Dynamics of a large multidrug-resistant plasmid encoding New Delhi metallo-β-lactamase-1 and oxacillinase-58 carbapenemases in Acinetobacter baumannii clinical isolates from a tertiary hospital in Malaysia
Nurul Saidah Din, Farahiyah Mohd Rani, Salwani Ismail, Nor Iza A. Rahman, Hong Leong Cheah, Hock Siew Tan, Sadequr Rahman, David W. Cleary, Qasim Ayub, Stuart C. Clarke, Chew Chieng Yeo

TL;DR
This study tracks the spread of a large plasmid in Acinetobacter baumannii that carries genes for antibiotic resistance, including NDM-1 and OXA-58, in a Malaysian hospital over several years.
Contribution
The study reveals the ongoing horizontal transmission and structural variability of a multidrug-resistant plasmid in unrelated A. baumannii isolates.
Findings
The plasmid carrying blaNDM-1 and blaOXA-58 was found in four new MDR A. baumannii isolates from 2018–2020.
Structural variability in the plasmid was driven by insertion sequences and xrs modules, with some isolates lacking blaOXA-58.
An 11.1 kb plasmid with tetracycline resistance genes was identified in one isolate, highlighting the role of xrs modules in AMR dissemination.
Abstract
Acinetobacter baumannii carrying the blaNDM-1 gene, which encodes New Delhi metallo-β-lactamase-1 (NDM-1), exhibits resistance to nearly all β-lactams and is not affected by β-lactamase inhibitors, limiting treatment options. The blaNDM-1 gene is often associated with other antimicrobial resistance (AMR) genes, resulting in multidrug-resistant (MDR) phenotypes. We previously reported a large, circa (ca.) 170 kb plasmid co-harbouring blaNDM-1- and blaOXA-58-encoded carbapenemases in clinical MDR Acinetobacter nosocomialis and A. baumannii isolates from a tertiary hospital in Terengganu, Malaysia, in 2015 and 2016, respectively. In this study, we identified four additional MDR A. baumannii isolates from the same hospital (2018–2020) carrying blaNDM-1 on a similar plasmid. Complete genome sequences were obtained using a hybrid assembly of short-read DNA Nanoball Sequencing(DNBSeq) and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
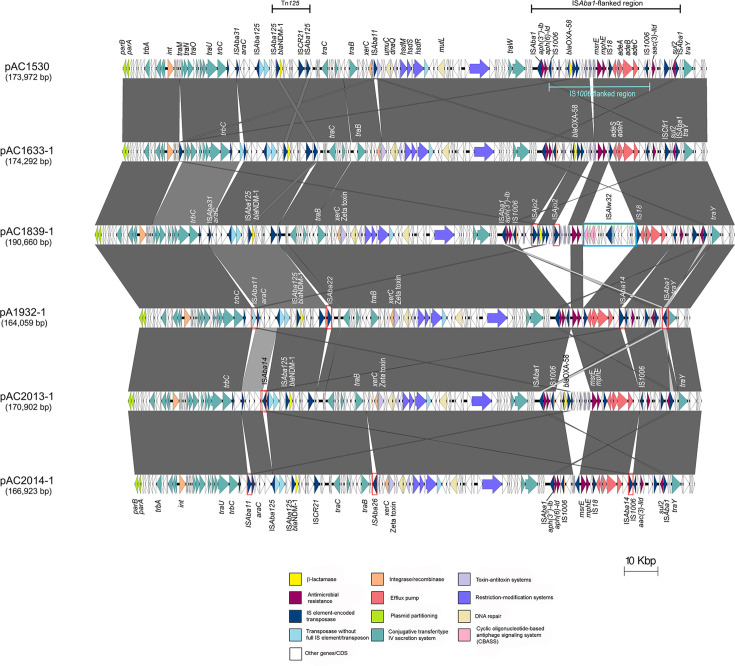 Fig. 1
Fig. 1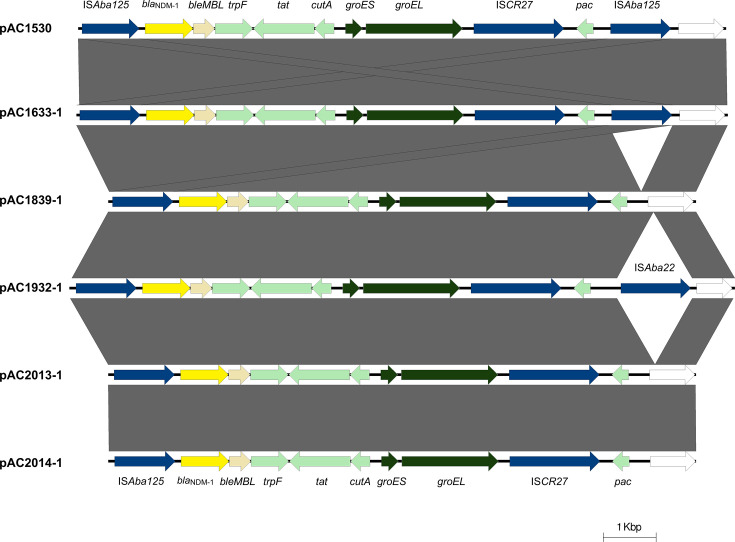 Fig. 2
Fig. 2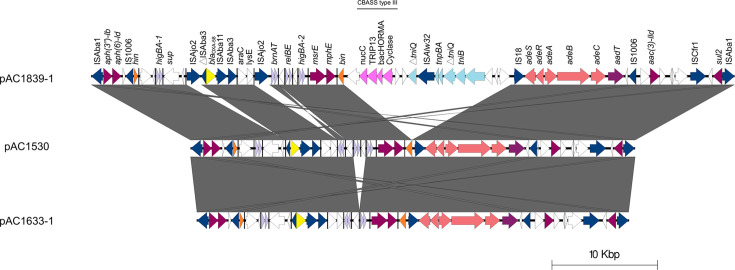 Fig. 3
Fig. 3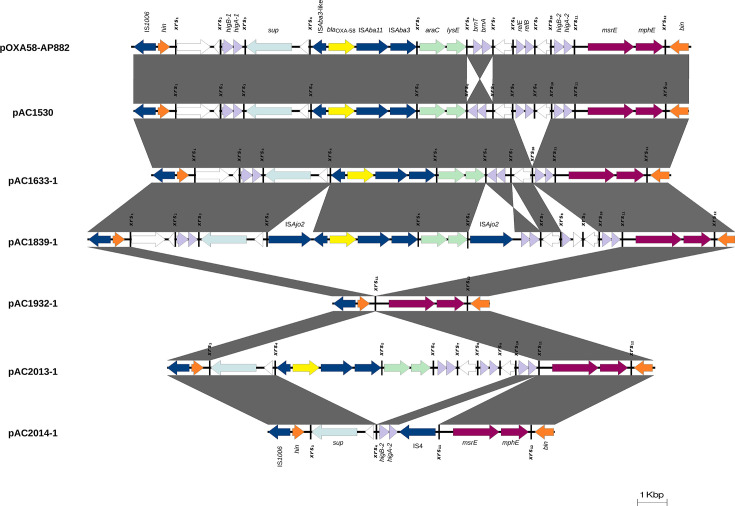 Fig. 4
Fig. 4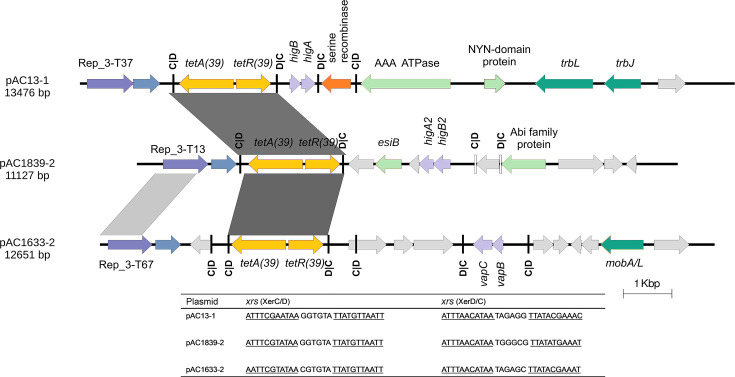 Fig. 5
Fig. 5| Isolate | Year of isolation/ward | Specimen | Oxford ST | Pasteur ST | Chromosome/ plasmid | Size (bp) | G+C content (mol%) | AMR gene | Plasmid |
|---|---|---|---|---|---|---|---|---|---|
| AC1839 | 2 April 2018/Paediatrics | Urine | 3348 | 2575 | Chromosome | 3,830,773 | 39.08 | ||
| pAC1839-1 | 190,660 | 38.41 |
|
| |||||
| pAC1839-2 | 11,127 | 33.62 |
| Rep_3 T13 | |||||
| AC1932 | 7 March 2019/Renal Ward | Blood | 1496 | 1405 | Chromosome | 3,857,525 | 39.13 | ||
| pAC1932-1 | 164,059 | 38.48 |
| ||||||
| pAC1932-2 | 96,136 | 40.26 |
| Rep_3 T7 | |||||
| pAC1932-3 | 35,240 | 39.38 |
|
| |||||
| AC2013 | 16 February 2020/Medical Ward 8C | Blood | 942 | 267 | Chromosome | 3,644,752 | 38.98 | ||
| pAC2013-1 | 170,902 | 38.32 |
| ||||||
| pAC2013-2 | 7145 | 31.99 |
| Rep_3 T5 | |||||
| pAC2013-3 | 5139 | 36.56 |
| Rep_3 T79 | |||||
| AC2014 | 16 February 2020/Medical Ward 8C | Blood | 3393 | 142 | Chromosome | 3,765,139 | 39.06 | ||
| pAC2014-1 | 166,923 | 38.53 |
| ||||||
| pAC2014-2 | 95,694 | 38.42 |
| Rep_3_T21 | |||||
| pAC2014-3 | 40,116 | 36.35 |
| Rep_3 T27 | |||||
| pAC2014-4 | 5304 | 32.77 |
| Rep_3 T13 |
| Antimicrobial class | Antimicrobial compound | AC1633 | AC1839 | AC1932 | AC2013 | AC2014 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC* | Interpretation† | MIC* | Interpretation† | MIC* | Interpretation† | MIC* | Interpretation† | MIC* | Interpretation† | ||
