Harnessing entomopathogenic nematodes for sustainable pest management: mechanisms, challenges, and innovations
Amandeep Kaur, David Kihoro Sirengo, Pratibha Karki, Thomas O. Powers, Amanda M. V. Brown

TL;DR
This paper reviews how entomopathogenic nematodes can be used for sustainable pest control, focusing on their biology, mechanisms, and challenges in large-scale application.
Contribution
The paper provides an integrative synthesis of molecular mechanisms and formulation strategies for entomopathogenic nematodes.
Findings
EPNs use symbiotic bacteria to infect and kill insect pests.
Advances in formulation and stress tolerance mechanisms have improved EPN field performance.
High production costs and environmental constraints limit large-scale commercialization of EPNs.
Abstract
Entomopathogenic nematodes (EPNs) of the genera Heterorhabditis and Steinernema are increasingly recognized as potent biological control agents due to their ability to infect and kill diverse insect pest taxa through a symbiotic partnership with insect-pathogenic bacteria. Over the last decades, substantial progress has been made in improving EPN field performance through advances in formulation and application methods, use of biodegradable polymers and nanocarriers, and elucidation of stress tolerance mechanisms. However, despite their proven efficacy, large-scale commercialization of EPNs remains limited by high production costs, formulation instability, and environmental constraints. While numerous reviews have separately addressed EPN biology, mass production, or field application independently, a critical and integrative synthesis linking molecular mechanisms, and formulation…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
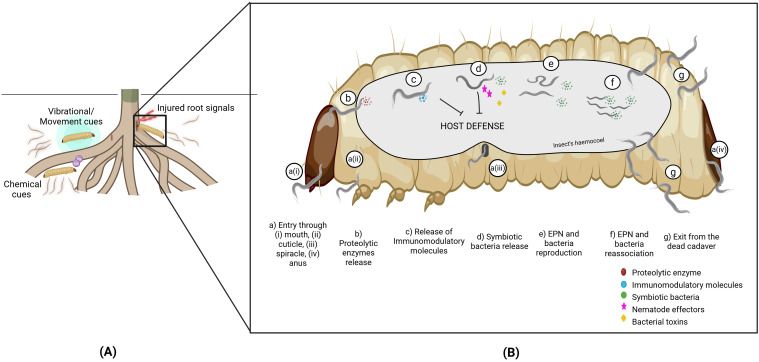 Figure 1
Figure 1| Formulation type | Adjuvant/additive | EPN species formulated | Mechanism of success (physiological/physical) | Mechanism of failure/limitation | Future optimization priorities | References |
|---|---|---|---|---|---|---|
| Activated charcoal gels | Activated carbon powder, wetting agents, buffers |
| Porous carbon matrix suspended nematodes, reduced sedimentation and helped adsorb toxic metabolites in storage. | Limited shelf-life and low instability at room temperature. | Integrate humectants and antioxidants, optimize carbon:nematode ratios | ( |
| Synthetic sponge | Polyether-polyurethane sponge sheets or cubes |
| The porous sponge matrix provides a large surface area that promotes oxygen exchange and maintains moisture, mimicking the nematode’s natural soil habitat. Sponge perforations limit sedimentation and support IJ survival during storage. | Recovery of nematodes from the sponge matrix is difficult, and formulation density is constrained by sponge pore capacity, thereby limiting scalability and uniform field application. | Develop biodegradable or dissolvable sponge matrices and optimize their pore size to improve IJ loading and release | ( |
| Wheat gluten granules (‘Pesta’) | Wheat flour, bentonite, kaolin, peat, 0.2% formaldehyde. | Wheat flour/gluten matrix slowly removed free water and induced quiescence and preserved lipid reserves within granules. | Low survival rate after 6 weeks of storage at 21 °C. Moreover, high humidity and flour matrix favored fungal and or bacterial growth. | Explore alternative cereal matrices less supportive of saprophytes, improve moisture control, and add antifungal and antibiotic agents. | ( | |
| Calcium alginate beads | Glycerol (18%), CaCO3·2H2O, CaCl2 (0.3-3% w/v), gluconolactate, xanthan gum. | solid Ca2+ alginate beads supplemented with glycerol retain IJs during storage and ensure a steady release when applied in soil. Glycerol-induced metabolic arrest in IJs result in quiescence and total retention of IJs when added to beads made with 0.5% sodium alginate and 2% CaCl2·2H2O solutions. | Dense or oversized beads can limit oxygen diffusion and nematode escape, leading to hypoxia and low field release. | Optimize bead size and porosity, design beads for mechanized delivery like banding or in-furrow applications. | ( | |
| Water-dispersible granules | Diatomaceous earth, | Controlled desiccation in granules induces partial anhydrobiosis, therefore extending shelf-life at moderate temperatures. This formulation achieved 90% survival for 6 weeks at room temperature. | Rehydration can cause osmotic shock if too rapid and some species lose infectivity after long-term desiccation | Improve drying-rehydration protocols, include osmoprotectants, and improve granule dispersibility | ( | |
| Polyacrylamide gels | Anhydrous polyacrylamide, vermiculite | Hydrophilic polymer retains water and physically immobilizes IJs, preventing sedimentation and mechanical damage. | The gels are difficult to dissolve, and IJs have a short survival time at room temperature. | Develop degradable gels to enhance IJs release | ( | |
| Infected cadavers and coated infected cadavers | Kaolin-starch mixture, unflavored gelatin, inert clay, protective powders | Host cadaver protects IJs from desiccation and UV. The coatings further reduce mechanical damage and enhance handling. | Cadavers can rupture or mold during storage, and producing them at large scale is bulky and labor-intensive. | Automate cadaver coating and engineer delayed emergence storage methods | ( | |
