid biosynthesis, which are associated with improved growth rates and enhanced feed conversion efficiency [24,25,26]. These traits indicate distinct advantages in terms of metabolic efficiency and cost-effectiveness under freshwater aquaculture systems. Indeed, our previous studies assessing the physiological responses of parr-stage Atlantic salmon prior to smoltification under varying dietary lipid levels reported no significant differences in growth performance, antioxidant enzyme activity, lipid metabolism, or expression of immune-related genes [27]. These findings suggest that early life stages of Atlantic salmon may not require high dietary lipid intake and that lipid levels have a limited physiological impact in the absence of seawater transfer. Consequently, low-lipid feed strategies may be viable in freshwater-based rearing systems. Recently, there has been growing interest in developing rearing technologies that enable the full life cycle of Atlantic salmon to be completed in freshwater environments [28,29]. Accordingly, the present study aims to evaluate the graded dietary lipid inclusion of Atlantic salmon reared entirely in freshwater, thereby providing foundational data to support the formulation of cost-effective and nutritionally optimized feeds for sustainable salmon aquaculture.
2. Materials and Methods
2.1. Experimental Diets
Four experimental diets were formulated to contain identical crude protein levels (45%) but varying lipid levels: 14.02% (L14), 16.51% (L16), 18.61% (L18), and 20.35% (L20). To compensate for variation in lipid inclusion across treatments, starch was used as a non-lipid substitute so that the total formulation remained constant. The formulation and proximate composition of diets are presented in Table 1. The proximate composition was determined in duplicate, and the mean values are reported. Fish oil and soybean oil were used as lipid sources and incorporated at a 1:1 ratio in all diets. In brief, feed ingredients and water were thoroughly mixed for 30 min using a vertical mixer (HYVM-1214; Hanyoung Corp., Hanam, Republic of Korea). The mixed feed was then pelleted using a pelletizer (SP-75; Geumgang ENG, Busan, Republic of Korea), followed by drying at 60 °C for 2 h in a hot air dryer (SHI-300; Shinhanil, Gwangju, Republic of Korea). The diets were stored at −20 °C until use.
2.2. Experimental Fish and Feeding Trial
Atlantic salmon used in this study were purchased as eggs from Benchmark Genetics (Hafnarfjörður, Iceland). Eggs hatched, and juvenile salmon were raised at the Gangwon-do Inland Fisheries Resource Center. All fish were raised in freshwater without undergoing transfer to seawater and were fully acclimated to the freshwater environment. The initial weight and total length of the fish used in the experiment were 241.5 ± 9.7 g and 30.6 ± 0.2 cm, respectively. A total of 200 fish were randomly distributed into four 1000-L circular fiberglass tanks (50 fish per tank) equipped with a recirculating aquaculture system and filtration unit. The fish were acclimated to experimental conditions in the same tanks for two weeks before the experiment began. A 12-week feeding trial was then conducted.
Experimental diets were provided twice daily to apparent satiation (09:00 and 16:00). During the feeding trial, pellet stability was visually monitored under freshwater conditions, and pellets remained intact until consumption, with no apparent disintegration observed. Water quality was monitored daily using a commercial water analysis kit (HUMAS, Daejeon, Republic of Korea). Throughout the trial, the water temperature was maintained at approximately 15 °C using a cooling and heating tank system, with pH 6.5–7.0, dissolved oxygen (DO) levels of 9–11 mg/L, and ammonia concentrations below 1 mg/L across all tanks.
2.3. Sampling
At the end of the 12-week feeding trial, sixteen fish per tank were randomly selected and anesthetized in ethyl 3-aminobenzoate methanesulfonate solution (100 mg/L; Sigma-Aldrich, St. Louis, MO, USA) for biometric measurements (total length and body weight). These measurements were used to evaluate growth performance, feed utilization, and condition indices
Among these, three individuals were immediately frozen at −20 °C for subsequent whole-body proximate composition analysis. Blood samples were collected from the caudal vasculature of the remaining four fish using sterile syringes. Blood was collected into 1.5 mL tubes coated with 20 μL of heparin (BD Microtainer, Franklin Lakes, NJ, USA) for each sample. Plasma was separated by centrifugation at 3000 rpm for 15 min at 4 °C and stored at −80 °C for further biochemical analyses. Biochemical parameters included antioxidant enzymes such as superoxide dismutase (SOD) and glutathione peroxidase (GPx), innate immune parameter lysozyme (LZM), and stress and health indicators such as glutamic-pyruvic transaminase (GPT), glutamic-oxaloacetic transaminase (GOT), glucose, and total protein (TP) concentrations.
