Overexpression of AtPROPEP6 enhances Arabidopsis thaliana resistance to Southern root-knot nematode Meloidogyne incognita
Payal Sanadhya, Kallahan Minor, Jiamei Li, Suxing Liu, Alexander Bucksch, Alisa Huffaker, Joanna Kud, Fiona L. Goggin

TL;DR
This study shows that overexpressing the AtPROPEP6 gene in Arabidopsis improves resistance to a specific type of nematode.
Contribution
The study identifies AtPROPEP6 as a novel gene involved in nematode defense in Arabidopsis.
Findings
AtPROPEP6 is specifically upregulated by nematode-derived ascr#18.
Overexpression of AtPROPEP6 reduces nematode gall formation and delays nematode development.
AtPROPEP6 overexpression correlates with elevated PR1a expression in Arabidopsis.
Abstract
Plant elicitor peptides (Peps), derived from PROPEP protein precursors, are stress-induced signaling molecules that enhance plant immunity. While previous studies of Pep-mediated immune signaling in Arabidopsis thaliana have focused on the roles of AtPROPEP1–3 genes in bacterial and fungal resistance, this study identifies the AtPROPEP6 gene as a contributor to defense against the Southern root-knot nematode (Meloidogyne incognita). In silico promoter analysis revealed enrichment of W box motifs, suggesting potential regulation by WRKY transcription factors associated with plant immune responses. Unlike other PROPEP gene family members, AtPROPEP6 shows specific upregulation in response to ascr#18, a nematode-derived molecular pattern, but not to other pathogen elicitors. Transgenic constitutive overexpression of AtPROPEP6 in A. thaliana significantly reduced gall formation and total…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
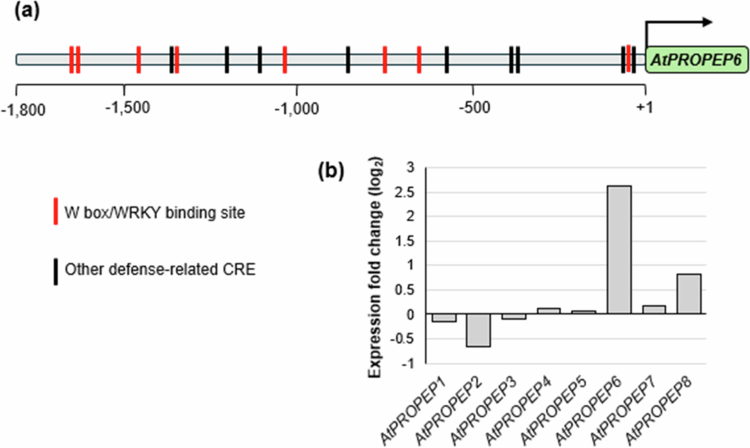 Figure 1
Figure 1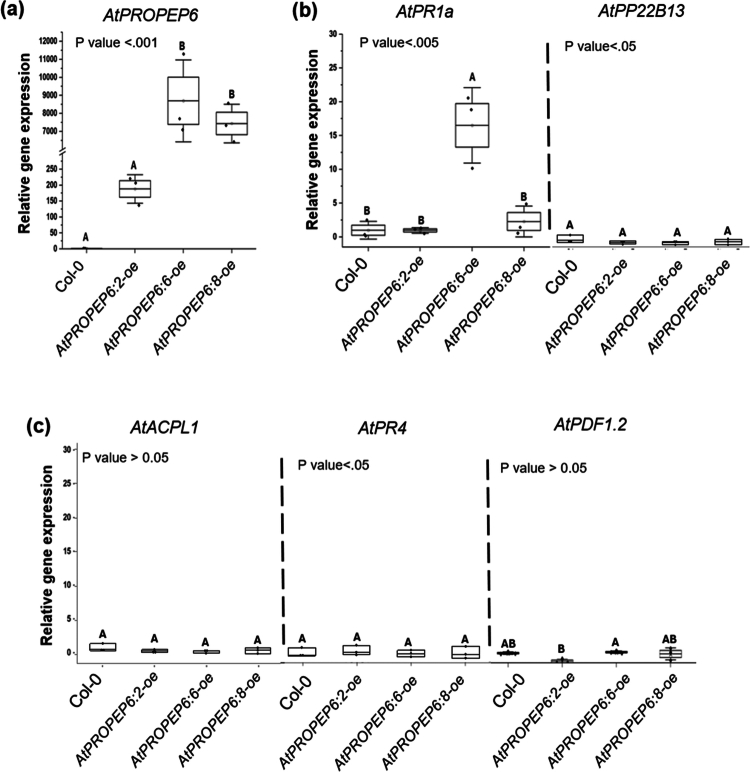 Figure 2
Figure 2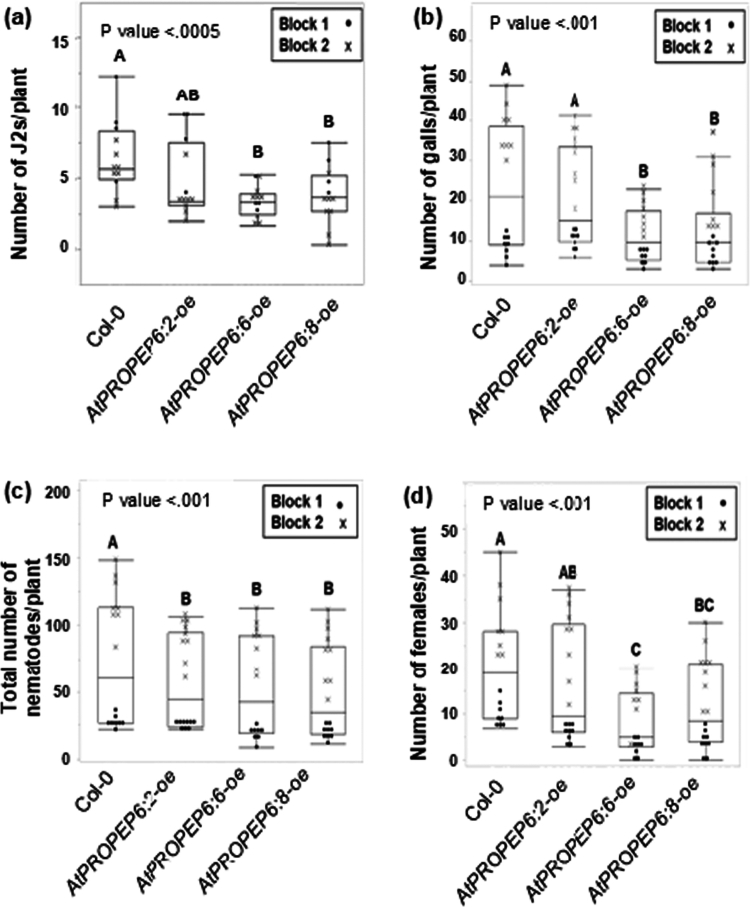 Figure 3
Figure 3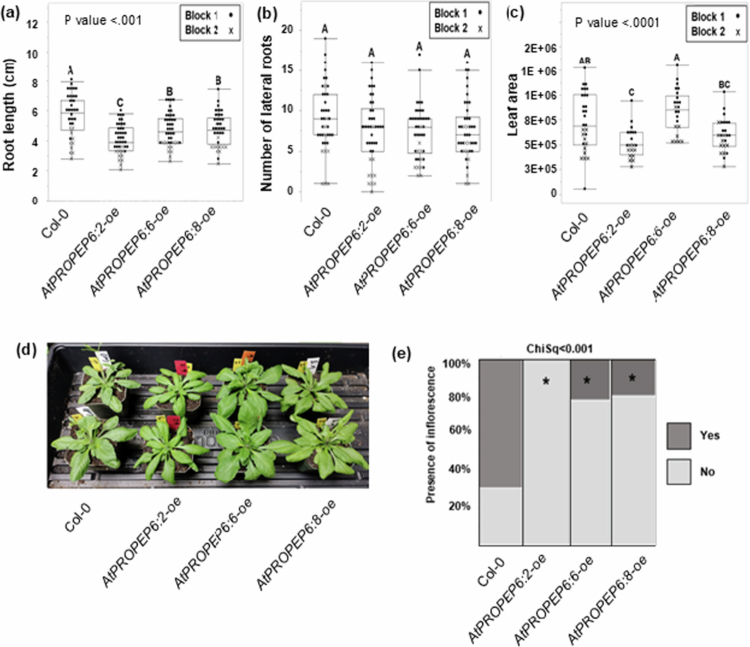 Figure 4
Figure 4- —Arkansas Agricultural Experiment Station10.13039/100007907
- —Arkansas Biosciences Institute10.13039/100008231
- —National Institute of Food and Agriculture10.13039/100005825
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNematode management and characterization studies · Polysaccharides and Plant Cell Walls · Plant-Microbe Interactions and Immunity
Introduction