| Aminoglycosides | AMK | ≤4 | S | ≤4 | S | ≤4 | S | 8 | S | ≤4 | S |
| GEN | >8 | R | >8 | R | >8 | R | >8 | R | >8 | R | |
| Carbapenems | ETP | >1 | R | >1 | R | >1 | R | >1 | R | >1 | R |
| IPM | >8 | R | >8 | R | >8 | R | >8 | R | >8 | R | |
| MEM | >32 | R | >32 | R | >32 | R | >32 | R | >32 | R | |
| Cephalosporins | CZO | >16 | R | >16 | R | >16 | R | >16 | R | >16 | R |
| FEP | >16 | R | >16 | R | >16 | R | >16 | R | >16 | R | |
| FOX | >16 | R | >16 | R | >16 | R | >16 | R | >16 | R | |
| CAZ | >16 | R | >16 | R | >16 | R | >16 | R | >16 | R | |
| CRO | >4 | R | >4 | R | >4 | R | >4 | R | >4 | R | |
| CXM | >16 | R | >16 | R | >16 | R | >16 | R | >16 | R | |
| Cephalosporins + β-lactamase Inhibitors | CZA | >8/4 | R | >8/4 | R | >8/4 | R | >8/4 | R | >8/4 | R |
| Fluoroquinolones | CIP | >2 | R | 0.25 | S | ≤0.125 | S | 0.5 | S | ≤0.125 | S |
| LVX | 4 | I | ≤1 | S | ≤1 | S | ≤1 | S | 2 | S | |
| NOR | >8 | I | 4 | S | ≤2 | S | 4 | S | ≤2 | S | |
| Monobactams | ATM | >16 | R | >16 | R | >16 | R | >16 | R | >16 | R |
| Nitrofurans | NIT | >128 | R | >128 | R | >128 | R | >128 | R | >128 | R |
| Penicillins | AMP | >16 | R | >16 | R | >16 | R | >16 | R | >16 | R |
| Penicillins + β-lactamase Inhibitors | SAM | >16/8 | R | >16/8 | R | >16/8 | R | >16/8 | R | >16/8 | R |
| TZP | >64/4 | R | >64/4 | R | >64/4 | R | >64/4 | R | >64/4 | R | |
| Phosphonic acid derivatives | FOS | >128 | R | >128 | R | >128 | R | >128 | R | >128 | R |
| Polymyxins | COL | ≤1 | S | ≤1 | S | ≤1 | S | ≤1 | S | ≤1 | S |
| Sulphonamides | SXT | >2/38 | R | >2/38 | R | >2/38 | R | >2/38 | R | >2/38 | R |
| Tetracyclines | MNO | ≤1 | S | ≤1 | S | ≤1 | S | ≤1 | S | ≤1 | S |
| TGC | ≤1 | S | ≤1 | S | ≤1 | S | ≤1 | S | ≤1 | S | |
- —http://dx.doi.org/10.13039/501100015515 Kementerian Pendidikan Malaysia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Infections and bacterial resistance · Antibiotic Use and Resistance
Data Summary
The authors confirm that all supporting data, code and protocols have been provided within the article or through supplementary data files. The assembled complete genomes in this study have been deposited in the National Center for Biotechnology Information (NCBI) Genomes database under the following accession numbers: Acinetobacter baumannii AC1839, JAQIRC000000000; A. baumannii AC1932, JAQIQK000000000; A. baumannii AC2013, JAQIPS000000000 and A. baumannii AC2014, JAQIPR000000000.
Introduction
For nearly half a century, no new class of antibiotics has been introduced for the treatment of Gram-negative bacteria, including Acinetobacter baumannii infections [12]. Thus, carbapenem-resistant A. baumannii (CRAB) is among the three Gram-negative bacterial pathogens that have received the highest priority classification, and it retained its critical status in the revised 2024 publication of the Bacterial Priority Pathogens List issued by the World Health Organization [3]. Globally, the prevalence of carbapenem resistance among A. baumannii has exceeded 90% in some countries, and hospital-acquired pneumonia and bloodstream infections caused by CRAB have been estimated to account for mortality rates of up to 60% [14]. During the European Antimicrobial Resistance Surveillance Network (EARS-Net) period, which covered 2017–2021, carbapenem resistance in Acinetobacter spp., including CRAB, increased to 74.5% in 2021 compared with 59.2% in 2017 [5], thus highlighting the challenges in combating CRAB infections in the face of increasing resistance [26].
