Activation of OsOxO2 by T-DNA Insertion Affects Plant Height and Leaf Angle in Rice
Xing-wen Hu, Hai-yan Wang, Xiao-lu Yu, David W. M. Leung, Yu-long Chen, Dan-hong Chen, Ke Chen, Xin-xiang Peng, E-e Liu

TL;DR
A rice mutant with altered leaf angle and dwarfism is caused by T-DNA insertion near OsOxO2, affecting plant architecture through OsOxO2 and OsIAAGLU.
Contribution
The study identifies OsOxO2 and OsIAAGLU as key regulators of rice leaf angle and plant height through T-DNA insertion.
Findings
T-DNA insertion upstream of OsOxO2 causes increased transcript levels of OsIAAGLU and related genes.
Mutation of OsIAAGLU partially restores wild-type leaf angle and plant height.
OsOxO2 regulates OsIAAGLU, OsOxO1, and OsOxO3 in a chromosome position-dependent manner.
Abstract
Inclination of leaf and plant height are important agronomic traits that are closely related to grain yields. Proteins involved in regulating free auxin (IAA) levels have a central role in modulating rice leaf angle and plant height. In this study, a rice mutant (named lc4) was found to have enlarged leaf angles and exhibit dwarfism compared to wild-type (WT) plants. Flanking analysis revealed that the mutation was caused by a T-DNA insertion at 270 bp upstream of the oxalate oxidase 2 (OsOxO2) ATG start codon. The insertion significantly increased the transcript levels of indole-3-acetic acid glucosyltransferase (OsIAAGLU), OsOxO1, OsOxO2 and OsOxO3 and OxO activity in lc4. Mutation of OsIAAGLU in lc4 led to partial restoration of the WT leaf angle and plant height phenotypes in iaaglu/lc4 which, however, still exhibited OxO activity comparable with that in lc4. On the other hand,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
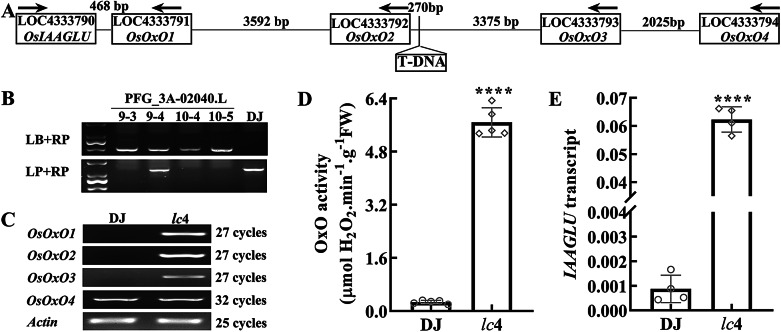 Figure 1
Figure 1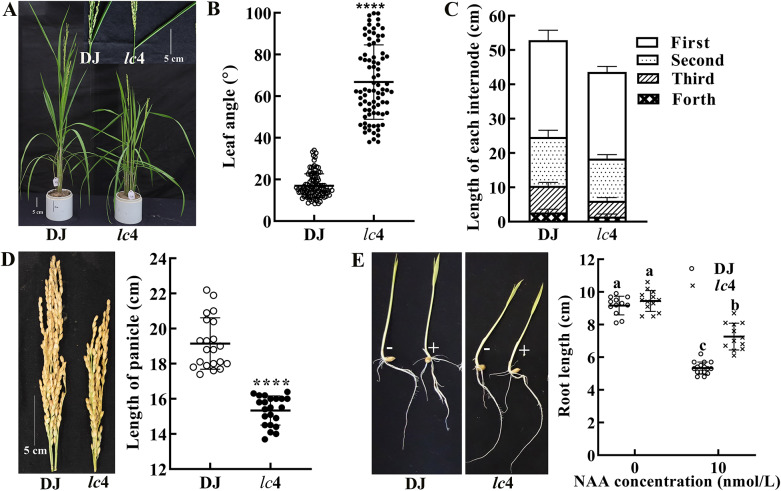 Figure 2
Figure 2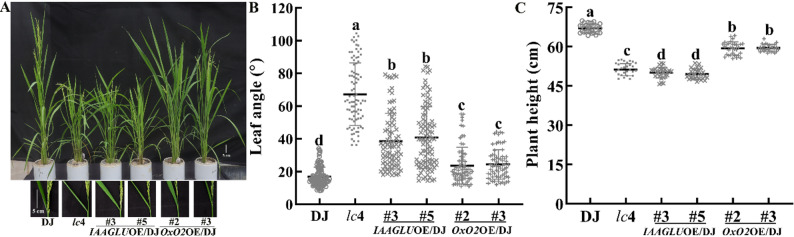 Figure 3
Figure 3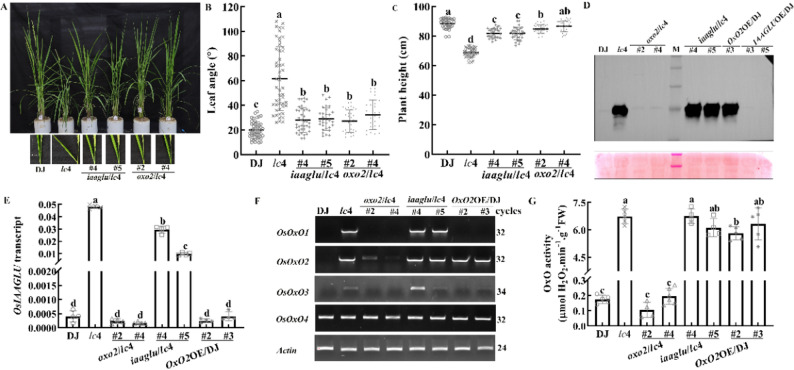 Figure 4
Figure 4- —National Natural Science Foundation of China
- —http://dx.doi.org/10.13039/501100003453Natural Science Foundation of Guangdong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant responses to water stress · Plant pathogens and resistance mechanisms · Plant Stress Responses and Tolerance
Introduction
Plant architectural attributes, including tiller number, leaf/tiller angle and plant height, are important agronomic traits in rice and play an important role in the relationships among photosynthetic efficiency, planting density and grain yield (Burgess et al. 2017; Huang et al. 2021b; Li et al. 2025; Weng et al. 2025). Many mutants affecting plant architecture have been identified genetically, genes underlying the mutant phenotypes are mainly related to auxin and/or brassinosteroid (BR) (Zhang et al. 2009, 2015; Du et al. 2012; Zhao et al. 2013; Yamamuro et al. 2000; Sakamoto et al. 2006). Auxin plays vital roles in maintaining normal development of rice blade because it is involved in the elongation of parenchyma cells in the lamina joint connecting the leaf and the sheath and regulation of leaf angle (Zhao et al. 2013; Zhang et al. 2015). Therefore, reduction in free auxin content in lamina joint tissues resulted in increased flag leaf angle (Zhang et al. 2009, 2015; Du et al. 2012; Zhao et al. 2013). A number of mutants with altered IAA levels and responses to IAA also showed modified leaf angle. IAA early responsive genes, including Aux/IAA, IAA receptor TIR1, IAA response factors ARF and GH3, have been reported involved in