Following blood collection, the viscera and liver were excised and weighed to determine viscerosomatic and hepatosomatic indices. A portion of the liver tissue was collected in 1.5 mL microcentrifuge tubes and immediately stored at −80 °C for subsequent molecular analyses, including RNA sequencing.
2.4. Proximate and Fatty Acid Composition of the Experimental Diets and Whole-Body
Experimental diets and whole-body samples from five randomly selected fish per tank were homogenized prior to analysis. Proximate composition was determined according to standard methods described by the AOAC. Moisture content was analyzed via oven-drying at atmospheric pressure (135 °C for 2 h). Crude protein was determined using the Kjeldahl method (nitrogen × 6.25) [30], and crude ash was determined using a muffle furnace. Moisture, ash and lipid contents were determined according to the methods described previously [31]. Crude lipid content was analyzed via Soxhlet extraction using samples that had been freeze-dried for 12 h (Soxtec system 1046; Tecator AB, Hilleroed, Sweden).
For the fatty acid analysis, total lipids from experimental diets and fish samples were extracted by following a previous method [32]. The extracted lipids were methylated using 14% boron trifluoride-methanol (Sigma-Aldrich, USA) to obtain fatty acid methyl esters. Fatty acid profiles were determined using gas chromatography (GC, Clarus 600; PerkinElmer, Waltham, MA, USA) equipped with a capillary column (SP™-2560, 100 m × 0.25 mm i.d., film thickness 0.20 μm; Supelco, Bellefonte, PA, USA). Helium was used as the carrier gas. The oven temperature was initially set at 140 °C and increased to 240 °C at a rate of 4 °C/min. Identification and quantification of fatty acids were performed by comparison with a known standard mixture (PUFA 37 Component FAME Mix; Supelco, Bellefonte, PA, USA). The detailed fatty acid composition of each experimental diet is presented in Table 2.
2.5. Hematobiochemical Analysis
Plasma samples from each treatment group were thawed completely on ice prior to analysis. Each sample was diluted 1:200 in the assay buffer prior to measurement. Antioxidant and immune-related parameters, including SOD, GPx, and LZM, were quantified using commercially available FISH-specific ELISA kits (CUSABIO, Houston, TX, USA), following the manufacturer’s instructions.
Clinical biochemical parameters, including GPT, GOT, glucose, and TP, were analyzed using commercial diagnostic kits (Asan Pharm, Seoul, Republic of Korea) according to the manufacturer’s protocols. Absorbance was measured using a microplate spectrophotometer (TECAN Infinite 200 Pro M, Männedorf, Switzerland) at wavelengths of 450 nm, 505 nm, 540 nm, and 570 nm, depending on the assay. Standard curves and sample concentrations were calculated using CurveExpert software (version 1.3).
2.6. RNA Sequencing and Transcriptomic Analysis
Liver tissue samples collected from the L14 and L16 dietary groups were used for transcriptomic analysis. For RNA-seq analysis, RNA was extracted from three fish per dietary treatment. Equal amounts of RNA from each individual were then pooled within each group prior to library preparation and sequencing. Total RNA was extracted using the QIAzol Lysis Reagent and RNeasy Mini Kit (Qiagen, Hilden, Germany), followed by DNase treatment to remove genomic DNA contamination. RNA libraries were prepared using the TruSeq Stranded Total RNA kit with Ribo-Zero (Illumina, San Diego, CA, USA). Purified RNA was randomly fragmented, and the resulting RNA fragments were reverse-transcribed into cDNA. The cDNA was then ligated with adapters, amplified by PCR, and size-selected to include insert sizes ranging from 200 to 400 bp.