The resistance mechanisms employed by CRAB are generally mediated by intrinsic or acquired mechanisms, or a combination of both. A major resistance pathway is the production of β-lactamase enzymes that hydrolyse carbapenems. In Acinetobacter spp., the most common carbapenem-inactivating carbapenemases are the Class D [oxacillinases (OXAs)] and Class B [metallo-β-lactamase (MBLs)] β-lactamases [7]. The New Delhi metallo-β-lactamase (NDM) has the ability to hydrolyse almost all β-lactam antibiotics, including carbapenems, and is typically located on conjugative plasmids, facilitating the spread of CRAB infections [8]. NDM-1 is not inhibited by β-lactamase inhibitors such as clavulanic acid, sulbactam, tazobactam and avibactam, thus precluding the use of β-lactam/β-lactamase inhibitor combination therapy [910]. In CRAB and other Gram-negative pathogens, the blaNDM-1 gene is often observed within a Tn125 composite transposon that is flanked by two copies of the ISAba125 insertion sequences (ISs) [1112] The horizontal transmission of Tn125 carrying the blaNDM-1 gene from a CRAB strain to a carbapenem-susceptible A. baumannii strain, which was hypothesized to occur by transduction, has been described [13]. More often, Acinetobacter spp. harbouring blaNDM-1 are also multidrug resistant (MDR) due to its association with other antimicrobial resistance (AMR) genes [1415].
Since its initial spread among the Enterobacterales, blaNDM-1 was quickly identified in Acinetobacter spp., which are considered a key reservoir for the gene [1617]. NDM-producing Acinetobacter spp. have been reported from nearly all continents, often causing difficult-to-treat nosocomial outbreaks [18]. This rapid globalization is driven by the gene’s association with mobile genetic elements, particularly the Tn125 transposon, which is highly prevalent in Acinetobacter genomes [17]. The phenomenon is not limited to A. baumannii; the blaNDM-1 gene has been identified in a wide range of Acinetobacter species, including Acinetobacter nosocomialis, Acinetobacter pittii and Acinetobacter junnii, and often on large transmissible plasmids, highlighting its extensive interspecies mobility within the genus [816].
This global challenge is reflected at the local level. In Malaysia, NDM-producing Acinetobacter have been documented but without any in-depth characterization of their genetic environment [19]. We had reported two blaNDM-1-producing, carbapenem-resistant MDR Acinetobacter spp. isolates from the main tertiary hospital in Terengganu, on the east coast of Peninsular Malaysia: A. nosocomialis AC1530, which was isolated in 2015, and A. baumannii AC1633, isolated in 2016 [20]. Both isolates harboured an almost identical, ca. 170 kb plasmid (designated pAC1530 and pAC1633-1, respectively) that encoded the blaNDM-1 gene in Tn125 and the blaOXA-58-encoded Class D carbapenemase located within a mobile Xer recombination site (xrs) or plasmid-dif (pdif) module. These modules form a novel class of discrete mobile elements that mobilize via Xer-mediated site-specific recombination through a cointegration/resolution mechanism [2122]. Since the discovery of the dif module, which carried blaOXA-24 in the A. baumannii plasmid pABVA01 [23], these elements have been identified in numerous Acinetobacter plasmids. They are characterized by flanking XerC/XerD recombination sites and were initially designated pdif (as well as Re27 and pXerC/XerD) to differentiate them from chromosomal dif sites [2425]. However, these elements have been recently redesignated xrs for ‘Xer recombination sites’ [18] since ‘dif’ is an acronym for ‘deletion-induced filamentation site’, which is a phenotype not associated with plasmid xrs sites.
Plasmids pAC1530 and pAC1633-1 also co-located numerous other AMR genes [20]. The discovery of this plasmid in two different species of Acinetobacter from the same hospital strongly suggested horizontal transmission, but conventional conjugative assays failed to detect such transfer [20]. Further surveillance from 2018 to 2020 identified four additional blaNDM-1-carrying CRAB isolates, two of which also co-harboured blaOXA-58 [26]. Since these isolates were initially sequenced using only short-read technology, determining the complete structure of the plasmids and confirming their relationship to the original pAC1530/pAC1633-1 was challenging.
In this study, the genomes of these four recent blaNDM-1-carrying A. baumannii isolates were subjected to long-read Nanopore sequencing and hybrid assembly with previous short-read data. The aim was to obtain their complete genome sequences, fully resolve the structures of their large blaNDM-1 plasmids and investigate the structural dynamics of this plasmid across different Acinetobacter genomes over a 6-year period (2015–2020).
Methodology
Bacterial collection and antibiotic susceptibility tests
Four clinical isolates of blaNDM-1-positive A. baumannii investigated in this study were collected between 2018 and 2020 at Hospital Sultanah Nur Zahirah (HSNZ), the main tertiary hospital in Terengganu, Malaysia. These isolates were among the 126 Terengganu A. baumannii isolates whose genomes were recently published [26].
Antimicrobial susceptibility testing was performed using the BD Phoenix™ M50 System (Becton, Dickinson and Company, USA) with the BD Phoenix NMIC-500 panel (Becton, Dickinson and Company, USA). The susceptibility criteria were determined following the latest Clinical and Laboratory Standards Institute guidelines (M100-S35) [27]. CRAB isolates exhibited resistance to imipenem and meropenem. The NMIC-500 panel that was used also detected and classified carbapenemase-producing organisms (CPOs) [28].
Whole-genome sequencing and sequence analysis
Bacterial genomes were extracted using the GenJET Genomic DNA Purification kit (Thermo Scientific, MA, USA) for short-read sequencing and a conventional phenol–chloroform protocol [29] for extracting high-molecular-weight genomic DNA for long-read sequencing. The DNA was subjected to whole-genome sequencing on the DNBSEQ (MGI Tech Co. Ltd., Shenzhen, China) and the Nanopore MinION (Oxford Nanopore Technologies, Oxford, UK) sequencing platforms. Paired-end DNA libraries were constructed with an average insert size of 100 bp for short-read sequencing using the DNBSEQ-G400 by a commercial service provider (BGI, Shenzhen, China). For long-read sequencing, DNA libraries were constructed using the Rapid Barcoding kit (SQK- RBK114.24) and sequenced using R10.4 flow cells on a MinION device. Basecalling was performed using MinKNOW in super-accuracy basecalling mode. FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) was used for quality control of raw reads. A hybrid assembly was then performed using Unicycler v0.5.1 (https://github.com/rrwick/Unicycler) [30], incorporating both the short paired-end reads and the long reads. The assembly quality was checked using QUAST v5.3.0 (https://github.com/ablab/quast).