regulating leaf angle in rice. Transgenic plants overexpressing OsIAA1 (a member of rice Aux/IAA family) displayed dwarfism and enlarged leaf angle phenotypes (Song et al. 2009). OsTIR1- or OsAFB2-RNAi transgenic lines also showed enlarged leaf inclination (Bian et al. 2012). Rice gain-of-functions mutant tld1-D (OsGH3-13) showed enlarged leaf angle, dwarfism and tiller number increase phenotypes (Zhang et al. 2009). Similarly, typical IAA-deficient phenotypes including dwarfism and enlarged leaf angle were also observed in plants overexpressing OsGH3-1 (Zhao et al. 2013) or OsGH3-2 (Du et al. 2012). Additionally, overexpression of ZmIAAGLU in Arabidopsis made the plants insensitive to IAA, and to display shorter roots and curly leaf blade (Ludwig-Müller et al. 2005). This gene encodes an enzyme that catalyzes IAA conjugation with uridine diphosphate glucose (UDPG) to form IAA-glucose (Szerszen et al. 1994). OsIAAGLU which shares 67% identity with ZmIAAGLU can also catalyze the reaction of free IAA with glucose to generate IAA-glucose. Transgenic rice plants overexpressing OsIAAGLU exhibited dwarfism, increased leaf angle and tiller number, and shorter spike (Choi et al. 2012; Yu et al. 2019). These findings demonstrate that endogenous free IAA levels play a crucial role in regulating rice leaf angle and plant height. Despite the importance of IAA in leaf angle regulation has been well established, the precise mechanisms underlying its fine-tuning remain unclear.
Besides auxin, BR, ethylene, GA and other hormones could also participate synergistically in regulation of rice plant architecture (Shimada et al. 2006; Sakamoto et al. 2006; Hong et al. 2002, 2003; Yamamuro et al. 2000). Additionally, ILA1 encodes a Ser/Thr kinase which can regulate leaf angle through altering strength of mechanical tissue and cell wall composition in lamina joint. Leaf angle of Ila1-deficent mutant increased remarkably, but the sensitivity to BR was similar to WT (Ning et al. 2011). Mutation of LC2 encoding a vernalization insensitive 3-like protein resulted in enlarged leaf angle due to increased cell division in the adaxial region of the lamina joint (Zhao et al. 2010). LPA1, a typical Cys-2/His-2 zinc finger protein, can also regulate leaf angle by modulating the cell elongation in the adaxial epidermis of lamina joint (Wu et al. 2013). Therefore, the regulation of leaf angle is very complex involving a multitude of factors, the molecular mechanism remains to be elucidated.
Although mutants associated with plant architecture are numerous, most genes underlying the mutant phenotype have been found to act through altering cell division or composition of cell wall (Ueguchi-Tanaka et al. 2000; Shimada et al. 2006; Tong et al. 2014; Liu et al. 2024). Oxalate oxidase (OxO), as a crucial members of the germin-like proteins which perform a variety of biological functions in plant development, stress response, and pathogen resistance in crops (Manosalva et al. 2009), catalyzes conversion of oxalate into H_2_O_2_ and CO_2_, simultaneously accompanying with the release of Ca^2+^ (Lane et al. 1993). It has been considered to be involved in the modification of cell wall architecture, because OxO is localized in cell wall and within the cell type which characteristically restricted in the extent of cellular enlargement, such as coleorhiza, coleoptile, scutellum and vascular bundles (Lane et al. 1993; Caliskan and Cuming 1998), but their definite physiological functions are still unknown.