Sequencing was performed using the NovaSeq 6000 platform (Illumina, San Diego, CA, USA) with paired-end reads (101 bp). Raw sequencing reads underwent quality control to remove low-quality reads, adapter sequences, contaminant DNA, and PCR duplicates. Clean reads were aligned to the reference genome (Ssal_v3.1) using HISAT2, and transcript assembly was conducted with StringTie (version 2.2.3). Transcript abundance was quantified using transcript per million of mapped reads (TPM). RNA-sequencing (RNA-seq) data processing and normalization were performed following methods described previously [33,34].
Differentially expressed genes (DEGs, an absolute fold change ≥ 2 and Benjamini-Hochberg adjusted p < 0.05) were identified based on statistical comparisons between dietary groups using hypothesis testing. Annotated DEGs were subjected to functional enrichment analysis using the Gene Ontology (GO) databases. GO enrichment analysis was conducted using gProfiler, covering the three GO domains: molecular function and cellular component.
2.7. Statistical Analysis
Fish growth performance parameters were expressed as the mean ± standard deviation (mean ± SD). Statistical analyses and data visualization were conducted using RStudio (version 2025.05.1+513), BioRender (BioRender.com) and CurveExpert (version 1.3). Normality and homogeneity of variance were assessed using the Shapiro–Wilk test and Bartlett’s test, respectively. One-way analysis of variance was then performed, and significant differences among groups were compared using Tukey’s honestly significant difference (p < 0.05). For non-parametric comparisons, the Kruskal–Wallis test was applied, followed by Dunn’s post hoc test to identify significant differences among experimental groups.
3. Result
3.1. Growth Performance and Feed Efficiency Ratio
Throughout the 12-week feeding trial, no pathological symptoms were observed in any experimental groups. Fish fed the L16 diet, containing 16.5% dietary lipid, exhibited the highest weight gain rate (WGR; 84.0 ± 9.1%) and feed efficiency ratio (FER; 102.7 ± 11.1%), representing the best growth performance among treatments. In contrast, fish fed the L14 diet (14% lipid) showed significantly lower WGR (44.9 ± 12.4%) and FER (72.3 ± 19.9%) than those fed the L16 diet (p = 0.049). Diets containing 18% and 20% lipids yielded comparable WGR values, with no significant differences between them (p = 0.991). Morphometric indices, including viscerosomatic index (VSI), hepatosomatic index (HSI), and condition factor (CF), did not differ significantly among dietary treatments (VSI: p = 0.965, HSI: p = 0.526, CF: p = 0.584) (Table 3).
3.2. Whole-Body Composition
3.2.1. Proximate Composition
The proportion of whole-body protein decreased with increasing dietary lipid levels, with the highest value observed in fish fed the L14 diet (p < 0.001). Conversely, increasing dietary lipids resulted in higher whole-body lipid content, with the L20 group showing a significantly greater proportion than the other groups and L14 the lowest (p < 0.001). Whole-body ash content was highest in the L14 group and lowest in the L18 group (p = 0.004). Moisture content did not differ significantly among dietary treatments (p = 0.224) (Table 4).
3.2.2. Fatty Acids Composition
Fish fed the L18 and L20 diets, which contained relatively higher dietary lipid levels, exhibited lower proportions of EPA and DHA in whole-body. In contrast, the lowest proportion of C15:0 was observed in fish fed the L14 diet, which contained the lowest dietary lipid level. The proportions of other fatty acids were comparable across all groups (Table 5).
3.3. Plasma Antioxidant Capacity and Biochemical Analysis
No significant differences in SOD activity were observed among dietary treatments (p = 0.553). However, GPx activity was highest in fish fed a diet containing 18% lipid and lowest in those fed the L14 diet (p = 0.041). LZM activity did not differ significantly among treatments (p = 0.797). Biochemical analyses revealed no significant differences in GPT, GOT, or TP values across dietary groups (GPT: p = 0.288, GOT: p = 0.798, TP: p = 0.211) (Table 6). Notably, plasma glucose concentration differed among dietary treatments (p = 0.017), with a significantly lower value in the L16 group compared with the L18 group (p = 0.020). No significant differences in glucose levels were detected among treatments other than L16.