Genome annotation was performed using PROKKA (https://github.com/tseemann/prokka), and the generated General Feature Format (GFF) file was then used in Roary (https://github.com/tseemann/prokka) to obtain the core-genome alignment using the criteria of 95% sequence identities and presence in 99% of genomes [2031]. The list of A. baumannii genomes used to construct the core-genome alignment is provided in Table S1, available in the online Supplementary Material. VeryFastTree (https://github.com/citiususc/veryfasttree) was used to construct the maximum-likelihood phylogenetic tree with 1,000 bootstrap replicates under the generalized time-reversible model [32]. The phylogenetic tree was then visualized using iTOL v6 (https://itol.embl.de/) [33]. Multilocus sequence typing (MLST) profiles were analysed using the MLST Oxford and Pasteur scheme via PubMLST (https://pubmlst.org/organisms/acinetobacter-baumannii) [34]. The capsular (K) and outer core (OC) loci were identified using Kaptive (https://kaptive-web.erc.monash.edu/) [35] with default parameters. ResFinder (http://genepi.food.dtu.dk/resfinder) [36] and CARD (https://card.mcmaster.ca/) [37] were used to screen for the presence of antibiotic resistance genes. IS elements were determined using ISfinder (https://isfinder.biotoul.fr/) [38], while the presence of xrs (or pdif) sites was determined by a combination of pdif finder (https://github.com/mjshao06/pdifFinder) [39] and manual blastn screening using known XerC/XerD and XerD/XerC sites in the pAC1530 and pAC1633-1 plasmids. Analysis of SNPs in the core regions of the plasmids was performed using snp-dists v0.8.2 (https://github.com/tseeman/snp-dists), with pAC1530 as the reference plasmid. The SNP-based phylogenetic tree of the blaNDM-1-encoded plasmids was generated using RAxML-NG (https://github.com/amkozlov/raxml-ng) and visualized with iTOL v6. Phage defence genes were identified using DefenseFinder (https://defensefinder.mdmlab.fr/) [40]. Comparative genome analysis was carried out using the National Center for Biotechnology Information (NCBI) blast (https://blast.ncbi.nlm.nih.gov/Blast.cgi) between the current plasmids and the reference genomes, and these were then visualized using EasyFig v2.2.5 (https://mjsull.github.io/Easyfig/) [41].
Results
Characteristics of CRAB clinical isolates from HSNZ harbouring blaNDM-1
Genome analyses of the 126 A. baumannii clinical isolates obtained over a 10-year period (2011–2020) from HSNZ led to the identification of five CRAB isolates harbouring the blaNDM-1 gene (AC1633, AC1839, AC1932, AC2013 and AC2014) [26]. The complete genome sequences of A. baumannii AC1633, which was isolated in 2016, had previously been determined together with AC1530, a blaNDM-1-carrying, carbapenem-resistant A. nosocomialis that was obtained from the same hospital in 2015 [20]. These two isolates were the earliest Acinetobacter spp. in our collection found to harbour an almost identical, ca. 170 kb plasmid (designated pAC1633-1 in A. baumannii AC1633; accession no. CP059301, and pAC1530 in A. nosocomialis AC1530; accession no. CP045561) that encodes the blaNDM-1 gene along with the blaOXA-58 carbapenemase [20]. Both plasmid sequences were used as reference sequences for this study.
Of the four remaining blaNDM-1-positive A. baumannii isolates, A. baumannii AC1839 was recovered from a urine culture of a 1-year-old male patient in the paediatric unit of HSNZ in April 2018, while A. baumannii AC1932 was isolated from a blood culture of a 50-year-old female patient in the renal care unit in March 2019. A. baumannii AC2013 and A. baumannii AC2014 were both obtained from blood cultures of two different female patients (aged 69 and 45, respectively) on the same day (16 February 2020) in the same medical ward, 8C (Table 1). These four A. baumannii isolates were phenotypically resistant to all β-lactam antibiotics tested, including cephalosporins and carbapenems [minimum inhibitory concentrations (MICs) of imipenem ≥8 µg ml^−1^, meropenem ≥32 µg ml^−1^ and ertapenem ≥1 µg ml^−1^] (Table 2), but were susceptible to ciprofloxacin, levofloxacin, minocycline, tigecycline and colistin. Among the aminoglycosides, all four isolates showed resistance to gentamicin but were susceptible to amikacin (Table 2). However, only A. baumannii AC1839 was resistant to tetracycline among the four isolates, as shown by previous disc diffusion assays [26]. All four isolates were also resistant to trimethoprim–sulphamethoxazole (Table 2). Based on the BD Phoenix™ M50 System, the four A. baumannii isolates were determined to be Class B CPOs.
Genome characteristics of the blaNDM-1-positive A. baumannii isolates
Hybrid assembly of DNBSeq paired-end short-read and Nanopore long-read sequence data led to the generation of the complete genome sequences of the four blaNDM-1-positive A. baumannii isolates. Each isolate comprised a circular chromosome of ca. 3.64–3.85 Mbp with highly similar mol% G+C content and plasmids of varying sizes (Table 1). MLST analysis of the blaNDM-1-positive A. baumannii genomes revealed that they were non-clonal and belonged to four distinct Oxford and Pasteur Sequence Types (STs) [26] (Table 1). A. baumannii AC1839 belonged to novel STs for both the Oxford (assigned by PubMLST curators as ST3348: gltA-1, gyrB-1, gdhB-80, recA-6, cpn60-42, gpi-110, rpoD-26) and Pasteur schemes (assigned by PubMLST as ST2575: cpn60-56, fusA-100, gtlA-2, pyrG-2, recA-9, rplB-4, rpoB-5) (Table 1). AC2014 belonged to a pre-existing ST142_Pasteur_ but a novel Oxford ST (assigned as ST3393: gltA-36, gyrB-12, gdhB-62, recA-31, cpn60-22, gpi-534, rpoD-4). A. baumannii AC2013, which was isolated on the same day and from the same ward, albeit from different patients, as AC2014, was ST942_Oxford_ and ST267_Pasteur_ (Table 1). Core genome phylogenetic analysis of Malaysian A. baumannii isolates, alongside several reference genomes, clearly demonstrated that the blaNDM-1-carrying isolates were genetically distinct and unrelated (Fig. S1).
These A. baumannii isolates harboured a large, blaNDM-1-encoding plasmid ranging in size between 164,059 and 190,660 bp, with two of these plasmids (namely pAC1839-1 and pAC2013-1 in A. baumannii AC1839 and AC2013, respectively) harbouring both blaNDM-1 and blaOXA-58 (Table 1). None of these four large plasmids carried any known plasmid replication initiator (or rep) gene that is found in the Acinetobacter Plasmid Typing database [4243], similar to what was observed for pAC1633-1 isolated from A. baumannii AC1633 and pAC1530 from A. nosocomialis AC1530 [20]. This is suggestive of a novel rep gene encoded by these plasmids that has yet to be characterized. These plasmids harbouring blaNDM-1 currently have no homologous counterparts in the databases and are, therefore, unique to Acinetobacter isolates from Malaysia. Of note, we had postulated that pAC1530 and pAC1633-1 were derived from an IS1006-mediated cointegration of two plasmids – one carrying blaNDM-1 and the other encoding blaOXA-58 – due to the high sequence identity with the two plasmids found in A. pittii AP882, a hospital isolate from the state of Perak, Malaysia, in 2014 [2044]. A blastn comparison of recently reported large Acinetobacter plasmids harbouring blaNDM-1 and blaOXA-58 [15] showed sequence similarities only at the Tn125 region that contained blaNDM-1 and in the sequences surrounding the blaOXA-58 region (Fig. S2), thus reinforcing the uniqueness of the plasmid backbone harbouring these two carbapenemase genes from Malaysia. Likewise, blastn with the NCBI non-redundant database did not result in any significant hits beyond the Tn125-blaNDM-1 and blaOXA-58 regions.