Plant elicitor peptides (Peps) are endogenous molecules released upon pathogen attack or physical injury, commonly referred to as damage- or danger-associated molecular patterns (DAMPs).1^,^2 Peps were originally discovered in Arabidopsis thaliana, but are broadly conserved across both monocots and dicots, including key crops such as maize, soybean, tomato, potato, rice, sorghum, and canola.3^,^4 In their bioactive form, Peps are only 23 to 36 amino acids in length, however they originate from the C-terminal region of larger precursor proteins known as PROPEPs. Individual PROPEPs localize either in the cytoplasm or the tonoplast and are cleaved by the metacaspases upon stress stimulus.3^,^5 Then, Peps are actively or passively released to the apoplast, where they are perceived by transmembrane Pep elicitor receptors (PEPRs) belonging to the LRR-RLK family (Leucine-rich repeat receptor-like kinase), which initiate defense signaling cascades.6^,^7 Interestingly, Pep treatments elicit more robust immune responses in roots than the typical pathogen-associated molecular patterns (PAMPs), such as flagellin and chitin.8
Several studies have shown that Peps play a role in activating plant defenses against both the above- and belowground biotic stresses, including microbes, insects, and nematodes. In A. thaliana, overexpression of AtPROPEP1 reduced the necrotrophic oomycete Pythium irregulare infection,3 and foliar application of AtPep2/3 imparted resistance to the hemibiotrophic bacterial pathogen, Pseudomonas syringae pv. tomato DC3000.7 Conversely, loss of function of both Pep receptor genes (AtPEPR1 and AtPEPR2) increased susceptibility to foliar herbivory by the Spodoptera littoralis.9 In crops, ZmPep1 pretreatment protected maize from the fungal pathogens Colletotrichum graminicola and Cochliobolus heterostrophus,10 and soybean seed treatments with GmPep1, GmPep2, and GmPep3 reduced the soybean cyst nematode (SCN: Heterodera glycines) and the Southern root-knot nematode (SRKN; Meloidogyne incognita) infection.11^,^12
In A. thaliana, the genomic organization and expression patterns of the eight PROPEP genes (AtPROPEP1–AtPROPEP8 genes, encoding AtPep1-AtPep8 peptides) suggest functional differences between those paralogous. AtPROPEP2/1/3 and AtPROPEP8/7/4/5 form two distinct gene clusters on chromosome 5,3^,^13 and wounding or pathogen elicitors activate transcription of AtPROPEP2/1/3 but not AtPROPEP8/7/4/5. Moreover, within these groupings, individual AtPROPEP genes display distinct spatial and temporal expression patterns.13 AtPeps also differ in their ability to stimulate mutants with impairments in AtPEPR1 or AtPEPR2, implying different binding affinities for the two receptors.13 Furthermore, AtPeps function appears to extend beyond regulating plant immunity. A mutant with loss of function of AtPROPEP2 displayed a constitutive reduction in root hair growth, indicating a role for AtPEP2 in normal root development independent of stress responses.14