In rice genome, four tandemly duplicated OxO-encoding genes (OsOxO1, OsOxO2, OsOxO3, and OsOxO4) are clustered on chromosome 3, exhibiting over 90% sequence similarity at both nucleotide and amino acid levels. These four genes have different temporal and spatial patterns of expression (Carrillo et al. 2009). Here, we identified a rice oxalate oxidase 2 (OsOxO2) T-DNA mutant, which exhibited enlarged leaf angles and dwarfism compared to the wild-type plants (WT). T-DNA insertion elevated the transcript levels of OsOxO1, OsOxO2, OsOxO3 and OsIAAGLU. Therefore, this T-DNA mutant was of particular interest to study regulation of phenotypes via in situ gene activation. Studies here on the mechanisms of how the T-DNA insertion results in the altered plant architecture support the hypothesis that OsOxO2 is involved in IAA signaling regulation of plant architecture by modulating the in situ expression level of OsIAAGLU in rice.
Materials and Methods
Plant Materials and Growth Conditions
Rice (Oryza sativa L) cv. Dongjin (DJ) was used to generate OsOxO2/OsIAAGLU overexpression transgenic plants. The lc4 mutant of rice was identified from the RiceGE database (http://signal.salk.edu/cgi-bin/RiceGE). After germination, seedlings were grown in Kimura B complete nutrient solution (Yoshida et al. 1976) under experimental conditions as described in the respective figure legend. For maturation-stage phenotyping, plants were grown in a paddy field in Guangzhou.
Identification of the lc4 Mutant
The insertion site of T-DNA was determined by PCR amplification using T-DNA-specific primer LB (matching the left border of the inserted T-DNA pGA2715) and gene-specific primer RP (matching the flanking sequence of the expected insertion site in the downstream of OsOxO2 gene), followed by sequencing to identify the insertion site. Homozygous T-DNA insertion mutants (with both alleles disrupted) were identified by PCR-based methods with gene-specific primer pairs LP (matching the flanking sequence of the expected insertion site in the upstream of OsOxO2 gene) and RP together with RP and LB (Jeong et al. 2002). The single T-DNA insertion event in lc4 was verified by analyzing the segregation ratio in progeny from heterozygous mutants (one allele disrupted).
Expression Analysis of OsOxO1-4 and OsIAAGLU
Approximately 0.1 g fresh rice leaves were collected, then frozen in liquid nitrogen immediately and ground into a fine powder. Subsequently, total RNA was extracted with RNAiso Plus (Takara) and reverse-transcribed following the manufacturer’s instructions (Vazyme Biotech, China). The transcripts of OsOxO1-4 were analyzed by semi-quantitative RT-PCR, while the expression of OsIAAGLU was quantified by qRT-PCR on PTC200 (BIO-RAD) using SYBR green master mix (Bimake, China). Actin was used as an internal control. The level of OsIAAGLU transcript was calculated using the following formula: Target gene transcript = 2^−(Ct Target gene−Ct Actin)^. All primer sequences are listed in Table S1.
Construction of Rice Transgenic Plants
For the construction of OsIAAGLUOE and OsOxO2OE overexpression vector, the full-length OsIAAGLU/OsOxO2 coding sequence was amplified using RT-PCR and cloned into the poX vector harboring the Ubi promoter (provided by Professor Yao-Guang Liu, College of Life Sciences, South China Agricultural University, China). The vectors were subsequently introduced into Agrobacterium tumefaciens strain EHA105 for transformation of the calli of rice variety Dongjin (DJ). iaaglu/lc4 or oxo2/lc4 mutants were constructed using the CRISPR/Cas9 system following the protocol described by Ma et al. (2015). First, one gRNA specific to OsOxO2/OsIAAGLU was inserted into the vector pYLCRISPR/Cas9Pubi-B (glyphosate resistance in rice), then the vector was introduced into Agrobacterium tumefaciens strain EHA105 for transformation of lc4 calli using the method of Hiei et al. (1994). The mutation of OsOxO2/OsIAAGLU in transgenic plants was genotyped by PCR amplification of the target region using DNA extracted from their leaves, followed by sequencing to verify mutations. The primer sequences used for vector construction and amplification of the target region are listed in Table S1.