| Protective gel-coating (Barricade® fire gel) | Polyacrylate fire gel | Highly hydrated gel layer protects IJs from UV and desiccation on foliage, which extends the survival window for host invasion. | High viscosity can increase pump pressure and shear stress in spray equipment. | Optimize gel concentrations for lower shear while maintaining protection of IJs. Redesign low-shear nozzle systems if needed. | ( | |
| Oil-in-water emulsions (vegetable oil) | Vegetable oils, emulsifiers, non-ionic surfactants, stabilizers | Oil droplets reduce cuticular water loss from IJs and can improve persistence on exposed surfaces. | Unstable emulsions or phase separation can concentrate nematodes in oil-rich phases and reduce oxygen availability. | Optimize oil:water ratio for oxygen and sprayability and stabilize emulsions across hard water and agitation regimes | ( | |
| Chitosan additives | Chitosan matrices | A chitosan matrix can enhance spray retention and may stimulate plant defenses, thereby indirectly supporting EPN persistence and performance. | The matrix’s solubility depends on pH. Moreover, poor dissolution or precipitation can cause clogging and uneven nematode distribution. | Standardize chitosan chemistry and buffer to a stable pH. Also, test blends with compatible polymers for better sprayability and release | ( | |
| Carboxymethyl cellulose (CMC) gels | Sodium polyacrylate, carboxymethyl cellulose, vermiculite coating, inorganic salts (NaCl, KCl, MgSO4). | CMC gel holds moisture around IJs and can be applied as a targeted gel plug on plants (e.g. maize whorl). It also slows evaporation and sedimentation. | Too high CMC concentration reduces sprayability; gel can still dry out under extreme heat without additional protectants and hard water can affect performance. | Optimize viscosity with humectants and UV protectants for application methods and validate performance under realistic field relative humidity/temperature conditions | ( | |
| Titanium Pickering emulsion (TPE) | TiO2 nanoparticles, mineral/vegetable oil | TiO2 nanoparticles in oil-in-water Pickering emulsions form a reflective coating that reduces UV damage and water loss while maintaining viability. | Nanoparticle aggregation or inappropriate oil:water ratios can reduce coating uniformity and harm IJs survival. | Optimize nanoparticle size and surface chemistry | ( | |
| Silica-Pickering emulsion gel (SPEG) | Silica nanoparticles, oil droplets, gelled aqueous phase | Silica-based Pickering gel slows desiccation and maintains IJs on foliage for extended periods under solar exposure. | Formulation cost and sensitivity to water chemistry can limit large-scale adoption at present. | Develop cheaper silica/clay hybrids and simplify mixing for end users | ( | |
| Potassium polyacrylate hydrogel (PPH) | Potassium polyacrylate superabsorbent polymers, other optional fillers | Superabsorbent hydrogel releases water gradually in soil, helping IJs disperse from the application point. | High salinity or fertilizer ions can collapse the gel and reduce water-holding capacity. | Formulate for salt tolerance; blend with organic carriers to buffer ions | ( | |
| Diatomaceous earth (DE) pellets | Diatomaceous earth (Celite 209), hygroscopic clays, binders. | Porous DE matrix controls evaporation and can induce partial anhydrobiosis while providing mechanical protection. | Non-uniform pore size and low moisture can over-dry some IJs, reducing post-rehydration infectivity. | Standardize porosity and moisture of DE pellets; add moisture buffers like SAP; optimize rehydration procedures | ( | |
| Liquid-core alginate capsules | Alginate shell, Ca²+ crosslinker, gluconolactate, buffers, and optional viscosity modifiers. | The liquid core keeps IJs active while the alginate shell protects them from desiccation and permits targeted root-zone release. | Capsules can leak or allow premature IJ escape during storage, and IJs have a limited shelf-life at room temperature. | Incorporate multilayer shells or coatings to minimize leakages | ( | |
| Bait formulations (edible gels) | Alginate-based edible gels + phagostimulants | Edible calcium-alginate gels incorporate phagostimulants to promote ingestion by the target insect larvae which are less exposed to surface penetration via gut route. | Gels can desiccate and lose attractiveness which reduces palatability; field performance is sensitive to bait placement and pest behavior. | Improve moisture retention by adding humectants; design pest-specific attractants to promote ingestion; integrate the baits with monitoring to time placement | ( | |
| Hygroscopic attapulgite clay ‘sandwich’ granules | Attapulgite clay layers | Layered attapulgite clay and nematode slurry induced controlled desiccation and partial anhydrobiosis for storage with survival time of 8 weeks at 23 °C. | Short shelf-life, nozzle clogging in spray equipment, and low nematode-clay proportion led to the discontinuation of this commercial product. | Re-engineer the formulation as soil-applied granules rather than sprayable | ( | |