The four blaNDM-1-positive A. baumannii isolates also harboured an additional one to three other plasmids that ranged in size from 5,139 to 96,136 bp, most of which encoded plasmid Rep proteins of the Rep_3 family (Table 1). Only the 11,127 bp pAC1839-2 from A. baumannii AC1839 harboured the tetA(39) tetracycline-resistance gene, while the other plasmids did not encode any AMR genes. An analysis of the pAC1839-2 structure is presented in Section 3.7.
Overview of the large blaNDM-1-encoding plasmids in the four A. baumannii isolates
Comparison of the four large blaNDM-1-encoding plasmids identified in this study with two previously described Acinetobacter plasmids harbouring blaNDM-1 (i.e. pAC1530 and pAC1633-1) [20] revealed several structural differences (Fig. 1). Notably, a 29,670 bp region flanked by IS1006 showed extensive variability. This region contains multiple xrs recombination sites and AMR genes, such as blaOXA-58 and msrE–mphE, and is inserted within a 14,750 bp composite transposon flanked by ISAba1, designated Tn6948. Tn6948 harbours additional AMR genes, including sul2 and the aminoglycoside resistance genes aac(3)-IId, aph(3'')-Ib and aph(6)-Id [20]. The IS1006 element was hypothesized to be the point of insertion (possibly through transposition or recombination) of a large segment from the blaOXA-58-encoding, 36 kb plasmid pOXA-58_AP882 (accession no. CP014479) into the 147 kb blaNDM-1-encoding plasmid pNDM-1_AP882 (accession no. CP014478). These plasmids were previously identified in an A. pittii clinical isolate from Perak, Malaysia, in 2014 [44], and were postulated to have given rise to the pAC1530/pAC1633-1 progenitor plasmid, which subsequently spread to Acinetobacter isolates in Terengganu [20].
Comparative linear maps of the blaNDM-1-encoded plasmids identified in this study, along with plasmid pAC1530 from A. nosocomialis AC1530 (accession no. CP045561) and pAC1633-1 from A. baumannii AC1633 (accession no. CP059301), which were identified and presented previously [20]. Arrows represent genes/open reading frames that are coloured according to the legend provided. Grey-shaded areas between each linear plasmid map indicate regions that shared >97% nucleotide sequence identities. Red boxes indicate IS elements that were not shared among all plasmids, while the sky blue-coloured box outlined in the pAC1839-1 linear map referred to the ca. 15 kb putative transposon that contained ISAlw32 and the type III CBASS phage defence system (see Section 3.5).
Among the plasmids harbouring blaNDM-1, pAC1839-1 was the largest, at 190,660 bp. A distinctive feature of this plasmid is the insertion of a unique 15,434 bp fragment between the bin resolvase gene and IS18 within the IS1006-flanked region. This fragment appears to represent a novel transposon-like element exclusive to pAC1839-1 and is described in detail in Section 3.5. Additionally, pAC1839-1 harbours two copies of ISAjo2 near the blaOXA-58 gene, which are absent in the other plasmids (Fig. 1). In the remaining three blaNDM-1-encoding plasmids, most of the structural variations were also confined to the IS1006-flanked region, which is rich in xrs recombination sites. Notably, the blaOXA-58 gene and several surrounding genes in xrs modules are absent in both pAC1932-1 and pAC2014-1 (Fig. 1). Further details regarding the insertions and deletions involving xrs modules are presented in Section 3.6.
Besides the variations within the IS1006-flanked xrs-rich regions, other structural changes include the insertions of various IS elements. Both pAC1932-1 and pAC2014-1 have the 1,282 bp ISAba14 inserted upstream of the IS1006 copy, which is located one gene away from the aac(3)-IId aminoglycoside resistance gene (Fig. 1). Both plasmids also have the 1,101 bp ISAba11 inserted adjacent to the ISAba31 found upstream of the araC gene; in pAC1633-1, there is a partial IS element with an IS4 family transposase inserted near the same location, and this insertion led to the truncation of ISAba31 (Fig. 1) [20]. An ISAba1 is found upstream of the traY gene in pAC1932-1, but this is not seen in pAC2014-1 or pAC2013-1. On the other hand, pAC2014-1 carries the insertion of ISAba26 (1,318 bp, an element of the IS256 family) near the traB gene, while pAC2013-1 contains ISAba14 (1,282 bp; IS3 family) next to ISAba125 upstream of the Tn125 transposon, which carries the blaNDM-1 gene (?Fig. 1). ISAba26 and ISAba14 are absent in the other blaNDM-1-encoded plasmids.
A SNP analysis was conducted on the core regions of the six blaNDM-1-encoded plasmids, with pAC1530 as the reference plasmid, revealing low SNP counts of between 2 and 200 (Table S2), thus inferring that the plasmids are highly conserved. Interestingly, pAC1633-1 had only two SNP differences compared with pAC1530, indicating that these two plasmids are nearly identical and underlining the suggestion that the plasmid was horizontally transmitted between A. nosocomialis AC1530 and A. baumannii AC1633 [20]. A phylogenetic tree generated from the pairwise core SNP differences of the plasmids (Fig. S3) supported the close relatedness of the six plasmids, which shared a recent common ancestor. pAC1932-1 (without blaOXA-58) was the most divergent, with 200 SNP differences when compared with pAC1530, and between 121 and 129 SNP differences when compared with the other three plasmids (including the other plasmid that was absent for blaOXA-58, pAC2014-1). Plasmid copy numbers were estimated by mapping the respective plasmid short reads to the assembled complete genomes as the reference and analysing the coverage depth relative to the chromosome, leading to average values of between 1.4 and 1.7 (Table S3).
The blaNDM-1 gene is found in variant Tn125 structures
The most common vehicle for the dissemination of the blaNDM-1 carbapenemase gene in Acinetobacter spp. is the composite transposon Tn125, which is usually flanked by ISAba125 [141845] and was found in both pAC1530 and pAC1633-1 [20]. However, in the four NDM-1-positive A. baumannii isolates, Tn125 had lost the ISAba125 located downstream of blaNDM-1. In pAC1932-1, this copy of ISAba125 was replaced by ISAba22 (Fig. 2). Variants of Tn125 have been previously reported with the loss of one copy of ISAba125 in a ca. 46 kb plasmid pNDM-BJ02 from a clinical isolate of Acinetobacter lwoffii from China [46] and in a ca. 64 kb plasmid in A. baumannii DT01139, isolated from a blood culture of a neonate in Tanzania [47]. In the case of the pAP-D499 plasmid of an A. pittii isolate from China, the downstream ISAba125 element was replaced by ISAba11 [48], whereas in A. baumannii AB-NDM-1, isolated in Spain, Tn125 was located on the chromosome and was flanked by ISAba125 and ISAba14 [49].