To date, relatively little is known about the role of AtPROPEP6. Whereas all other AtPROPEP genes are clustered on chromosome 5, AtPROPEP6 is a solitary member located on chromosome 2.3^,^13 Studies of the activity of all eight AtPROPEP genes in roots indicated that constitutive patterns of AtPROPEP6 are most similar to AtPROPEP1 and AtPROPEP2, with strong expression throughout root tissues and localization in the tonoplast.13^,^14 However, little information is available about AtPROPEP6 stress responses because most previous analyzes of the responsiveness of AtPROPEP family to stimuli have excluded this paralog.13^,^15 Based on this evidence for multifunctionality and potential functional diversification among Peps, it is important to characterize the impacts of each family member on both growth and defense.
To address this knowledge gap, the objective of this study was to determine if AtPROPEP6 plays a role in A. thaliana defense responses to M. incognita infection and to assess whether AtPROPEP6 contributes to host resistance independently of changes in root traits. Unlike other PROPEP genes, AtPROPEP6 is specifically induced by the nematode-derived signal ascr#18. Transgenic constitutive overexpression of AtPROPEP6 in A. thaliana enhanced resistance by reducing early nematode root penetration, nematode development, and root galling, effects that correlated with elevated AtPR1a expression rather than changes in root length. Our data support a role for AtPROPEP6 in modulating Pep-mediated immunity during nematode infection.
Materials and methods
Nematode cultures
Meloidogyne incognita was maintained on greenhouse grown tomato (Solanum lycopersicum cv. Rutgers), and nematode eggs were extracted from infected roots using 0.05% (v/v) sodium hypochlorite (NaOCl), followed by sucrose flotation.16 Eggs were surface sterilized by sequential treatment with 0.7% (v/v) streptomycin for 10 min and 0.01% (w/v) mercuric chloride for 10 min, followed by thorough rinsing with sterile distilled water. Sterile eggs were collected on 25-μm sieves and incubated in 0.01 M MES buffer (Sigma-Aldrich) in the dark for 3 days to allow hatching.