3.4. Liver Transcriptomic Profiling Analysis
3.4.1. Differentially Expressed Genes Analysis
Based on the growth results, we selected the L14 and L16 treatments, which showed a significant difference in growth for RNA-seq. Comparative transcriptomic analysis between the L14 and L16 dietary groups identified a total of 2117 DEGs, comprising 905 upregulated and 1212 downregulated genes in the L14 group relative to the L16 group (Figure 1A and Supplementary Table S1). The overall expression patterns of DEGs are visualized using a heatmap. Genes (apoal4, acsl1, acsl1a, acsl3, acsl5, ebp and lrpl8) involved in fatty acid metabolism, lipogenesis and activation were significantly upregulated in L16 group. However, several proteases and amino acid transporter-related genes (slc16a10, slc7a7, slc7a14b, slc25a26 and slc38a5b) displayed higher expression in the L14 group (Figure 1B). The expression levels of immune and inflammation-related genes were higher in the L14 dietary group than in the L16 group. Representative transcripts that were more highly expressed in the L14 group included antioxidant enzymes (sod1, sod3 and gpx2), cytokine-related genes (iox, tnfrsf21, tnflsf11, tnfaip8, tnfrsf5, il15 and il31r), regulators of cellular immunity and inflammation (tgfb3, tgfbr3, tgfbip and asb3) and complement-associated genes (cfd, c1ql, c1ql2 and c1qtnf3l) (Figure 1C).
3.4.2. GO Functional Analysis
In the GO category of cellular components, the most significantly enriched terms included membrane, cell periphery, plasma membrane, extracellular region, ATPase-dependent transmembrane transport, cation-transporting ATPase, and sodium–potassium-exchanging ATPase. The DEGs associated with these terms indicated that the membrane was the most prominent GO term differentiating the L14 and L16 dietary groups (Figure 2A).
In the molecular function category, the top enriched terms were protein binding, catalytic activity, small molecule binding, and ion binding, along with specific terms such as transferase activity, transporter activity, transmembrane transporter activity and transmembrane signaling (Figure 2B).
4. Discussion
Atlantic salmon is one of the most economically important species in global aquaculture, and extensive nutritional research has focused on ensuring stable and sustainable production [35]. With the expansion of aquaculture, high-lipid diets have increasingly been used to promote rapid growth; however, excessive lipid intake has been associated with adverse physiological effects [36,37]. Atlantic salmon undergo smoltification and seawater migration, during which adequate essential fatty acids are critical for maintaining metabolic and osmoregulatory balance [38]. In contrast, juveniles reared in freshwater exhibit greater lipid biosynthetic capacity and a reduced dependence on dietary fatty acids [23]. Previous observations suggest that high-lipid diets are not essential for freshwater-reared Atlantic salmon, and that low-lipid feeding strategies may improve FER while supporting more efficient feed utilization [27]. Therefore, this study evaluated the effects of graded dietary lipid levels on growth performance, physiological responses, and transcriptomic profiles in Atlantic salmon reared entirely in freshwater. These findings provide fundamental insights for optimizing dietary lipid requirements in freshwater-based salmon aquaculture.
4.1. Inadequate Dietary Lipid Levels Impair the Growth Performance of Atlantic Salmon
WGR and SGR are key performance indices in aquaculture [39]. In the present study, fish fed the diet containing 16% lipid exhibited the highest WGR and SGR, whereas those fed the 14% lipid diet showed the lowest growth performance. In the study on rainbow trout [40], fish fed a low-lipid diet exhibited reduced WGR, which was attributed to insufficient supply of essential fatty acids. Similarly, it has been reported that in Atlantic salmon at the parr stage, growth rate did not increase proportionally with higher dietary lipid levels, and no significant differences were observed between diets containing 16% and 28% lipid [41]. This outcome was explained by delayed digestion and reduced absorption efficiency associated with excessive lipid intake. Although excessive lipid intake is frequently associated with somatic lipid deposition and increased VSI and HSI in fish [42,43,44,45,46], we detected no significant differences, indicating that dietary lipid levels of up to 20% did not promote visceral fat accumulation under the present experimental conditions. Collectively, these results indicate that dietary lipid levels of 16% or higher are adequate for freshwater-reared Atlantic salmon, whereas lower levels (≤14%) are inadequate and compromise growth performance.