| Hydrogenated vegetable oil paste (microgels) | Hydrogenated oils, mono-/diglycerides | A hydrophobic matrix slows water loss and buffers thermal fluctuations during shipment of IJs. | Low oxygen diffusion through the paste can limit IJ survival under suboptimal storage. | Introduce microchannels or porous fillers to improve oxygen circulation within the paste | ( | |
| Pre-infected live hosts (living insect bombs) | Live insect hosts as delivery cargo |
| Live infected hosts can move and actively deliver IJs into cryptic habitats before nematode emergence and help in delivering inoculum directly to target zones. | Biologically complex multi-organism rearing makes it disadvantageous; timing when to apply and regulatory concerns limit practical use. | Develop predictive timing models to enhance efficacy; restrict application to niche pests like borers where benefits justify complexity | ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEntomopathogenic Microorganisms in Pest Control · Polymer-Based Agricultural Enhancements · Insect Resistance and Genetics
Introduction
1
The global population is projected to increase significantly by 2050 (United Nations, 2017), warranting a substantial increase in crop productivity through sustainable intensification and the adoption of innovative pest management strategies to ensure global food security. In this context, as agriculture shifts toward ecologically informed approaches, biological control has emerged as a cornerstone of environmentally sound, sustainable pest management. This paradigm shift has paved the way for the exploitation of naturally occurring beneficial microorganisms as alternatives and complements to synthetic chemical pesticides. As a result, the adoption of entomopathogenic nematodes (EPNs) as biocontrol agents against major agricultural insect pests is increasing across diverse cropping systems worldwide (Modic et al., 2020; Fallet et al., 2024; Polat et al., 2024; Ramakuwela et al., 2025). The successes of EPNs as biological control agents have been well demonstrated in various cropping systems against different insect taxa. In particular, EPNs have been effective in controlling stem flea beetle (Psylliodes chrysocephala) in winter oilseed rape (Godina et al., 2023), Pumpkin fruit fly (Bactrocera tau) in Chinese cucumber (Liu et al., 2025), false codling moth (Thaumatotibia leucotreta) in citrus (Moore et al., 2024), and codling moth (Cydia pomonella) in apple (Odendaal et al., 2016) among others. Moreover, compatibility of EPN with synthetic insecticides has been evaluated primarily through invitro bioassays, revealing species- and formulation-specific responses (Krishnayyaand and Grewal, 2002; Srivastava et al., 2011; Sabino PH de et al., 2014; Mahmoud et al., 2016; Wu et al., 2017; Dias S da et al., 2024). However, chemical compatibility with formulation components and application strategies needs to be tested under realistic agroecosystem conditions.
Beyond their practical applications, EPNs also serve as model organisms for advancing fundamental research on microbial mutualism and ecological interactions (Campos-Herrera et al., 2012; Tarasco et al., 2023; Stock et al., 2025). Notably, among EPN taxa, the infective juveniles (IJs) of Heterorhabditis and Steinernema, which maintain obligate symbioses with gammaproteobacteria of the genera Photorhabdus and Xenorhabdus, respectively, are the most intensively studied due to their effectiveness in biocontrol and relevance in both research and commercial applications. Within this highly specialized partnership, the nematode provides physical protection for its bacterial symbiont against environmental stresses such as temperature changes, moisture loss, and harmful microbes, and then releases the symbiont into the insect hemocoel upon infection (Stilwell et al., 2018). Once inside the host, these symbionts produce secondary metabolites and various toxins (Eleftherianos et al., 2007; Hinchliffe et al., 2010; Tobias et al., 2017) that suppress immune defenses and kill the host through septicemia, thereby creating a conducive environment for the nematode’s feeding and reproduction in the cadaver, facilitating later reacquisition of the bacteria before nematodes emerge as new IJs (Tarasco et al., 2023; Glazer et al., 2025). The insecticidal, antimicrobial and immunomodulatory roles of these secondary metabolites have been comprehensively reviewed elsewhere (Bode, 2009; Stock et al., 2017; Liao et al., 2025). Their application, however, is limited by host dependence and unresolved toxicity concerns, underscoring the need for systematic evaluation of stability, safety, and delivery strategies. Traditionally, this partnership has been viewed as species-specific, with each nematode species maintaining a single coevolved symbiont. Experimental evidence shows that these partnerships are tightly governed by partner recognition and fidelity and that pairing nematodes with non-cognates reduces fitness, virulence, and bacterial carriage (McMullen et al., 2017). Nonetheless, recent studies revealed the coexistence of other bacterial species with the primary symbiont. These secondary or facultative associates may enhance infective juvenile (IJ) insecticidal activity and expand the nematode’s host range (Pervez et al., 2020; Maher et al., 2021; Sugiyama and Hasegawa, 2025).