Assay of Oxalate Oxidase (OxO) Activity
OxO activity was carried out according to Zhang et al. (1996). Fresh tissue (about 5 mg) was homogenized using liquid nitrogen and the powder used for determination of OxO activity. The assay mixture contained the powder, 40 mmol/L succinic acid/NaOH buffer, pH 3.8, 60% ethanol (V/V), 0.4 mmol/L oxalic acid, 0.025% N, N-dimethylaniline, 0.1 mg/mL 4-aminoantipyrine and 5 U/mL of horseradish peroxidase. The mixture was incubated at room temperature for 6 min and trichloroacetic acid (0.1%) was added to terminate the reaction, then the absorbance of the supernatant was measured at 555 nm after the mixture were centrifuged at 12,000 g for 3 min at 4℃. OxO activity was determined as the amount of H_2_O_2_ (µM) produced in 1 g tissue per min. For in-gel OxO activity staining, proteins were extracted from leaves with 50 mmol/L Tris-HCl buffer (pH = 7.5) and separated in 7.5% native-PAGE. After gel electrophoresis, the gel was immersed in a staining solution containing 5 U/mL peroxidase (POD), 2 mmol/L oxalate, 0.5 mg/mL 4-chloro-1-naphthol, 60% ethanol (V/V) and 40 mmol/L succinic acid/NaOH buffer (pH 3.8) at room temperature until visible bands of enzyme activity could be observed.
Protein Immunoblotting
The proteins were extracted from the leaves of rice seedlings using 50 mmol/L Tris-HCl buffer (pH 7.5). The extracts were mixed with 2×SDS loading buffer (100 mmol/L pH = 6.8 Tris-HCl buffer containing 20% glycerol, 4% SDS, 4% mercaptoethanol and 0.04% bromophenol blue) before the proteins in the extracts were separated using 12.5% SDS-PAGE. After electrophoresis, the proteins were then transferred to nitrocellulose / PVDF membranes. Anti-His-OsOxO4 prepared in our laboratory was used as the primary antibody at 1: 2000 dilution. Horseradish peroxidase (HRP)-conjugated anti-rabbit (BBI D110011) was used as the secondary antibody. Immunodetection was performed using an ECL Enhanced Plus Kit (ABclonal, Wuhan, China) and detected using a ChemiScope 6000 Touch chemiluminescence imaging system (Clinx).
Results
A pGA2715 T-DNA Vector Tagged 270 bp Upstream of OsOxO2 Resulted in Enhanced Levels of OsOxO1, OsOxO2, OsOxO3 and OsIAAGLU Transcripts
To elucidate the physiological functions of oxalate oxidase (OxO) in rice, a T-DNA (Transfer DNA) insertion line (PFG_3A-02040.L) associated with the Os03g0693800 locus (OsOxO2) obtained from the Rice Functional Genomic Express Database (http://signal.salk.edu/cgi-bin/RiceGE) was screened. To determine the T-DNA insertion site in PFG_3A-02040.L, PCR amplification and sequencing were performed with LB (primer specific to the left border of the inserted T-DNA pGA2715) and RP (downstream primer specific to the flanking sequence of the expected insertion site in the native OsOxO2 gene) as primers, which revealed that the T-DNA was inserted 270 nucleotides upstream of the OsOxO2 ATG start codon (Figure S1 and Fig. 1A). PCR amplification using rice genomic DNA as the template and primers sets LB + RP, together with LP (upstream primer specific to the flanking sequence of the expected insertion site in the native OsOxO2 gene) + RP showed the progenies of PFG_3A-02040.L 9 − 3, 10 − 4 and 10 − 5 appeared to be homozygous mutants (Fig. 1B), because amplicon only with the LB + LP combination was observed and no amplicon with the LP + RP combination in the above lines. Subsequently, the 10 − 4 mutant, named herewith as lc4, was used in following experiments. Compared to WT, the transcript levels of OsOxO1, OsOxO2 and OsOxO3 (Fig. 1C) were significantly elevated in the leaves of lc4, OxO activity also increased remarkablely in lc4 mutants and represented 20-fold increase compared to WT controls (Fig. 1D). However, OxO activity gel staining in native PAGE and immunoblot assay (Figure S2) showed OsOxO2 is the predominant OxO isoform present in lc4 mutant leaves, because only one band was present in lc4 and the electrophoretic mobility of the band was similar to that of in OsOxO2OE/DJ. Additionally, the expression of OsIAAGLU, a gene encoding an IAA-conjugating enzyme that generates IAA-glucose by conjugating free IAA to glucose (Yu et al. 2019) was also upregulated in lc4 (Figs. 1A and E).