4.2. An Appropriate Level of Dietary Lipid Intake Is Required to Maintain Efficient Body Lipid Composition
Whole-body composition is closely coupled to growth in fish [47]. Consistent with this, numerous studies have documented a significant positive correlation between dietary lipid level and whole-body lipid content [18,48,49,50], a pattern also observed in the present work. As whole-body lipid increased, the proportional contribution of protein declined. Similar inverse shifts between body lipid and protein have been reported in Atlantic salmon and marble goby (Oxyeleotris marmorata) with increasing lipid intake [48,51]. The optimal body composition for growth differs among fish species. Previous studies have reported that lipid levels are not necessarily proportional to growth performance [52,53]. The present results demonstrate that, under the current experimental conditions, a body composition containing approximately 57% protein and 35% lipid is most favorable for growth performance.
With respect to fatty acid profiles, fish fed comparatively lower dietary lipids exhibited higher whole-body EPA+DHA proportions. Analogous responses have been described in gilthead seabream (Sparus aurata), where the lowest dietary EPA+DHA supply yielded the highest DHA retention [54]. In European seabass (Dicentrarchus labrax), DHA deficiency elicited selective conservation and endogenous synthesis to compensate for limited supply [55], and other studies likewise report maintenance of tissue EPA+DHA despite reduced intake [56]. Notably, increased dietary fatty acid intake does not necessarily lead to proportional increases in whole-body EPA and DHA levels, as metabolomic evidence indicates that inadequate EPA and DHA may disrupt hepatic metabolic regulation and promote hepatic lipid accumulation [57]. In Atlantic salmon, limited essential fatty acid intake has been shown to trigger substantial endogenous production to offset dietary shortfalls [58,59]. Collectively, these observations suggest that the higher EPA+DHA proportions observed in fish fed lower dietary lipid levels may reflect selective retention and metabolic regulation of essential fatty acids rather than a direct linear response to dietary supply.
4.3. Appropriate Dietary Lipid Intake Is Essential for Maintaining the Health of Atlantic Salmon
SOD and GPx constitute first-line enzymatic defenses against reactive oxygen species (ROS), limiting lipid peroxidation and maintaining redox homeostasis in fish [60,61]. High-lipid feeding has been reported to disrupt lipid metabolism, exacerbate oxidative stress, and depress antioxidant enzyme activities in zebrafish (Danio rerio) [36] and carp (Cyprinus carpio) [62]. In contrast to these patterns, we detected no statistically significant differences in SOD activity among dietary treatments in the present study, whereas GPx activity was lowest at 14% dietary lipid and highest at 18%. The direction of this response is consistent with the elevated lipid requirements typical of cold-water salmonids, including Atlantic salmon and Amur grayling (Thymallus arcticus grubei) [13,63]. Although high EPA+DHA intake has been proposed to foster an oxidative milieu [64], variation in experimental diets did not alter the antioxidant status of Atlantic salmon under our conditions, suggesting that relatively high EPA+DHA did not precipitate measurable oxidative stress and that this species maintains robust redox control. Plasma glucose is a recognized proxy of metabolic stress and health in fish [65,66], and typically rises with dietary stress, inflammation, or metabolic dysregulation [67,68]. Notably, the lowest plasma glucose concentration occurred at 16% dietary lipid, indicating attenuated stress at this level. Taken together, these findings indicate that a dietary lipid level around 16% supports antioxidant capacity and physiological stability in freshwater-reared Atlantic salmon.