In addition to causing insect mortality, application of EPN in soil has been shown to modulate plant antioxidant defenses by influencing the activities of key enzymes such as peroxidases and catalase (Jagdale et al., 2009). For instance, the chemical cues such as ascarosides and 2,3-butanediol released by EPN-symbiont complex trigger oxidative bursts in root tissues, leading to activation of salicylic acid and jasmonic acid signaling pathways (Jagdale et al., 2009; Manohar et al., 2020). These hormonal cascades stimulate the expression of defense-related genes like PR-1 (Helms et al., 2019; Wang et al., 2025) and enhance enzymatic scavenging of reactive oxygen species through increased peroxidase and catalase activity. These multitrophic interactions reflect the ecological versatility of EPNs and their growing importance in integrated pest management (IPM) frameworks. Their effectiveness can be further augmented through co-application with other biocontrol agents, such as entomopathogenic fungi, where synergistic interactions frequently result in enhanced pest suppression and increased crop productivity (Sáenz-Aponte et al., 2020; Půža and Tarasco, 2023). The remarkable successes of EPNs across diverse cropping systems demonstrate their potential as reliable biological control agents and alternatives to chemical pesticides. However, realizing their full potential requires overcoming challenges related to formulation, shelf life, field persistence, regulatory policies, and large-scale delivery systems (Blanco-Pérez et al., 2025). Several comprehensive reviews have examined EPNs with emphasis on EPN biology, mass production, application, field efficacy and challenges (Shapiro-Ilan and Dolinski, 2015; Koppenhöfer et al., 2020; Abd-Elgawad, 2023; Tarasco et al., 2023). However, most existing reviews treat EPN biology, symbiosis, formulation strategies as largely independent topics. This review aims to bridge this gap by evaluating how molecular mechanisms underlying host immune suppression, stress tolerance affects field performance and commercialization.
Molecular mechanisms of EPNs on insect infection
2
Host localization, foraging strategies, and invasion
2.1
Successful infection by EPNs relies on host localization mediated by chemical, physical, and other environmental cues (Figure 1A) (O’Halloran and Burnell, 2003; Ali et al., 2010; Hallem et al., 2011; Filgueiras et al., 2016; Laznik and Trdan, 2016; Baiocchi et al., 2017; Laznik et al., 2020; Zhang et al., 2021). Among the important chemical cues are the paired BAG sensory neurons that detect olfactory signals such as CO_2_, a byproduct of insect respiration. In Heterorhabditis bacteriophora, BAG neuron ablation abolishes chemotaxis toward the greater wax moth, Galleria mellonella, confirming the essential role of BAGs in host localization (Hallem et al., 2011). BAG neurons also contribute to nictation-triggering circuits in ambusher EPN species like Steinernema carpocapsae (Hallem et al., 2011). In addition to insect-derived signals, EPNs also respond to herbivore-induced plant volatiles (HIPVs) through chemosensory neurons (Rasmann et al., 2005; Degenhardt et al., 2009). Root-derived compounds such as carvone have been shown to activate S. carpocapsae, suggesting that plant volatiles can trigger specific chemosensory responses in these nematodes (Bai et al., 2024). Although the underlying neuronal circuits in EPNs remain poorly characterized, evidence from the model nematode, Caenorhabditis elegans, suggests that attraction and repulsion behaviors are mediated by complex interneuronal networks (Hart and Chao, 2010; Guillermin et al., 2017).
Schematic representation of (A) host-finding and (B) infection by entomopathogenic nematodes. Figure was created using BioRender.