In silico promoter analysis
The 1,743 kb region upstream from AtPROPEP6 translation start site, which represents the entire intragenic region up to the next 5’ open reading frame, was used for cis-regulatory elements (CREs) identification with NewPLACE.17
AtPROPEP6 overexpression lines
The 35S:AtPROPEP6-oe lines were generated using the same methods as we previously described for 35S:AtPROPEP1-oe lines.3 Briefly, the expression cassette containing the AtPROPEP6 genomic sequence under the control of the cauliflower mosaic virus 35S promoter was constructed using the pART7/pBART binary vector system.18 A. thaliana (ecotype Columbia-0, Col-0) plants were transformed via the Agrobacterium tumefaciens-mediated floral dip method,19 and transgenic A. thaliana seedlings were selected on Murashige and Skoog media (MS; PhytoTech Labs, Lenexa, KS, USA) containing BASTA (10 μg/ml). The transgenic status of each line was confirmed both by BASTA selection and by PCR with pART F (5’ CTT CGC AAG ACC CTT CCT CTA 3’) and pART R (5’ CAT AGG CGT CTC GCA TAT CTC 3’) primers to amplify the inserted T-DNA region.20 AtPROPEP6 expression was compared in more than eight independent events using semi-quantitative RT-PCR as described before,20 and those with at least ten-fold higher expression than untransformed controls were selected for further propagation and analysis. Events were selfed, and segregation was tracked in subsequent generations using both BASTA selection and pART primers, through which we developed three non-segregating lines, each from a different independent event (35S:AtPROPEP6:2-oe, 35S:AtPROPEP6:6-oe, and 35S:AtPROPEP6:8-oe). Lines were passed through at least three generations before use for this study. An overview of the analyzes performed on these lines is presented in Figure S1.
RNA extraction and qRT-PCR
Rosette tissues from wild-type Col-0 and transgenic A. thaliana 35S:AtPROPEP6-oe plants cultivated in potting mix were collected 14 days after transfer to soil and flash-frozen in liquid nitrogen. Total RNA was extracted from the leaf tissue using RNeasy Plant Mini Kit (Qiagen, Germany), and mRNA was converted to cDNA with SuperScript III reverse transcriptase and oligo dT primers (Invitrogen, Carlsbad, CA, USA). Gene expression was determined by qRT-PCR (initial denaturation 95 °C for 5 min, followed by 40 cycles of 95°C/15 s, primer specific °C/30 s, and 72°C/30 s. A melting curve analysis was performed at the end of the PCR run over the range 60–95°C, increasing the temperature by 0.5 °C every 10 s. Gene expression was calculated using the ^ΔΔ^Ct method with values normalized to the geometric mean of expression levels of three stable endogenous reference genes: Actin2 (At3G18780), EF1A (At5G60390), and F-Box (At5g15710).21‐25 All primers, their corresponding annealing temperatures, and amplification efficiencies are listed in Table S1.
In vitro M. incognita infection assays
A. thaliana seeds were surface-sterilized by 5 min treatment in 70% ethanol, followed by five sterile distilled water washes, then 10 min in a 1.4% aqueous NaOCl solution. The seeds were then washed five times once again with sterile distilled water. Sterilized A. thaliana seeds were germinated on the Murashige and Skoog (MS; PhytoTech Labs, Lenexa, KS, USA) medium supplemented with 2% (w/v) sucrose and 0.8% Gelrite (Sigma-Aldrich, St. Louis, MO, USA). Germinated seedlings were transferred to individual Petri Dish plates after 7 days and plates were then grown vertically in a growth chamber (22 °C; 12 hrs. light, 12 hrs. dark). At 14 days after germination, each seedling was inoculated with either 300 or 500 freshly hatched, sterile M. incognita J2s, applied near the root zone for nematode penetration and galling assay, respectively. Each experiment had at least 8 biological replicates (n ≥ 8). Roots were gently harvested at either 3- or 28-days post-inoculation (dpi), stained with acid fuchsin,26 and observed under the stereomicroscope (Nikon SMZ800 microscope, Nikon Metrology Inc.) to visualize different nematode development stages and galls (Figure S1). Total female counts reported the number of both mature and young females, with examples of these stages shown in Figure S1d. Each experiment was repeated twice and included at least eight biological replicates per transgenic line.
Root morphology analysis
A. thaliana seeds were germinated and seedlings grown in tissue culture conditions on MS media as described earlier. Roots from 14-day-old plants were scanned using an EPSON Perfection V30 scanner (Epson America Inc., USA), and primary root length as well as the number of secondary roots were quantified with ImageJ software.27 The experiment was repeated twice with n = 10.