Fig. 1T-DNA insertion in OsOxO2 increased OxO activity and OsIAAGLU transcript levels. (A) schematic diagram of the T-DNA insertion site in PFG_3A-02040.L and the genes in its flanking genomic region, arrowheads indicate the transcriptional direction of each gene. (B) confirmation of the T-DNA insertion in PFG_3A-02040.L and identification of the homozygous lines by PCR amplification using genomic DNA and primers sets (LB+RP and LP+RP). (C) semi-quantitative RT-PCR analysis of OsOxO1-4 transcription levels in the leaves of wild-type (DJ) and lc4 seedlings. (D) OxO activity in DJ and lc4, data represent as means ± SD (n=5). (E) relative OsIAAGLU transcript levels in DJ and lc4, calculated as described in Materials and Methods, data are shown as mean ± SD (n = 4). The means±SD assigned with asterisks indicate significant differences between mean values by Student’s t-test (P<0.0001)
The Activation of OsOxO2 in Rice Resulted in Increased Leaf Angle and Semi-dwarfism Phenotype
Compared with WT rice plants, lc4 showed increased leaf angle and decreased plant height at 4-leaf-stage (Figure S3A-C) and the differences were more obvious at the booting stage (Fig. 2A-C). Moreover, the flag leaf angle and adaxial distance in the lamina joint of lc4 were increased, while the abaxial distance in the lamina joint was reduced (Figure S3D-E). However, the culm of lc4 was shorter than that of WT plants and the length of each internode was uniformly reduced in lc4 plants (Fig. 2C). Additionally, the length of panicle was also decreased compared to WT (Fig. 2D). Because the expression of OsIAAGLU, encoding an IAA-conjugating enzyme, was increased in the leaves of lc4, the sensitivity to NAA in lc4 mutant was investigated in a root elongation assay, the results showed the growth inhibition induced by exogenous NAA in roots of lc4 seedlings was decreased in comparison with WT plants (Fig. 2E), suggesting that modified plant architecture of lc4 might related to IAA. To explore that the increased leaf inclination and semi-dwarf phenotype of lc4 was due to the T-DNA insertion, the co-segregation analysis was carried out by analyzing the phenotype of progeny from heterozygous PFG_3A-02040.L mutant 3A9-2. Plants with T-DNA insertion and high level of OxO activity all exhibited increased leaf inclination and semi-dwarf phenotype, whereas those without insertion resembled WT (Figure S4). These results indicated that the T-DNA insertion was linked to enhanced OxO activity as well as alterations in plant architecture.
Fig. 2. Characterization of the phenotype of the lc4 T-DNA insertion mutant of rice. (A) the phenotype and flag leaf of representative wild-type (DJ) and the lc4 mutant plants. (B) quantification of leaf angle of plants depicted in (A). (C) the length of the internodes (n≥18), and panicle length (D) of DJ and lc4 plants (n=21) at 120 days from sowing. (E) the phenotype and root length of DJ and lc4 seedlings treated with Kimura B complete nutrient solution containing NAA kept in the dark for 5 d. “-” and “+” represent 0 and 10 nmol/L NAA. The means±SD assigned with asterisks indicate significant differences between mean values by Student’s t-test (P<0.0001). The means±SD of root length assigned with different letters were significantly different as determined by one-way analysis of variance (ANOVA) with Duncan test (P <0.05)
Transgenic Rice Plants Overexpressing OsIAAGLU, but not OsOxO2, Displayed Phenotype Resembling lc4
The insertion of T-DNA caused upregulated expression of both OsOxO1-3 and OsIAAGLU (Fig. 1C and E). Since the T-DNA was inserted into the promoter of OsOxO2 while OsIAAGLU has been shown to significantly regulate plant architecture (Yu et al. 2019), we focused on investigating whether OsIAAGLU and OsOxO2 would contribute to the alterations of plant architecture in lc4. For this purpose, we examined the architecture in plants overexpressing OsIAAGLU or OsOxO2. Overexpression of OsIAAGLU (driven by a constitutive promoter, pUbi: OsIAAGLU) in WT of the DJ background resulted in significantly exaggerated leaf angles (Fig. 3A and B) and decreased plant height (Fig. 3A and C) compared to WT. However, overexpression of OsOxO2 in WT of the DJ background, unlike its increased expression in lc4, only slightly increased leaf angle and decreased plant height compared to WT (Fig. 3), although there was a remarkable increase in OxO activity in the leaves (Figure S2). These results suggest that overexpressing OsIAAGLU, but not those of OsOxO2, resulted in enhanced leaf angle and dwarfism, suggesting that the increased leaf angle and dwarfism phenotypes of lc4 are mainly related to an increase in OsIAAGLU expression in lc4, rather than the elevated expression levels of OsOxO2.
Fig. 3. Transgenic rice plants overexpressing OsIAAGLU, not OsOxO2, in the wild-type background of rice variety Dongjin (DJ) displayed phenotype similar to lc4. (A) the phenotype of representative plants of DJ, the lc4 mutant, those overexpressing OsIAAGLU (lines #3 and #5) or OsOxO2 (lines #2 and #3) and flag leaf at the heading stage. (B) and (C) quantification of leaf angle at the filling stage and plant height at the maturation stage. The means ± SD of leaf angle and plant height assigned with different letters were significantly different as determined by one-way analysis of variance (ANOVA) with Duncan test (P < 0.05)
Partial Restoration of WT Phenotype by Mutation of OsIAAGLU in lc4
To investigate whether the increased expression of OsIAAGLU in lc4 is directly associated with the altered plant architecture in lc4, we mutated OsIAAGLU in lc4 by the CRISPR-Cas9 system (Figure S6A). The leaf inclination as well as plant height in the transgenic plants were significantly different from those of lc4 and appeared to be partially restored back to the WT phenotype (Fig. 4), which confirms that OsIAAGLU plays a major role in affecting the plant height and leaf angle phenotype of lc4. Although transgenic plants with different mutations in OsIAAGLU of lc4 (iaaglu/lc4) had reduced levels of OsIAAGLU transcripts in leaves compared to lc4, the levels were still significantly higher than that of WT (Fig. 4E). This indicates that the transcription of OsIAAGLU is influenced by other factors. To further confirm the role of OsIAAGLU in regulating the lc4 phenotype, we examined OxO protein, OsOxOs transcripts and OxO activity in iaaglu/lc4. The results showed that OxO protein, OsOxOs transcripts and OxO activity in the leaves of iaaglu/lc4 together with plant height at 3-leaf seedlings stage were not significantly different from those in lc4 (Fig. 4D, F, G and Figure S5A). In addition, the OxO protein levels remained unchanged in IAAGLUOE/DJ (Fig. 4D). All the above data not only suggest that the expression level of OsIAAGLU does not influence OxO activity and the transcripts of OsOxO1-4 which are co-localized on the same chromosome, but also further confirm that the lc4 phenotype is directly regulated by the expression of OsIAAGLU rather than that of OsOxOs.