While these sensory mechanisms drive host detection, the specific cues used for host localization can vary depending on the EPN’s foraging strategy. Cruiser EPN species (e.g., H. bacteriophora) track long-range chemical, vibrational, and electrical signals, while ambushers (e.g., S. carpocapsae) use nictation to intercept mobile hosts (Lortkipanidze et al., 2016; Grunseich et al., 2021). Electrical fields generated by insect movement or root injury can elicit species-specific directional responses (Rasmann et al., 2005; Grunseich et al., 2021). For instance, the cruiser Steinernema glaseri moves toward higher electrical potentials, while the ambusher S. carpocapsae is attracted to lower potentials (Shapiro-Ilan et al., 2009; Shapiro-Ilan et al., 2012). Seismic cues produced by insect movement help detect hosts, particularly when chemical gradients are disrupted in organic-rich soils (Torr et al., 2004).
Following localization, host invasion is a critical and often challenging step in the infection process. EPNs utilize multiple routes to enter their hosts, and penetration routes vary among insect species. The choice of penetration pathway is both species- and host-specific (Batalla-Carrera et al., 2014; Kallali et al., 2024). Typically, EPNs invade through natural openings such as the mouth, spiracles, or anus, but some species can also directly penetrate the insect cuticle (Dowds and Peters, 2002).
Overcoming the insect immune system
2.2
While penetration marks the beginning of an infection process, the survival of EPNs within their hosts depends on their ability to bypass host immunity through coordinated interactions with their symbiotic bacteria (Figure 1B) (Ciche and Ensign, 2003; ffrench-Constant et al., 2007). Upon invasion, IJs of H. bacteriophora use their anterior tooth to pierce host tissues while secreting proteolytic enzymes that degrade structural barriers. On the other hand, Steinernema species degrade midgut epithelia with enzymes before they can enter the hemocoel (Ciche and Ensign, 2003; Balasubramanian et al., 2010). Immediately after penetration into the hemocoel, the IJs shed their second-stage cuticle, exposing a dynamic surface coat that masks immunogenic epitopes and releases small immunomodulatory molecules (Wang and Gaugler, 1999), an early immune evasion strategy that suppresses host recognition and prepares the hemocoel for bacterial release and establishment.
Once in the hemocoel, IJs release effector molecules via their excretory-secretory glands, which target both humoral and cellular immunity. Among these effectors, some of the most important examples include proteolytic enzymes that suppress the prophenoloxidase (pro-PO) cascade, therefore blocking melanization and nodulation. Similarly, a Kunitz-type protease inhibitor (Sc-KU-4) disrupts hemocyte encapsulation and aggregation (Balasubramanian et al., 2010; Chang et al., 2019). As a result, transient immune paralysis is induced, halting the host defense response and enabling the establishment of IJs. This process creates favorable conditions for the subsequent release and proliferation of the EPN’s symbiotic bacteria within the host (Lu et al., 2017). Once released from the nematode gut, the symbionts reinforce and extend host suppression through multiple molecular pathways. For instance, the symbiotic bacterium Xenorhabdus nematophila inhibits Toll and IMD (immune deficiency) signaling by suppressing eicosanoid biosynthesis, thereby halting the production of antimicrobial peptides (AMPs) such as cecropins and lysozymes (Park and Kim, 2000; Hwang et al., 2013).
Moreover, extracellular proteases from Xenorhabdus degrade pre-existing AMPs, hence enhancing humoral suppression (ffrench-Constant et al., 2007; Balasubramanian et al., 2010). Similarly, the symbionts Photorhabdus luminescens and X. nematophila secrete rhabduscin. This phenoloxidase inhibitor suppresses melanization and Mcf-family toxins and induces death of hemocytes, the primary immune cells in the hemolymph of invertebrates responsible for defense against pathogens, foreign entities, and gut epithelial cells (Dowling et al., 2004; Crawford et al., 2012). Serralysin-type proteases such as PrtA further degrade immune-related proteins, thereby enhancing systemic infection (Felföldi et al., 2009). These events later lead to a collapse of humoral and cellular defenses, resulting in rapid septicemia in the target host (Forst et al., 1997; da Silva et al., 2020).
Interestingly, several studies have demonstrated the pathogenicity of axenic EPNs under laboratory conditions. For instance, Ehlers et al. (1997) investigated the pathogenicity of the Steinernema feltiae-Xenorhabdus bovienii complex to the marsh crane fly, Tipula oleracea, and G. mellonella larvae by injecting dauer IJs using monoxenic nematodes (with bacteria), axenic nematodes (bacteria-free), and the symbiotic bacteria alone. Axenic IJs were less pathogenic than monoxenic IJs in both insects, indicating that bacterial symbionts significantly enhance virulence. Similarly, when axenic cultures of G. mellonella larvae were treated with axenic IJs of H. bacteriophora and S. carpocapsae, all larvae were killed by S. carpocapsae, whereas H. bacteriophora failed to cause mortality despite a few nematodes being recovered from the hemocoel (Han and Ehlers, 2000). Although these results represent partial pathogenic success, the nematode-bacterium association remains essential for complete pathogenicity and successful reproduction (Bisch et al., 2015). These results support the potential of EPNs as candidate model systems for studying the molecular mechanisms of parasitism and insect immune defense.