Shoot phenotypic analysis and bolting
A. thaliana seeds were germinated in tissue culture conditions on MS media for 7 days and then moved to pots with Sungro LC1 Sunshine potting mix (Sungro Horticulture, Belevue, WA) supplemented with Osmocote Plus 15-9-12 slow-release fertilizer (Scotts-MiracleGro Company, Marysville, OH). Plants were maintained in a growth chamber (22 °C, 12 hrs. light, 12 hrs. dark). After 14 days, plants were photographed from above to capture the area of the rosette; leaf area was calculated from images using the SMART tool.28 This timepoint was selected because it was the last point at which the plants could be imaged from above without extensive interference from the developing inflorescences. Bolting, defined as the transition from vegetative growth to reproductive development marked by elongation of the inflorescence stem, was assessed by counting the number of plants that had bolted (i.e., produced visible inflorescences) 21 days after transfer to soil. The experiment was repeated twice with n = 10.
Results
In silico promoter analysis of AtPROPEP6 and its expression in response to ascr#18
To address the relative lack of knowledge of AtPROPEP6 function, we first conducted an in silico promoter analysis for this overlooked family member. The 1,743 kb region upstream from AtPROPEP6 translation start site, which represents the entire intragenic region up to the next 5’ open reading frame, was used for cis-regulatory elements (CREs) identification with NewPLACE.17 The analysis revealed 19 defense-associated cis-regulatory elements (CREs), which accounted for approximately 8% of the total predicted transcription factor binding motifs (Figure S2). Notably, within that subset of immunity-linked CREs, we uncovered 8 binding sites for putative WRKY transcription factors (W box) (Figure 1a, Table S2). Given that prior in silico and in vivo analyzes of defense-related AtPROPEP1–3 promoters demonstrated that W boxes are critical for mediating their transcriptional upregulation in response to elicitor treatments,29 this finding suggests a potential role for AtPROPEP6 in plant defense. Additionally, our analysis of previously published RNAseq data obtained from A. thaliana Col-0 treated with ascr#18, a nematode-associated molecular pattern (NAMP), showed strong 6-fold induction of AtPROPEP6 compared with other AtPROPEPs displaying only weak or no induction at all 30 (Figure 1b). Building on these findings, we hypothesize that AtPROPEP6 may play an important role in plant defenses to phytonematodes.
Transcriptional regulation of AtPROPEP6 expression suggests an involvement in plant defenses. (a) Defense-related Cis-Regulatory Elements (CREs) in the promoter of the AtPROPEP6 gene. The positional distribution of predicted CREs is shown as vertical color-coded bars for individual binding sites (see also Table S2). Red–W box/WRKY binding site; black - other defense-related CREs. (b) Log2 fold change of AtPROPEPs expression in response to ascr#18 treatment. The expression pattern data were obtained from the published RNAseq analysis of Arabidopsis genes induced by ascr#18 treatment (1 µM ascr#18 treatment for 6 hours) -NCBI Bio Project Accession PRJNA550121. Numbers represent averages of 4 biological replicates, p-value < 0.05, FDR < 0.05.30
Overexpression of AtPROPEP6 uniquely impacts the expression pattern of marker genes
To assess the effects of AtPROPEP6 overexpression on nematode infection, we generated multiple independent overexpression events and selected three homozygous, single-insertion lines exhibiting varying levels of AtPROPEP6 transcript abundance for subsequent experiments. The AtPROPEP6:6-oe line had the highest transgene transcript accumulation, AtPROPEP6:8-oe showed moderate levels, and AtPROPEP6:2-oe had the lowest expression (Figure 2a). Previous studies have revealed that overexpression of AtPROPEP1 and AtPROPEP2 upregulates the expression of several defense-related genes, including salicylic acid (SA) marker genes (AtPR1a and AtPP22B13) and the ethylene and jasmonic acid (ET/JA) marker genes (AtPDF1.2, AtPR4, AtACPLA1).31^,^32 Interestingly, AtPR1a basal expression was significantly higher in the AtPROPEP6:6-oe line with the highest AtPROPEP6 transcript level. In contrast, the expression of other tested marker genes remained unchanged (Figure 2b and 2c). These results show that AtPep6 is capable of uniquely priming plant defenses before biotic challenge.
AtPROPEP6 overexpression increased the basal expression of the AtPR1a marker gene. The relative expression of (a) AtPROPEP6, (b) SA marker genes (AtPRa1, AtPP22B13), and (c) JA marker genes (AtACPL1, AtPR4, AtPDF1.2) in homozygous AtPROPEP6-oe lines (6:2, 6:6, and 6:8) was determined by qRT-PCR. Results are presented as the fold change in relative basal expression compared to non-transgenic control (Col-0). Different letters indicate statistically significant differences among treatments (one-way ANOVA followed by Tukey’s post hoc test; α = 0.05; n = 3).