4.4. Low Dietary Lipid Intake Influences Lipid Metabolism and Affects the Structural Integrity of Cell Membranes
The liver is a principal organ for lipid metabolism and immune regulation [69,70]. To resolve treatment-specific hepatic responses, we performed RNA-seq on livers from fish fed the 14% and 16% lipid diets, which differed in growth performance. APOA4 (apolipoprotein A-IV), predominantly synthesized in the liver and functionally engaged with chylomicrons and high-density lipoproteins, facilitates intestinal lipid absorption and contributes to high-density lipoprotein remodeling [71]. Concordant with prior observations in fish, higher dietary lipids have been associated with apoa4 upregulation, and a similar pattern was observed in the present study. ACSLs (acyl-CoA synthetase long-chain) activate fatty acids to acyl-CoA, thereby gating their entry into β-oxidation and lipid biosynthetic pathways and accelerating cellular fatty acid handling [72]. In our dataset, acsl1, acsl1a, acsl3 and acsl5 were upregulated in L16 relative to L14, consistent with augmented hepatic lipid uptake, activation, and downstream metabolism at 16% dietary lipid. Cholesterol regulates membrane fluidity and is essential for maintaining membrane structure and intracellular signal transduction [73,74,75]. In the present study, ebp and lrp8, which were upregulated in the L16 group, were identified as key regulators involved in cholesterol biosynthesis and lipid metabolism, respectively [74,76]. Consistent with this, a study demonstrated a positive correlation between LRP8 expression and the expression of lipid metabolism–related genes [77]. These genes are all associated with the cell membrane, and the enriched GO terms identified in this study included ‘membrane’, ‘cell periphery’ and ‘plasma membrane’ under the “cellular component” category. Together, these results indicate that dietary lipid levels are associated with differences in hepatic lipid metabolism–related gene expression, with the 16% lipid diet showing a transcriptomic profile more consistent with enhanced fatty acid synthesis and metabolism compared with 14%.
Members of the solute carrier family mediate amino acid and nutrient transport and are central to growth and metabolic homeostasis; their expression is broadly tuned by nutrient-sensing pathways [78,79]. Notably, amino-acid transporter genes (slc16a10, slc7a7, slc7a14b, slc25a26, slc38a5b) were comparatively higher in L14. Other studies interpret increased amino-acid transport capacity as a compensatory response to nutrient insufficiency [80,81]. The enriched GO terms under the “molecular function” category included ‘transferase activity’, ‘transporter activity’ and ‘transmembrane signaling’. Alterations in genes associated with transporter proteins may reflect impaired nutrient transport capacity. These results appear to be related to dietary nutrient composition, suggesting that the 14% lipid diet is associated with a transcriptional profile indicative of altered nutrient transport and metabolic regulation.
4.5. A Dietary Lipid Level Below the Optimal Range May Induce Nutritional Stress in Freshwater-Reared Atlantic Salmon
The complement system is tightly linked to inflammation and innate immunity and is frequently used as a marker of inflammatory activation [82,83]. Concordantly, several complement-associated genes (c1ql2, c1ql, c1qtnf3l and cfd) were upregulated in the L14 group, reflecting higher expression of inflammation-related genes compared with L16. Among cytokine pathways, IL-15, a member of the IL-2 family, induces IFN-γ and primarily signals through downstream cascades to modulate immune cell function [84]. In gilthead sea bream (Sparus aurata), increased il15 expression has been attributed to dietary immunostimulants and associated cytokine responses [85], and in orange spotted grouper (Epinephelus coioides), expression levels of il15 rises significantly following LPS or poly I:C exposure [86]. IL-31R, which binds IL-31, is typically elevated in mucosal and chronic inflammatory settings [87,88]. TNF, a potent pro-inflammatory cytokine, is well known to cause tissue injury when excessive [89] and can be induced not only by pathogens but also by thermal, salinity, and nutritional stress [90,91,92]. TGF-β is a pleiotropic cytokine broadly involved in immune regulation, and increased expression has been reported with soybean-meal enteritis, oxidative stress, and immune stimulation (LPS, poly I:C) [93,94]. Although TGF-β is commonly stored in a latent form, ROS can activate it by oxidatively modifying the latency-associated peptide, thereby altering its conformation and releasing active TGF-β [94]. Overall, the DEG analysis between the L14 and L16 groups showed higher expression of immune and inflammation-related genes in L14. This transcriptional pattern is consistent with the 14% lipid diet being associated with increased expression of genes related to immune and inflammatory pathways, which may reflect a state of nutritional stress.