Fig. 4. Mutation of OsIAAGLU or OsOxO2 in the lc4 background led to restoration of wild-type (WT) phenotype. (A) phenotype at the heading stage and leaf angle at the filling stage of representative plants of WT (DJ), the lc4 mutant, iaaglu/lc4 (lines #4 and #5), and oxo2/lc4 (lines #2 and #4). (B) quantification of leaf angle of plants depicted in (A). (C) plant height at the maturation stage. (D) immunoblot analysis of OxO protein levels in the leaves (second leaves from top to bottom) of 5-leaf stage wild type (DJ), the lc4 mutant, oxo2/lc4 (lines #2 and #4), iaaglu/lc4 (lines #4 and #5), OsOxO2OE/DJ (lines #3) and IAAGLUOE/lc4 (lines #3 and #5). Ponceau-stained ribulose-1, 5-bisphosphate carboxylase/oxygenase large subunit (RbcL) is shown as a loading control. (E) the expression levels of OsIAAGLU in WT, lc4, oxo2/lc4 (#2 and #4), iaaglu/lc4 (#4 and #5) and OsOxO2OE/DJ (#2 and #3) grown under natural sunlight to the 4-leaf stage, quantitative data was calculated using the formula in Materials and Methods and are presented as mean ± SD (n = 4). (F) the expression levels of OsOxO1, OsOxO2, OsOxO3, and OsOxO4 in the same genotypes as in (E). (G) OxO activity in the same genotypes as in (E), quantitative data of OxO activity are shown as means ± SD (n ≥ 4). The means ± SD of leaf angle, plant height, OsIAAGLU transcripts and OxO activity assigned with different letters were significantly different as determined by one-way analysis of variance (ANOVA) with Duncan test (P < 0.05)
Restoration of WT Phenotypes by Mutation of OsOxO2 in lc4
Besides mutation of OsIAAGLU in the lc4 background partially restored WT phenotypes, these plants maintained comparable OxO activity along with transcript levels of OsOxO1, OsOxO2,* OsOxO3* and OsIAAGLU with lc4. Therefore, OsOxO2 was mutated in lc4 by the CRISPR-Cas9 system (Figure S6A). Compared to WT plants, the leaf angle and plant height in oxo2/lc4 were mostly restored (Fig. 4B and C). Additionally, compared to lc4, in the leaves of oxo2/lc4, not only was OsOxO2 transcript significantly reduced, but OxO activity, OxO protein, and the transcripts of OsOxO1, OsOxO3 and OsIAAGLU were also significantly decreased to the levels similar to those in WT (Fig. 4D-G), while T-DNA were present in the original position (Figure S6B). These results suggest that OsOxO2 may regulate the OsOxO1, OsOxO3 and OsIAAGLU expression, thereby modulating rice plant height and leaf inclination. However, in the leaves of OsOxO2OE/DJ, no significant changes were detected in the transcripts of OsIAAGLU, OsOxO1 and OsOxO3 (Fig. 4E and F), indicating that ectopic expression of OsOxO2 does not affect the transcripts of OsIAAGLU, OsOxO1 and OsOxO3. Based on the above results, we propose that the regulatory effect of OsOxO2 may be related to its chromosomal position relative to these three genes. Ectopic expression of OsOxO2 in WT had no effect on transcripts of OsIAAGLU, OsOxO1 and OsOxO3, but significantly increased OxO protein levels and OxO enzymatic activity in leaves (Fig. 4D and G), indicating that OsOxO2-encoded protein possesses OxO activity.