Overexpression of AtPROPEP6 reduces M. incognita ability to penetrate roots and delays nematode development
To evaluate the impact of the AtPROPEP6 overexpression on both early and late stages of the M. incognita infection process, we examined nematode penetration at 3 dpi and nematode development at 28 dpi. At 3 dpi, significantly fewer juveniles had entered AtPROPEP6:6-oe and AtPROPEP6:8-oe than the untransformed control (Col-0), and nematode numbers on AtPROPEP6:2-oe were intermediate but not statistically different compared with the control (Figure 3a). Consistently, AtPROPEP6:6-oe and AtPROPEP6:8-oe also exhibited fewer galls at 28 dpi, a measure of root damage by nematode (Figure 3b). In addition to the total nematode count (all life stages) at 28 dpi (Figure 3c), we also quantified the number of females to assess effects on nematode development (Figure 3d; see Figure S1 for details). While total nematode numbers were reduced by 21%, 27%, and 32% in all overexpression lines compared to Col-0 control, the number of females was decreased more substantially by approximately 20%, 58%, and 45% in the AtPROPEP6:2-oe, AtPROPEP6:6-oe, and AtPROPEP6:8-oe lines, respectively. These results suggest that AtPROPEP6 overexpression delays nematode maturation, with the strongest effect observed in AtPROPEP6:6-oe, which also exhibited the highest accumulation of transgene transcripts. Collectively, these findings demonstrate that AtPROPEP6 overexpression not only limits nematode entry but also impairs their development within host roots.
AtPROPEP6 overexpression reduces Arabidopsis susceptibility to M. incognita infection. (a) Root penetration assay scored at 3 dpi, and (b–d) galling and nematode development assay scored at 28 dpi (including (b) number of galls, (c) total nematode count, and (d) number of females). Nematode infection assays were performed on three transgenic AtPROPEP6-oe lines (6:2, 6:6, and 6:8) and wild-type Col-0 as described in Figure S1. Data points were obtained from two independent experiments (Blocks 1 and 2) with a total of n = 16 (a) and n = 8 (b–d) for each line per experiment. Different letters indicate statistically different results between treatments (one-way ANOVA followed by Tukey’s post hoc test p < 0.05).
AtPROPEP6 overexpression influences plant morphology and bolting
In light of the considerable evidence supporting the idea that, PROPEP genes also influence plant growth and development, we next investigated whether overexpression of AtPROPEP6 results in changes to plant morphology. To assess belowground growth, which could potentially impact nematode infection, we measured the length of the primary root and the number of lateral roots in 14-day-old seedlings, the growth stage used for nematode inoculation in our infection assays. Overexpression of AtPROPEP6 resulted in significantly shorter primary roots across all three tested lines, although root length did not correlate with transgene expression. The AtPROPEP6:2-oe line, which had the lowest transcript levels, exhibited the most pronounced root growth inhibition, while AtPROPEP6:6-oe (highest expression) and AtPROPEP6:8-oe (intermediate expression) showed milder effects (Figure 4a and Figure S4). Despite differences in the length of the primary root, there were no significant differences in the number of lateral roots between wild-type and transgenic lines (Figure 4b), indicating that all lines had comparable numbers of root tips available for M. incognita infection. Lastly, because AtPROPEP1-oe and AtPROPEP2-oe were previously linked to increased aboveground growth, we also evaluated leaf area and weight at 14 days, as well as bolting at 21 days post transfer to soil. While AtPROPEP6:2-oe and AtPROPEP6:8-oe had reduced leaf area and weight compared to wild-type plants, AtPROPEP6:6-oe did not significantly differ from to wild-type control (Figure 4c-d, Fig S5); thus, once again, the effects of AtPROPEP6 overexpression on growth were not directly correlated with transgene expression levels or with nematode resistance.
Overexpression of AtPROPEP6 impacts root morphology and bolting. Belowground morphological measurements included (a) root length and (b) number of lateral roots in 14-day-old plants. Aboveground traits were recorded as (c–d) leaf area and (e) bolting at 21 days. For (a–c), data points were obtained from two independent experiments (Blocks 1 and 2). Different letters indicate significant differences (one-way ANOVA with Tukey's HSD, P < 0.05, n = 10). For (e), lines marked with an asterisk are significantly different from Col-0 control based on Chi-square test with a total of n = 10 for each line per experiment (p < 0.001).
Discussion
Peps are conserved proteinaceous signaling molecules that play important roles in plant stress responses and the regulation of both growth and development. In A. thaliana, the eight PROPEP paralogs differ in their genomic organization, expression patterns, and affinity to bind PEPR receptors, strongly supporting the notion that, despite some functional redundancy, substantial functional divergence exists among family members. Here, we focus on the previously largely uncharacterized AtPROPEP6, particularly in the context of its role in plant immune responses to M. incognita. Our findings expand the known set of defense-associated A. thaliana PROPEPs beyond AtPROPEP1-3, identifying AtPROPEP6 as a paralog involved in plant defenses to nematodes.