Stress tolerance mechanisms and environmental persistence challenges
3
Despite advances in the biology, development, and deployment of EPNs, their integration into IPM programs is constrained by interrelated challenges ranging from formulation and application to environmental limitations. The following sections highlight the stress tolerance and environmental persistence mechanisms, and the challenges of formulation and application constraints that affect EPN-based biocontrol.
Stress tolerance and environmental persistence
3.1
EPNs encounter both abiotic and biotic stresses in the environment. Among these, extreme temperatures, desiccation, and UV radiation represent key abiotic challenges, while microbial antagonists constitute major biotic threats. The field efficacy of EPNs as biological control agents largely depends on their ability to withstand diverse biotic and abiotic stresses. The environmental persistence is further determined by stress tolerance mechanisms that may be species- or strain-specific as well as by interactions with soil type and the availability of suitable hosts. IJs acclimate to thermal stress by accumulating trehalose (Grewal and Jagdale, 2002; Jagdale and Grewal, 2003; Grewal et al., 2006) and by reducing metabolic activity (Brown and Gaugler, 1996; Andalo et al., 2011; Ali and Wharton, 2013). Although IJs possess mechanisms to lower their lipid reserves, species-specific differences exist in how metabolic adjustments occur. For instance, Heterorhabditis megidis reduces production of proteins involved in metabolism and protein synthesis, whereas H. carpocapsae downregulates proteins involved in intermediary metabolism and oxidative phosphorylation (Jagdale and Grewal, 2003; Andalo et al., 2011).
Under severe dehydration, IJs enter a state of anhydrobiosis, and osmoregulant molecules such as, heat-stable, water-stress-related protein (DESC47), and heat-shock protein (HSP40) are synthesized (Solomon et al., 2000; Gal et al., 2003; Somvanshi et al., 2008). In S. feltiae, desiccation induced a two-fold increase in trehalose content (Solomon et al., 1999). Furthermore, the induction of casein kinase (CK2) has been observed, which is hypothesized to participate in the transcriptional activation of nucleosome-assembly protein (NAP-1) (Gal et al., 2003). While UV tolerance mechanism in EPNs is unclear, the general assumption is that UV tolerance mechanisms parallel those described in C. elegans. In C. elegans, UV light is detected and avoided through four sensory neurons (ASJ, ASK, AWB, and ASH) with the neurotransmitter glutamate receptors (glc-3, mgl-1, mgl-2) expressed in ASH and ASK. These neurons play key roles in mediating this response (Bargmann, 2006; Ozawa et al., 2022).
In addition to species- and strain-specific physiological stress responses, ecological adaptation plays a critical role in determining EPN persistence and field efficacy. Locally adapted beneficial microbes exhibit superior tolerance to changing weather and climatic conditions, enhanced dispersal, and form a more stable association with the native soil microbiota (Campos-Herrera et al., 2013). Native EPNs have been shown to exhibit strong virulence against pests for major crops and turf grasses (Yadav and Lalramliana, 2012; Sun et al., 2021; Thakur et al., 2022; Tumialis et al., 2023; Fallet et al., 2024; Koppenhöfer and Luiza Sousa, 2024) and show greater potential when integrated with chemical control strategies (Shields et al., 2018; Tomar and Thakur, 2022). In addition, the EPN-symbiont complex produces a myriad of natural compounds that include scavenger deterrent factors (SDFs) with antifungal, insecticidal, and antibacterial properties. These metabolites not only deter scavengers from colonizing insect cadavers but also help the EPNs and their symbiotic bacteria overcome host immune defenses, ensuring successful infection and reproduction (Furgani et al., 2008; Gulcu et al., 2012; Fang et al., 2014; Hazir et al., 2016; Tobias et al., 2017; Gulcu et al., 2018).
Formulation and application
3.2
Successful commercialization of EPNs depends on their formulation and handling. However, one major obstacle limiting their field performance arises during formulation (Chen and Glazer, 2005). To increase EPN shelf life and efficacy, EPN treatments have been formulated using several carriers such as liquid concentrates, alginate gels, water-dispersible granules, synthetic sponge, and vermiculite applied in aqueous suspension through agricultural sprayers or irrigation systems (Georgis, 1990; Grewal, 2000) (Table 1). Traditionally, early work on water-dispersible granules showed enhanced improvements in EPN longevity and ease of handling compared with aqueous formulations (Grewal, 2000). Nonetheless, most aqueous products require refrigeration to maintain IJ viability, which results in increased production and transport costs and limits use in regions without reliable cold storage (Chen and Glazer, 2005; Karimi et al., 2025).