Discussion
The Increase in OsIAAGLU Transcript Level Plays a Predominant Role in Alterations of Plant Architecture in lc4
Rice genome presents four tandem repeat OxO genes (LOC4333791-LOC4333794 and herewith referred to as OsOxO1-4) on chromosome 3 coding for OxO. Similar sequence and the very close genomic location for OsOxO1-4 suggest their functional redundancy and duplication. Here, we identified a OxO2 T-DNA mutant with significantly increased OxO activity and alternations in plant architecture. Compared to wild-type (WT) plants, leaf angle of lc4 increased at the 4-leaf-stage and more significantly at the booting time. Similarly, the differences in plant height between WT and lc4 became more obvious at the booting time than at the seedling stage. Moreover, the phenotype of enlarged leaf angle and semi-dwarfism in the co-segregated progeny that also had higher OxO activity and T-DNA insertion than the progeny from the heterozygous PFG_3A-02040.L mutant 3A9-2. Because cell wall OxO might be involved in the development from an immature cell wall to a more rigid, mature cell wall by promoting peroxidase activity in the presence of a supply of hydrogen peroxide (Wakabayashi et al. 2011). OxO is localized within cell type which is characteristically restricted in the extent of cellular enlargement. For example, during wheat seed germination, OxO is found in tissues such as coleorhiza, coleoptile, scutellum and vascular bundles. This is consistent with the hypothesis that the biological function of OxO is to restrict cell growth by participating in cell-wall restructuring through the localized H_2_O_2_ provision for cross–linking of wall components (Caliskan and Cuming 1998). Auxin response factors (ARFs), OsARF6 and OsARF17 are highly expressed in lamina joint tissues. The osarf6 and osarf17 mutants displayed an exaggerated flag leaf angle due to the reduced secondary cell wall deposition of the lamina joint sclerenchymatous cells (Huang et al. 2021a). Asymmetric cell division and elongation as well as cell wall development on the adaxial or abaxial sides of lamina joint can bring about leaf angle changes (Xu et al. 2021). Therefore, aberrant plant architectural phenotype of lc4 was originally thought to be due to an increase in OxO expression. In the transgenic plants overexpressing OsOxO2 in the DJ (WT) background, the levels of OsOxO2 transcript and OxO activity were increased greatly, but their phenotype was only slightly similar to lc4. In the transgenic plants of the DJ background overexpressing OsIAAGLU, which is in close proximity to OsOxO1 on the chromosome, the increased level of OsIAAGLU transcript was also accompanied with phenotypes including the decreased plant height, sensitivity to NAA and enlarged leaf inclination similar to lc4. Likewise, decreased plant height and panicle length in transgenic rice overexpressing OsIAAGLU were also reported by Choi et al. (2012), while decreased plant height and increased leaf angle in rice overexpressing OsIAAGLU by reducing IAA content was reported by Yu et al. (2019). Conversely, with mutation of OsIAAGLU in lc4, the phenotype of the iaaglu/lc4 resembled WT, although there were still significant differences between WT and iaaglu/lc4 as far as plant height and leaf angle were concerned. In addition, iaaglu/lc4 had comparable levels of OxO activity as well as OsOxO1 and OsOxO2 transcripts with lc4, suggesting OsIAAGLU might play a predominant role in alteration of lc4 phenotypes. Strikingly, the phenotypes including leaf angle and plant height of the plants (oxo2/lc4) with mutation of OsOxO2 in the lc4 background were restored close to those of DJ. In addition, the levels of OsIAAGLU, OsOxO1 transcript and OxO activity in the leaves of oxo2/lc4 were comparable with those in DJ, suggesting that OsOxO2 also plays an important role in the regulation of plant height and leaf angle. Taken together, the enhanced expression of OsIAAGLU and OsOxO2 in lc4 resulted in altered plant architecture. It appeared that OsIAAGLU could play a direct role in the regulation of plant architecture, and a regulatory relationship might exist between OsOxO2 and OsIAAGLU.
OsOxO2 Might Be Involved in Regulating OsIAAGLU, OsOxO1 and OsOxO3 in a Chromosome position-dependent Manner
Activation tagging is a method to generate dominant mutations in plants or plant cells by random insertion of a T–DNA carrying constitutive enhancer element, which can cause transcriptional activation of flanking plant genes (Memelink 2003; Jeong et al. 2002). Rice nicotianamine synthase gene (OsNAS3) transcript levels were 60- and 30-fold higher in the leaves of seedlings from 2 independent activation-tagged alleles OsNAS3-D1 (35 S enhancer elements were inserted approximately 1.5 kb downstream of an ORF of OsNAS3) and OsNAS3-D2 (the enhancer elements were inserted approximately 1.9 kb downstream of the OsNAS3 ORF), respectively, compared with WT (Lee et al. 2009). In this study, a T-DNA, i.e. pGA2715, was inserted at 270 bp upstream of OsOxO2 CDS. Therefore, the enhanced OxO activity and expression of OsOxO2 in the lc4 mutants might be related to the 4 copies of 35 S enhancers which were inserted next to the left border of T-DNA pGA2715. Likewise, 4 of the 10 candidate genes harboring pGA2715 and the enhancers within 4.5 kb displayed elevated expression (Jeong et al. 2002). However, transcript levels of OsOxO1 and OsOxO3 which are neighboring genes of OsOxO2 were also increased significantly in the lc4 mutants. In addition, OsIAAGLU with its 3′ UTR being 468 bp away from the 3′ UTR of OsOxO1 also showed enhanced