The available literature describes the basal expression of AtPROPEP6 as being most closely related to AtPROPEP1 and AtPROPEP2, with strong activity throughout root tissues,13^,^14 but comparatively little is known about stress-inducible regulation of this paralog. Transcriptional control of several AtPROPEP family members has been linked to WRKY transcription factors. In particular, Logemann et al. demonstrated that W-box cis-elements in the promoters of AtPROPEP2 and AtPROPEP3 are directly bound by AtWRKY33 upon defense elicitor treatments.29 AtWRKY33 is a key immune-related transcription factor involved in multiple defense pathways and has been shown to positively contribute to A. thaliana resistance to cyst nematodes; 33 however, its role in immunity against root-knot nematodes has not been directly tested. Nevertheless, other WRKY transcription factors are also implicated in plant-nematode interactions. For example, AtWRKY11 is induced by M. incognita infection and functions as a positive regulator of defenses against both root-knot and cyst nematodes.34 Given the enrichment of W-box motifs in the AtPROPEP6 promoter, we speculate that WRKY-mediated transcriptional regulation is a plausible mechanism controlling AtPROPEP6 expression during nematode infection and warrants further investigation.
Unlike AtPROPEP1–3, AtPROPEP6 is uniquely responsive to ascr#18 treatment, but it is not induced by methyl jasmonate, methyl salicylate, or other pathogen elicitors, including a necrosis-inducing Phytophthora protein (NPP1), Pseudomonas harpin protein HrpZ, or the most conserved domain of bacterial flagellin protein (flg22).3^,^30 This unique expression pattern suggests a potential role in plant-nematode interactions. Our analysis of the AtPROPEP6 overexpression lines revealed reduced susceptibility to M. incognita, manifested by a lower early penetration rate, which correlated with fewer galls and diminished nematode development at later infection stages. While total nematode numbers were reduced across all three lines, the decline in female counts was more pronounced, indicating that AtPROPEP6 expression interferes not only with host entry but also with nematode maturation. The strongest suppression of female development was observed in AtPROPEP6:6-oe, consistent with its highest transgene expression, suggesting a potential dose-dependent effect. Together, these results identify AtPROPEP6 as a contributor to nematode resistance that limits parasitic success within host roots at more than one stage of infection. However, the precise mechanisms underlying this effect remain to be elucidated.
Our data show that constitutive overexpression of AtPROPEP6 selectively elevates basal accumulation of AtPR1a transcripts, while having no significant effect on the other tested markers previously reported to be induced by defense-related AtPep1-3.31^,^32 Specifically, the absence of AtPDF1.2 induction in AtPROPEP6 overexpression lines contrasts with the elevated transcript accumulation of this marker gene observed upon overexpression of AtPROPEP1 or AtPROPEP2, further supporting the notion that AtPROPEP6 represents a functionally distinct member of the PROPEP family.31 This expression profile aligns with earlier studies showing that M. incognita infection of A. thaliana roots strongly induces *SA-*associated PR genes, including AtPR1a, whereas ET/JA-associated PR genes such as AtPR4 show no transcriptional response.35 M. incognita is a sedentary endoparasite that establishes a long-term biotrophic relationship with the host plant, making activation of SA-mediated defense pathways consistent with the nature of this interaction.32 Notably, AtPR1a appears to play an uniquely important role in nematode defense, as it was the only PR gene among RKN-induced SA markers (AtPR1, AtPR2, and AtPR5) whose constitutive overexpression in transgenic A. thaliana significantly suppressed M. incognita galling.35
Interestingly, exogenous application of AtPep1 caused dose-responsive, AtPEPR2-dependent inhibition of root elongation.8^,^36^,^37 On the other hand, constitutive overexpression of AtPROPEP1 and AtPROPEP2 resulted in visibly larger root and shoot biomass,3 and the AtPEPR2 receptor mutant, atpepr2, displayed a shorter root phenotype than wild-type plants.38 Thus, the relationship between Pep dosage and growth responses may be complex, given the conflicting reports on the effects of PROPEP overexpression or ectopic Pep treatment on plant growth. In this study, while AtPROPEP6-oe lines display slightly shorter primary roots than Col-0 control, this trait did not correlate with transgene expression in a similar manner to AtPR1a gene induction and impact on M. incognita infection. Unlike cyst nematodes, which can penetrate roots at variable positions, root-knot nematodes preferentially enter roots just behind the root tip, within the meristematic and elongation zones.39^,^40 Root tips are particularly permissive to root-knot nematode entry because these tissues exhibit minimal lignification, resulting in comparatively thinner and less rigid cell walls than those of mature taproot tissues.41 Consequently, differences in overall root length are unlikely to substantially influence infection rates, especially given that root tip availability remained unchanged in the AtPROPEP6 overexpression lines. Additionally, the observed reductions in leaf area and weight could in part be due to developmental delays because all three lines, but especially AtPROPEP6:2-oe, were delayed in the onset of bolting compared to wild-type controls. The combined results of our bioassays and morphological measurements indicate that AtPROPEP6 impacts both nematode infection and plant growth, but the effects of AtPROPEP6 overexpression on nematodes do not seem to be correlated with or dependent upon gross morphological changes in the plant. Rather, the effects of AtPROPEP6 overexpression are likely attributed to enhanced immune signaling.