EPN-infected cadavers have been explored as natural carriers (Shapiro and Glazer, 1996; Shapiro-Ilan et al., 2001). The IJs emerging from infected hosts disperse efficiently through soil, are highly infective, and provide better pest control than nematodes applied in suspension (Shapiro and Lewis, 1999; Perez et al., 2003; Shapiro-Ilan et al., 2003). For instance, field trials using H. bacteriophora-infected cadavers effectively reduced infestations of the sweet potato weevil, Cylas formicarius, and the guava weevil, Conotrachelus psidii. The reliability of cadavers as a dispersal agent, however, is limited because they desiccate and rupture during production and storage. These challenges were addressed by formulating cadavers with protective materials that prevent sticking and mechanical damage (Shapiro-Ilan et al., 2003). However, the influence of cadaver age on stability and efficacy remains unclear.
Recent advancements concentrate on improving stability in ambient conditions. For example, Darsouei et al. (2025) demonstrated that several EPN species encapsulated in calcium-alginate beads survived up to three months at room temperature under laboratory conditions. Similarly, hydrogel and emulsion capsules were shown to improve persistence of S. feltiae against the yellow mealworm beetle, Tenebrio molitor, under laboratory and field conditions (Perier et al., 2025). Other studies have demonstrated that encapsulation in alginate gels and hydrophilic colloids also shields IJs from desiccation and UV stress (Kim et al., 2021). Additionally, carboxymethyl cellulose (CMC), a water-soluble cellulose derivative, has emerged as an effective substrate for EPN formulation, particularly for S. carpocapsae (Kary et al., 2021). Despite these advances, cost-effective commercialization remains a challenge (San-Blas, 2013; Ramakuwela et al., 2016). Even with successful formulations, effectiveness ultimately depends on proper handling and field application.
During application, IJs are introduced to stressors such as tank sedimentation (Schroer et al., 2005), nozzle shear or pressure damage (Moreira et al., 2013), and post-application desiccation mortality (Platt et al., 2018). Several adjuvants and surfactants have been formulated recently to improve efficiency. The addition of xanthan gum or potassium alginate resulted in a two-fold increase of insect mortality at 80% relative humidity (RH) and a five-fold increase at 60% RH, while the mixtures of 0.3% xanthan or alginate with 0.3% surfactants further improved efficacy for foliar application of S. carpocapsae against diamondback moth larvae (Schroer et al., 2005). Similarly, S. carpocapsae formulated in an alkyl polyglycoside polymeric surfactant reduced the tomato pinworm, Tuta absoluta, larvae populations within two weeks post-treatment (Kajuga et al., 2025). Nonetheless, the efficacies of these adjuvants or surfactants are not consistently reliable. Some, for instance, increase longevity and enhance host invasion, whereas others reduce infectivity (Beck et al., 2013).
Future directions and opportunities
4
Given the growing demand for safer food and environmentally sustainable pest management practices, the commercial potential of EPNs is expanding (Dara, 2019). However, unlike chemical pesticides, their commercialization is constrained by multiple factors (Koppenhöfer et al., 2020). Addressing these limitations requires a stronger focus on biological and ecological factors that influence EPNs’ performance. Systematic screening of native EPN isolates, coupled with formulation optimization could further enhance persistence while reducing reliance on protective encapsulants.
Beyond strain selection, interactions among EPNs, their bacterial symbionts, and soil microbial communities may influence EPN efficacy. Nevertheless, the underlying mechanisms remain poorly understood. Therefore, metagenomic profiling of soil microbial community in soils with contrasting EPN efficacies can identify microbial taxa that promote EPN success or suppress their activity and enable the design of synthetic microbial communities that can be co-inoculated with EPNs to prime the soil environment, ensuring higher success rate in non-native soils. On the other hand, limited efficacy or genetic diversity in natural strains can be addressed through genomic-assisted breeding and molecular genetic approaches, including mutagenesis, transgenesis, and targeted gene modification. Over the past decades, these approaches have been applied to enhance EPN stress tolerance and longevity (Shapiro et al., 1997; Hashmi et al., 1998; Vellai et al., 1999; Sumaya et al., 2018). Selective breeding, however, can introduce fitness trade-offs, as demonstrated by altered reproduction, dispersal, and storage stability in modified S. carpocapsae strains (Gaugler et al., 1990; Bal et al., 2014). Future efforts to develop host-specific EPNs may extend their efficacy beyond soil-dwelling insects to targeted pests across various cropping systems (Lacey and Georgis, 2012). The application of precise and advanced genetic engineering technologies such as CRISPR/Cas9 offers a pathway to precisely enhance EPN environment resilience through targeted modification.