expression in lc4, which seemed to play a predominant role in the altered plant architecture. Strikingly, mutation of OsOxO2 in lc4 reduced the expression of OsIAAGLU, OsOxO1 and OsOxO3 as well as OxO activity in the leaves of oxo2/lc4 to the same level as those in DJ, suggesting a regulatory relationship might exist between OsOxO2 and OsOxO1, OsOxO3, OsIAAGLU. However, the transcripts of OsOxO1, OsOxO3 and OsIAAGLU are not up-regulated in the leaves of transgenic plants of DJ background overexpressing OsOxO2 and with significantly increased OxO activity. Moreover, in these overexpression transgenic plants, there was no significant auxin deficiency phenotype which was present in lc4, IAAGLUOE/DJ plants, while there only slightly enlarged leaf angle and decreased plant height. Interestingly, in Drosophila melanogaster, Acsx1L (CG6300) and Acsx1R (CG11659) are tandem duplicates of a putative acyl-CoA synthetase gene, and Acsx1L expression is substantially reduced upon deleting the right-hand duplicated block Acsx1R (Loehlin et al. 2022). Moreover, tandem duplication often results in more than double the gene activity (Loehlin and Carrolla 2016). In view of the relative position of OsIAAGLU, OsOxO1, OsOxO2 and OsOxO3 on chromosome 3, we would speculate that OsOxO2 might affect the levels of OsOxO1 and OsIAAGLU transcripts in a position-dependent manner. Mutation in OsOxO2 might influence the expression of OsOxO1 and OsOxO3, and the change in expression of OsOxO1 might then influence the expression of OsIAAGLU. Because gain or loss of the accessible flanking chromatin regions (ACRs) and mutation of cis-regulatory elements (CREs) within ACRs can change the balance of the expression level and/or tissue specificity of the duplicated genes (Fang et al. 2023). Moreover, the genes that are clustered closely in the genome often tend to be co-regulated and exhibit correlated expression, while neighboring genes might also show correlated expression change (Ghanbarian and Hurst 2015). It has been suggested that the expression of one gene could directly impact on the expression of its neighboring genes in the immediate vicinity and this effect is stronger for tandem duplicates which are often co-regulated by shared genomic elements (Lan and Pritchard 2016). However, the underlying mechanism of chromosome position-dependent gene expression changes is still elusive and remains to be explored further.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1. Figure S1 PCR product sequence amplified from lc4 mutant using leaf genomic DNA as template and LB (T-DNA-specific) and RP (OsOxO2 specific) as primers. The blue-highlighted bases represent the 270-nucleotide region upstream of the OsOxO2 start codon (ATG). Figure S2 T-DNA insertion in 270 nucleotides upstream from the OsOxO2 start codon (ATG) resulted in enhanced OxO activity and OsOxO2 abundance. (A) OxO isoforms in rice leaves were separated by 7.5% CN-PAGE and visualized by in-gel OxO activity staining. Leaf extracts were analyzed from the following plants: wild-type Zhonghua 11 (ZH11) and Dongjin (DJ), transgenic plants overexpressing (OE) OsOxO1, OsOxO2, OsOxO3, or OsOxO4 in the ZH11 background, and OsOxO2OE plants in the DJ background, the lc4 mutant and hybrid plants generated by crossing OsOxO1OE and OsOxO2OE (ZH11 background) lines. (B) immunoblot analysis of proteins separated by 12.5% SDS-PAGE. Leaf extracts from the same genotypes as in (A) were probed with an anti-OsOxO4-His antibody. Figure S3 Characterization of the lc4 T-DNA insertion mutant at the 4-leaf stage and flag leaf lamina joints at the booting time. (A) phenotype of the 4-leaf stage representative rice variety Dongjin (DJ) wild-type and lc4 plants. (B) and (C) quantification of leaf angle and plant height of plants depicted in (A). (D) adaxial/abaxial phenotype of flag leaf lamina joints at the booting time. (E) quantification of adaxial/abaxial distance depicted in (D). Data are presented as means ± SD. Asterisks indicate statistically significant differences (Student’s t-test, P < 0.0001). Figure S4 Co-segregation analysis of the phenotype of the progeny from heterozygous PFG_3A-02040.L mutant 3A9-2: (A) phenotypes of progeny of heterozygous PFG_3A-02040.L mutant 3A9: 3A9-3 was homozygous, and its OxO activity in the leaves was higher than 3A9-2 which was heterozygous. There was no T-DNA insertion in 3A9-1 and no OxO activity was detected in its leaves. (B) primers sets (LP + RP and LB + RP) were used for genotyping part of progeny lines of the heterozygous PFG_3A-02040.L mutant 3A9-2 by PCR amplification using genomic DNA as template. (C) OxO activity in leaves and leaf angle (D) of progeny lines of the heterozygous PFG_3A-02040.L mutant 3A9-2 at the 4-leaf-stag. Figure S5 Mutation of OsIAAGLU or OsOxO2 in the lc4 background can restore its leaf angle phenotype similar to 3-leaf-stage wild-type (DJ) seedlings. (A) phenotypes of whole plants and leaf angle of representative seedlings. (B) and (C) quantification of leaf angle and plant height of seedlings depicted in (A). Figure S6 Identification of iaaglu and oxo2 mutant plants in lc4 background. (A) the sgRNA target region of oxo2/lc4 and iaaglu/lc4. The numbers on the bases represent its position in OsIAAGLU or OsOxO2 CDS, the inserted base is marked with red. (B) primer sets (LP + RP and LB + RP) were used for genotyping part of oxo2/lc4 and iaaglu/lc4 by PCR amplification using leaf genomic DNA as template. Table S1 Primer sequences used for PCR and vector construction.
The reference list from the paper itself. Each links out to its DOI / PubMed record.