Although Pep–PEPR signaling relies on a broadly conserved receptor-based recognition mechanism, apart from a few C-terminal residues that mediate binding to the LRR domain of PEPR receptors.42 The remaining non-conserved regions are highly disordered, making it difficult to define clear orthologous relationships across species. Peps exhibit striking sequence divergence. Even within Arabidopsis, individual paralogs share only 12–47% sequence identity.6 Consequently, Pep paralogs often carry out distinct and non-redundant biological functions.43 Furthermore, Pep-PEPR signaling represents highly species-specific communication, which is supported by experiments showing that A. thaliana PEPR1 cannot perceive maize ZmPEPs, and vice versa.43 Hence, a comprehensive understanding of the Pep–PEPR system requires independent functional characterization of individual paralogs and putative orthologs, as their distinct biological roles cannot be inferred from sequence alone. Thus, characterizing AtPROPEP6 in the context of nematode stress and impact on plant morphology provides a meaningful step toward deciphering the complexity of this signaling network.
Supplementary Material
SUPPLEMENTARY MATERIALS.docxSUPPLEMENTARY MATERIALS.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Endo S, Betsuyaku S, Fukuda H. Endogenous peptide ligand-receptor systems for diverse signaling networks in plants. Curr Opin Plant Biol. 2014;21:140–146. doi: 10.1016/j.pbi.2014.07.011.25113600 · doi ↗ · pubmed ↗
- 2Macho AP, Zipfel C. Plant PR Rs and the activation of innate immune signaling. Mol Cell. 2014;54(2):263–272. doi: 10.1016/j.molcel.2014.03.028.24766890 · doi ↗ · pubmed ↗
- 3Huffaker A, Pearce G, Ryan CA. An endogenous peptide signal in Arabidopsis activates components of the innate immune response. Proc Nat Acad Sci USA. 2006;103(26):10098–10103. doi: 10.1073/pnas.0603727103.16785434 PMC 1502512 · doi ↗ · pubmed ↗
- 4Zelman AK, Ma Y, Berkowitz GA. Pathogen elicitor peptide (pep), systemin, and their receptors in tomato: sequence analysis sheds light on standing disagreements about biotic stress signaling components. BMC Plant Biol. 2024;24(1):728. doi: 10.1186/s 12870-024-05403-y.39080569 PMC 11289955 · doi ↗ · pubmed ↗
- 5Shen W, Liu J, Li JF. Type-II metacaspases mediate the processing of plant elicitor peptides in Arabidopsis. Mol Plant. 2019;12(11):1524–1533. doi: 10.1016/j.molp.2019.08.003.31454707 · doi ↗ · pubmed ↗
- 6Yamaguchi Y, Pearce G, Ryan CA. The cell surface leucine-rich repeat receptor for At Pep 1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells. Proc Nat Acad Sci USA. 2006;103(26):10104–10109. doi: 10.1073/pnas.0603729103.16785433 PMC 1502513 · doi ↗ · pubmed ↗
- 7Yamaguchi Y, Huffaker A, Bryan AC, Tax FE, Ryan CA. PEPR 2 is a second receptor for the Pep 1 and Pep 2 peptides and contributes to defense responses in Arabidopsis. Plant Cell. 2010;22(2):508–522. doi: 10.1105/tpc.109.068874.20179141 PMC 2845411 · doi ↗ · pubmed ↗
- 8Poncini L, Wyrsch I, Tendon VD, Vorley T, Boller T, Geldner N, Métraux JP, Lehmann S. In roots of Arabidopsis thaliana, the damage-associated molecular pattern At Pep 1 is a stronger elicitor of immune signalling than flg 22 or the chitin heptamer. P Lo S One. 2017;12(10):1–21. doi: 10.1371/journal.pone.0185808.PMC 562656128973025 · doi ↗ · pubmed ↗