Comparative linear map of the Tn125 structure found in the blaNDM-1-encoded plasmids pAC1530 and pAC1633-1 identified in the genomes of A. nosocomialis AC1530 and A. baumannii AC1633, respectively [20]), and in the plasmids identified from A. baumannii AC1839, AC1932, AC2013 and AC2014 in this study. Grey-shaded areas between the linear plasmid maps indicate regions of 100% nucleotide sequence identities. IS element-encoded transposases are indicated in dark blue arrows; the blaNDM-1 gene is depicted as a yellow arrow, with its downstream bleMBL bleomycin-resistance gene as a beige arrow. Genes encoding the GroES and GroEL chaperonin proteins are shown in dark green arrows, while light green arrows indicate genes with known functions, and white arrows indicate reading frames encoding hypothetical proteins.
A novel 15 kb transposon-like element with a phage defence system was inserted into pAC1839-1
In pAC1839-1, there is an additional 15,434 bp fragment between the bin resolvase and IS18 within an xrs-rich 29,670 bp region flanked by IS1006 (Fig. 3). Numerous transposases and transposase-like sequences were found within this 15.4 kb region. Three transposase genes identified by ISfinder belong to a 2,512 bp ISAlw32 element of the IS66 family, which is flanked by 16 bp inverted repeats. ISAlw32 was first described in A. lwoffii M2a, isolated from honey [50]. In pAC1839-1, ISAlw32 was inserted into a tniQ-encoded putative transposase, and this insertion led to an 8 bp direct repeat of the target site (5′-ATTTCTTT-3′). An intriguing set of four genes belonging to the cyclic oligonucleotide-based antiphage signalling system (CBASS) was identified within this 15.4 kb, transposase-rich region in pAC1839-1 (Fig. 3). CBASS is a phage defence system in bacteria and archaea that utilizes cGAS/DncV-like nucleotidyltransferase (CD-NTase, also known as cyclase) proteins to sense phage infection. It is activated to synthesize a cyclic nucleotide immune signal, which then binds to Cap (CD-NTase-associated protein) effectors that induce host cell death to inhibit phage propagation, as reviewed in [51]. CBASS systems are highly diverse, with four types that have been described so far; all four types have the CD-NTase and Cap effector genes as their core, with genes encoding different additional Cap proteins that regulate the CD-NTase function as ancillary proteins [5152]. The CBASS system found in pAC1839-1 belongs to type III, as it includes ancillary genes that encode Cap proteins with HORMA and TRIP13 domains [52], in addition to the core CD-NTase and Cap effector genes (Fig. 3). The pAC1839-1-encoded type III CBASS system is not found in the other four A. baumannii genomes analysed in this study (i.e. AC1633, AC1932, AC2013 and AC2014). Nevertheless, a blastn search of the other 122 A. baumannii genomes from the same hospital, as published in [26], showed a hit with A. baumannii AC2010 (contig_39; accession no. JAQIPT0000000039.1) and only for the four genes that made up the type III CBASS (at 80% sequence similarity). The ISAlw32 sequence was identified across three contigs (contig_70, contig_77 and contig_80), and no other sequences from the 15.4 kb region of pAC1839-1 could be found in the AC2010 genome. We are also unable to ascertain whether the type III CBASS system in AC2010 is plasmid or chromosomally encoded due to the low contiguity of the short-read assembly. This suggests the rarity of the type III CBASS system in our A. baumannii collection, and in AC1839, it was likely acquired along with the transposon-like element that had inserted into pAC1839-1. Unfortunately, very little is known about CBASS and most other phage defence systems in Acinetobacter spp. [53]. Although a very comprehensive in silico survey of phage defence systems and their hotspots in Acinetobacter spp. was recently published [54], their functional characterization awaits further investigations.
Comparative linear genetic map of the ISAba1-flanked regions of the blaNDM-1-encoded plasmids pAC1839-1 with pAC1530 and pAC1633-1, showing the putative 15,434 kb transposon inserted between the bin recombinase and IS18 in pAC1839-1. AMR genes are indicated in maroon-coloured arrows, except blaOXA-58, which is depicted as a yellow arrow. Efflux-mediated genes are shown as salmon-coloured arrows; IS-encoded transposases are indicated as dark blue arrows, while the transposases found within the 15,434 bp transposon are shown as light blue arrows. Toxin-antitoxin genes are shown as light purple arrows, while genes that made up the type III CBASS system are depicted as pink arrows and labelled. White arrows represent hypothetical genes. Vertical bars on the linear maps indicate xrs sites. The extent of regions with >99% nucleotide sequence identities is indicated in the grey-shaded area between each linear map.
Rearrangements surrounding the blaOXA-58 region likely driven by Xer recombination
In addition to the blaNDM-1 carbapenemase gene, plasmids pAC1530 and pAC1633-1 also co-harboured the blaOXA-58 class D carbapenemase gene, and a hallmark of this gene is its location within a xrs module flanked by XerC–XerD recombination sites [20]. The region surrounding blaOXA-58 is rich in XerC–XerD sites (or xrs sites) Table 3 with xrs modules harbouring toxin–antitoxin systems, the msrE–mphE macrolide resistance genes, and others found within a 29 kb region surrounding blaOXA-58 that is flanked by IS1006 [20]. When the plasmids from this study were compared with pAC1530 and pAC1633-1, several rearrangements were observed within this region.
Only two of the four plasmids characterized in this study (namely pAC1839-1 and pAC2013-1) were found to harbour both the blaNDM-1 and blaOXA-58 genes; in the other two plasmids (i.e. pAC1932-1 and pAC2014-1), the blaOXA-58 gene was absent. The xrs sites are numbered here according to their order in pAC1530, which had two additional xrs sites compared with pAC1633-1 [20]. In pAC1932-1, there is an absence of an 11.5 kb fragment spanning the xrs1 site downstream of the hin recombinase to the higBA-2 toxin–antitoxin genes upstream of the xrs11 site, leaving only the msrE–mphE macrolide resistance module flanked by xrs11 and xrs12 (Fig. 4). A smaller deletion was observed in pAC2014-1, which involved a 7.7 kb fragment spanning the xrs4 site downstream of the sup gene to the hypothetical gene that was flanked by the xrs9 and xrs10 sites in pAC1530 (Fig. 4). In addition, there was an insertion of IS4 immediately downstream of the higBA-2 toxin–antitoxin genes located in between the xrs4 and xrs11 sites of pAC2014-1 (Fig. 4). The absence of blaOXA-58 in A. baumannii AC1932 and AC2014 does not appear to affect their resistance to carbapenems, as both isolates showed similar MIC levels for meropenem (>32 µg ml^−1^), imipenem (>8 µg ml^−1^) and ertapenem (>1 µg ml^−1^) compared with their blaOXA-58-positive counterparts (i.e. AC1633, AC1839 and AC2013) (Table 2). This suggests that in these A. baumannii isolates, carbapenem resistance was primarily attributed to the blaNDM-1 gene*.*
Comparative linear map of the xrs-rich regions surrounding blaoxa-58 in pAC1839-1 and pAC2013-1, with reference plasmids pAC1530 from A. nosocomialis AC1530, pAC1633-1 from A. baumannii AC1633 [20] and pOXA58-AP882 from A. pittii AP882 [44]. AMR genes are depicted as maroon arrows, except blaOXA-58, which is shown as a yellow arrow. IS element-encoded transposases are shown as dark blue arrows and labelled accordingly. Light purple-coloured arrows depict toxin–antitoxin systems, while orange arrows indicate putative recombinases. Light green and light blue arrows indicate genes with known functions, and white arrows indicate reading frames encoding hypothetical proteins. xrs sites are shown as vertical bars and labelled as listed in Table S4, which details the respective xrs nucleotide sequences.