Recent advancements in IJ recovery through biodegradable polymers and nanocarriers are emerging as promising tools for enhancing the EPN performance and tolerance under various field stress conditions. For instance, alginate-starch hydrogel increased S. feltiae persistence and infectivity across different water regimes (Perier et al., 2025), while titanium-dioxide-based Pickering emulsions maintained S. carpocapsae viability for up to 120 days (Kotliarevski et al., 2022). Future research should prioritize precision-based delivery systems that synchronize nematode release with pest phenology and prevailing microclimatic conditions compared to conventional aqueous sprays. EPN application via drip irrigation systems resulted in more uniform nematode distribution, improved persistence, and significantly greater control of onion thrips compared with standard foliar sprays (Metwally et al., 2025). Emerging AI-driven automatic and controlled treatment systems could further optimize site-specific deployment via drones or automated irrigation, targeting pest “hotspots” and reducing costs and environmental impact. To maximize field efficacy, EPNs should be applied during early morning or late evening to reduce UV exposure, in pre-moistened soils to enhance IJ motility, and using methods that minimize mechanical shear. When combined with improved formulations and targeted genetic enhancements, such synchronized, precision-based delivery can elevate EPNs from supplemental inputs to reliable, site-specific components of modern IPM programs.
Concluding remarks
5
In summary, EPNs offer a promising avenue for sustainable pest management, providing a biologically robust alternative or complement to synthetic chemical control. Continued innovation in formulation, strain selection, and application technologies, supported by insights from genomics, nanotechnology, and microbial ecology, will be essential to enhance their persistence and reliability under field conditions. Omics-based discovery of genes and regulatory pathways in EPNs can guide more precise, targeted improvements in strain performance. However, potential trade-offs between enhanced traits should be considered. Future innovations should also focus on ecological interactions and development of strategies that integrate EPNs with beneficial microbes to enhance EPN persistence, infectivity, and strain stability in field deployment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abd-Elgawad M. M. M. (2023). Optimizing entomopathogenic nematode genetics and applications for the integrated management of horticultural pests. Horticulturae 9, 865. doi: 10.3390/horticulturae 9080865 · doi ↗
- 2Ali F. Wharton D. A. (2013). Cold tolerance abilities of two entomopathogenic nematodes, Steinernema feltiae and Heterorhabditis bacteriophora. Cryobiology 66, 24–29. doi: 10.1016/j.cryobiol.2012.10.004, PMID: 23142823 · doi ↗ · pubmed ↗
- 3Ali J. G. Alborn H. T. Stelinski L. L. (2010). Subterranean herbivore-induced volatiles released by citrus roots upon feeding by Diaprepes abbreviatus recruit entomopathogenic nematodes. J. Chem. Ecol. 36, 361–368. doi: 10.1007/s 10886-010-9773-7, PMID: 20309617 · doi ↗ · pubmed ↗
- 4Andalo V. Moino A. Maximiniano C. Campos V. P. Mendonca L. A. (2011). Influence of temperature and duration of storage on the lipid reserves of entomopathogenic nematodes. Rev. Colomb. Entomol. 37, 203–209. doi: 10.25100/socolen.v 37i 2.9075 · doi ↗
- 5Ansari M. A. Hussain M. A. Moens M. (2009). Formulation and application of entomopathogenic nematode-infected cadavers for control of Hoplia philanthus in turfgrass. Pest Manag. Sci. 65, 367–374. doi: 10.1002/ps.1699, PMID: 19165730 · doi ↗ · pubmed ↗
- 6Aquino-Bolaños T. Ruiz-Vega J. Hernández Y. D. O. Castañeda J. C. J. (2019). Survival of entomopathogenic nematodes in oil emulsions and control effectiveness on adult engorged ticks (Acari: ixodida). J. Nematol. 51, 1–10. doi: 10.21307/jofnem-2019-001, PMID: 31115202 PMC 6929640 · doi ↗ · pubmed ↗
- 7Bai P. H. Yu J. P. Hu R. R. Fu Q. W. Wu H. C. Li X. Y. . (2024). Behavioral and molecular response of the insect parasitic nematode Steinernema carpocapsae to plant volatiles. J. Invertebr. Pathol. 203, 108067. doi: 10.1016/j.jip.2024.108067, PMID: 38278342 · doi ↗ · pubmed ↗
- 8Baiocchi T. Lee G. Choe D. H. Dillman A. R. (2017). Host seeking parasitic nematodes use specific odors to assess host resources. Sci. Rep. 7, 6270. doi: 10.1038/s 41598-017-06620-2, PMID: 28740104 PMC 5524962 · doi ↗ · pubmed ↗