5. Conclusions
This study evaluated the effects of dietary lipid levels in freshwater-reared Atlantic salmon. Across a 12-week feeding trial, diets containing ≥16.51% lipid produced superior growth performance, and accompanying whole-body composition patterns were consistent with these outcomes. Hepatic transcriptomic analyses further indicated that a 14% lipid regimen was associated with less favorable expression of genes involved in fatty acid metabolism and immune function, together with differences across membrane-associated processes, ion-related functions, and growth factor activity. Taken together, the results suggest that under the present freshwater rearing conditions, a dietary lipid level of approximately 16% appears to be a practical and suitable target within the range tested for guiding lipid inclusion. These findings provide a foundation for the future development of feeds for freshwater-reared Atlantic salmon. However, because this study was conducted at a single life stage over a 12-week period, lipid requirements may differ during subsequent growth phases or during smoltification; therefore, longer-term studies spanning later growth stages are warranted to confirm the durability of the observed effects. In addition, in future studies, we will use the key genes identified in this RNA-seq analysis as targets for qPCR to further refine and validate the molecular responses to dietary lipid levels.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wilfart A. Garcia-Launay F. Terrier F. SoudéE. Aguirre P. Skiba-Cassy S. A step towards sustainable aquaculture: Multiobjective feed formulation reduces environmental impacts at feed and farm levels for rainbow trout Aquaculture 202356273882610.1016/j.aquaculture.2022.738826 · doi ↗
- 2Torres-Maravilla E. Parra M. Maisey K. Vargas R.A. Cabezas-Cruz A. Gonzalez A. Tello M. Bermúdez-Humarán L.G. Importance of probiotics in fish aquaculture: Towards the identification and design of novel probiotics Microorganisms 20241262610.3390/microorganisms 1203062638543677 PMC 10975645 · doi ↗ · pubmed ↗
- 3Nederlof M.A. Verdegem M.C. Smaal A.C. Jansen H.M. Nutrient retention efficiencies in integrated multi-trophic aquaculture Rev. Aquac.2022141194121210.1111/raq.12645 · doi ↗
- 4Kong W. Huang S. Yang Z. Shi F. Feng Y. Khatoon Z. Fish Feed Quality Is a Key Factor in Impacting Aquaculture Water Environment: Evidence from Incubator Experiments Sci. Rep.20201018710.1038/s 41598-019-57063-w 31932648 PMC 6957510 · doi ↗ · pubmed ↗
- 5Davis D.A. Hardy R.W. Feeding and fish husbandry Fish Nutrition Elsevier Amsterdam, The Netherlands 2022857882
- 6Hardy R.W. Kaushik S.J. Mai K. Bai S.C. Fish nutrition—History and perspectives Fish Nutrition Elsevier Amsterdam, The Netherlands 2022116
- 7Zhang G. Guan J. Chen F. Xu J. Su N. Zhang H. Wang Z. Wang S. Xu C. Xie D. Effects of dietary lipid source and level on growth performance, antioxidant capacity, and hepatic metabolism in marine teleost Trachinotus ovatus: Insights from fatty acid composition Aquac. Rep.20254310291110.1016/j.aqrep.2025.102911 · doi ↗
- 8Fang H.-H. Zhao W. Xie J.-J. Yin P. Zhuang Z.-X. Liu Y.-J. Tian L.-X. Niu J. Effects of dietary lipid levels on growth performance, hepatic health, lipid metabolism and intestinal microbiota on Trachinotus ovatus Aquac. Nutr.2021271554156810.1111/anu.13296 · doi ↗