When comparing pAC2013-1 with pAC1530, a 2.2 kb fragment that spans the xrs1 to xrs3 sites and covers the higBA-1 toxin–antitoxin genes was absent. However, the remaining regions that span sup, blaOXA-58, until the higBA-2 and msrE–mphE genes remain intact (Fig. 4). In the case of pAC1839-1, insertions of two copies of the 1,482 bp ISAjo2 were observed: one copy is located immediately after the xrs4 site, upstream of the partial ISAba3 element found next to blaOXA-58. This insertion of ISAjo2 led to a 6 bp AGAGAG duplication of the insertion site. Another copy of ISAjo2 is found after the xrs6 site (its insertion resulting in a 5 bp GCAGC target site duplication) and is located upstream of the brnTA toxin–antitoxin xrs module, which is in inverted orientation compared with its counterparts in pAC1530 and pAC1633-1 (Fig. 4). One of the differences between pAC1530 and pAC1633-1 is the absence of the relBE toxin–antitoxin genes, which were flanked by the xrs8 and xrs9 sites, and a hypothetical gene flanked by xrs9 and xrs10 in pAC1633-1 [20]; these genes are, however, present in pAC1839-1 as well as pAC2013-1 (Fig. 4). The organization and orientation of the xrs sites are almost identical in all the plasmids analysed; comparative analysis of the 28 bp core regions of the xrs sites showed only up to two mismatched nucleotides within the XerC and/or XerD binding sites (highlighted in yellow in the sequences shown in Table S4).
The tetA(39) tetracycline resistance gene in pAC1839-2 is also located on a mobile xrs module
A. baumannii AC1839 also harboured the tetA(39)-tetR(39) gene pair in a smaller plasmid designated pAC1839-2 (11,127 bp), which is classified as an R3-T13-type plasmid based on the Acinetobacter plasmid classification scheme [42]. Like the blaOXA-58 gene in pAC1839-1, the tetA(39)-tetR(39) gene pair is located within a xrs module (Fig. 5). The 2,001 bp tetA(39)-tetR(39) xrs module has been found in several Acinetobacter plasmids [24], and we have previously reported its presence in a 13,476 bp plasmid of the R3-T37 group designated pAC13-1 in three A. nosocomialis isolates from HSNZ in 2011 [55] and a 12,651 bp plasmid of the R3-T67 group designated pAC1633-2 in A. baumannii AC1633, isolated from the same hospital in 2016 [20]. Comparison of these three plasmids showed sequence similarity only in the tetA(39)-tetR(39) genes within the xrs module (Fig. 5), inferring the mobility of the tetracycline resistance xrs module among different strains of Acinetobacter spp. in HSNZ since 2011. The xrs sequences flanking the tetA(39)-tetR(39) genes were also highly similar (Fig. 5). The tetA(39) gene conferred resistance to tetracycline but not to doxycycline or minocycline [2455], and this was also observed in A. baumannii AC1839 (Table 2) [26].
Linear maps of the plasmids harbouring the tetA(39)-tetR(39) xrs module from Acinetobacter spp. clinical isolates from HSNZ. The plasmids shown are pAC13-1 from A. nosocomialis AC13, which was isolated in 2011 [55]; pAC1633-2 from A. baumannii AC1633, isolated in 2016 [20] and pAC1839-2 from A. baumannii AC1839, isolated in 2018 (this study). The plasmid replicase gene is indicated in purple arrows, while its downstream gene, designated orfX, is shown in dark blue arrows. The tetA(39)-tetR(39) tetracycline resistance genes are shown as yellow arrows. Toxin–antitoxin genes are depicted in light purple arrows; plasmid mobilization-related genes are shown in green arrows; and light green arrows indicate genes with known functions or domains. NYN-domains are protein domains typified by the eukaryotic Nedd4-binding protein 1 and the bacterial YacP-like proteins (Nedd4-BP1, YacP nucleases). Orange arrows depict putative recombinases. The xrs sites are indicated as thick vertical lines, labelled as C|D for XerC–XerD and D|C for XerD–XerC recognition sites. The nucleotide sequence for the xrs sites flanking the tetA(39)-tetR(39) genes is shown below the linear map.
Discussion
Ever since the first report of NDM-1 in Klebsiella pneumoniae 05–506, isolated in New Delhi, India, in 2008 [56], NDM-1 and its many variants have been reported in various species and have spread worldwide [57]. A. baumannii clinical isolates harbouring the blaNDM-1 were first reported in India [58] and China [59] in 2010. Although OXA-23 is the prevalent acquired carbapenemase in clinical A. baumannii isolates in most parts of the world, there have been increasing reports of A. baumannii producing NDM-1 in recent years. In these cases, the blaNDM-1 gene is often found within Tn125 on large, transmissible plasmids that serve as vehicles for its dissemination [81560]. We previously reported a ca. 170 kb plasmid harbouring blaNDM-1 along with blaOXA-58 and several other AMR genes in A. nosocomialis AC1530 and A. baumannii AC1633 from HSNZ in Terengganu, Malaysia [20]. Subsequent genome sequencing of other A. baumannii isolates from the same hospital revealed four additional A. baumannii isolates that carry blaNDM-1. However, the data were obtained from short-read assemblies [26], which made it challenging to piece together the complete genetic structure of the plasmid(s) that harboured the gene. In this study, hybrid assembly with long-read Nanopore sequencing data enabled us to resolve the complete structure of these plasmids, allowing the dynamics of the blaNDM-1-encoded plasmid to be investigated as it was transmitted within the same hospital between 2015 and 2020.
Several large, conjugative Acinetobacter plasmids carrying blaNDM-1 have been characterized, and recent comparative studies have highlighted their remarkable diversity [815]. Tang et al. [8] identified a 45,911 bp plasmid in A. junnii YR7, designated pNDM-YR7, which harbours blaNDM-1 within a truncated Tn125 that lacked the downstream ISAba125. They also identified 30 additional Acinetobacter plasmids that shared a similar conjugative transfer region and origin of transfer (oriT) with pNDM-YR7. Notably, none of these plasmids contained an identifiable rep gene, an observation similar to the blaNDM-1 plasmids described in this study. In addition, Tang et al. [8] identified 32 other blaNDM-1-harbouring plasmids that differ from pNDM-YR7 in various Acinetobacter spp. Among these, 19 of which encoded a replicase gene of the Rep_3 family. Rodrigues et al. [15] described a very large, ca. 340 kb plasmid, pCCBH31258, which harboured blaNDM-1 within a complete Tn125 structure, as well as blaOXA-58 embedded in an xrs module that differs from the module found in the Malaysian plasmids. In pCCBH31258, the blaOXA-58 xrs module comprises ISAba3-blaOXA-58-ΔISAba3-ISApi2 [15], whereas in pAC1839-1 and the other Malaysian NDM-1 plasmids, the xrs structure comprises ΔISAba3-blaOXA-58-ISAba11-ISAba3. Rodrigues et al. [15] also identified a rep gene in pCCBH31258, which reportedly does not conform to the current Acinetobacter plasmid classification scheme [4243]. Four other A. baumannii isolates from the same hospital in Brazil harbour plasmids similar to pCCBH31258, but their complete structure could not be resolved due to fragmented assembly from Illumina short-read data [15]. Intriguingly, five other NDM-1-producing A. baumannii isolates sequenced in the same study were found to possess plasmids with a distinct genetic background from pCCBH31258 and other known Acinetobacter NDM-1 plasmids [15]. These findings underscored the considerable genetic diversity of plasmids that harbour the blaNDM-1 gene in Acinetobacter spp.
The comparative analysis of the large blaNDM-1-encoded plasmids in this study highlights their structural fluidity, mediated by various mobile elements, as they move between different strains. Notably, all five A. baumannii isolates carrying this plasmid were clonally unrelated, strongly suggesting horizontal transfer of the plasmid, which was initially identified in A. nosocomialis AC1530. Genes associated with conjugative transfer were identified in plasmids pAC1530 and pAC1633-1, sharing 50–70% amino acid sequence identity with the corresponding conjugative transfer proteins of plasmid pA297-3 from A. baumannii A297 [2061]. The same conjugative transfer genes were also found in the four large plasmids analysed in this study. Although functional validation through plasmid transfer experiments (e.g. conjugation or transformation into a carbapenem-susceptible A. baumannii strain) could further validate the resistance phenotype, such assays had previously been performed and reported [16]. In our earlier work, conventional conjugation assays using azide-resistant, carbapenem-susceptible A. baumannii recipients did not yield transconjugants for plasmids pAC1633-1 and pAC1530, despite multiple attempts [20]. This finding was attributed to the insertion of a 42 kb fragment containing the ISAba1-flanked Tn6948 element, which disrupted the conjugative transfer region and likely reduced the transfer efficiency below detectable levels. Given these prior findings and the current study’s focus on comparative genomic characterization rather than functional assays, additional plasmid transfer experiments were not repeated here. Nevertheless, the detection of similar plasmids among clonally distinct A. baumannii isolates over subsequent years provides strong genomic evidence of horizontal dissemination within the hospital environment.
One of the structural variations discovered in these blaNDM-1-encoded plasmids lies within the Tn125 transposon that harbours blaNDM-1. In most Acinetobacter isolates, including A. nosocomialis AC1530 and A. baumannii AC1633, Tn125 is flanked by two ISAba125 elements. However, in three of the four newly characterized blaNDM-1-encoding plasmids, only a single ISAba125 remains. In the fourth plasmid, pAC1932-1, one of the ISAba125 elements has been replaced by ISAba22. In all four plasmids, the ISAba125 that remained was the copy located immediately upstream of blaNDM-1 (Fig. 2). This copy of ISAba125 provides the external promoter that drives the expression of blaNDM-1 [9], which accounts for the carbapenem-resistant phenotype observed in all four A. baumannii isolates. The production of MBLs in these isolates was confirmed using the Etest MBL kit (bioMérieux, La Balme-les-Grottes, France). Such structural variations in Tn125 have been reported previously, and in these reports, it was always the ISAba125 downstream of blaNDM-1 that was either absent or replaced by another IS element [8464849]. The ISAba125 upstream of blaNDM-1 is required for its expression and is, thus, more often conserved than other genes or elements within Tn125 [45].
Two of the A. baumannii HSNZ isolates (i.e. AC1839 and AC2013) also co-harboured the blaOXA-58 carbapenemase gene, similar to A. nosocomialis AC1530 and A. baumannii AC1633 [20]. In contrast, blaOXA-58 was absent in the genomes of AC1932 and AC2014. Despite this variation, all four A. baumannii isolates, along with A. baumannii AC1633, exhibit carbapenem resistance with comparable MIC values for imipenem, meropenem and ertapenem (Table 2), indicating that blaNDM-1 is likely the primary driver of carbapenem resistance in these isolates. In A. pittii 44551 that co-harbours blaNDM-1 and blaOXA-58 in separate plasmids, carbapenem resistance was attributed to blaNDM-1, while blaOXA-58 remained silent, likely due to the absence of an upstream promoter to drive its expression in plasmid pOXA58-44551 [62]. In the A. pittii 44551 plasmid pNDM-44551, blaNDM-1 was found within a truncated Tn125 structure, whereby the ISAba125 downstream of blaNDM-1 was absent, but the ISAba125 upstream of blaNDM-1, which provided the promoter for its expression, was present [62]. The sequences immediately upstream of blaOXA-58 in pAC1839-1, pAC2013-1 and pAC1633-1 were identical to pOXA58-44551 (i.e. a 369 bp truncated ISAba3 adjacent to blaOXA-58). This suggests that blaOXA-58 may be transcriptionally silent or expressed at very low levels in pAC1839-1, pAC2013-1 and pAC1633-1, but such a postulation warrants further experimental validation through transcriptomic analysis, which is beyond the scope of this paper. Nonetheless, the genomic data, together with biochemical confirmation of MBL activity (via the Etest MBL kit and the BD Phoenix M50 susceptibility test results), provide strong evidence supporting the functional relevance of the identified carbapenemase genes.
Downstream of blaOXA-58, the genetic environment differs. In the Malaysian A. baumannii plasmids, blaOXA-58 is followed by ISAba11 and ISAba3, forming the blaOXA-58 xrs module (see Fig. 4), whereas ISAba11 is absent in pOXA58-44551, and its xrs module comprises only of ΔISAba3-blaOXA-53-ISAba3 [62]. Although xrs modules have been found in various bacterial genera, they are most extensively characterized in Acinetobacter, where their frequent transmission has been well documented [2139]. Alongside IS elements, integrons and transposons, xrs modules contribute significantly to the evolutionary dynamics of Acinetobacter plasmids [3963]. These xrs modules are the target sites for the XerC–XerD site-specific tyrosine recombinase system. While the canonical function of this system is to resolve plasmid multimers into monomers to ensure stable inheritance [64], it is increasingly recognized as a key driver of plasmid structural evolution. The presence of compatible xrs sites on different plasmids allows the system to be hijacked, mediating inter-molecular recombination and fostering the formation of plasmid cointegrates. The subsequent resolution of these cointegrates can then facilitate the exchange and shuffling of genetic modules bounded by xrs sites [2565]. As recently highlighted by Blanchais et al. [22], this interplay between the Xer system and plasmid-carried xrs sites is a powerful mechanism for the dissemination of antibiotic resistance. Moreover, the presence of a plasmid-encoded xerC gene, as we identified in the large blaNDM-1 plasmids of this study (Fig. 1), likely enhances this process. It provides the necessary recombinase encoded by the plasmid itself, making the formation of cointegrates and subsequent gene exchange less dependent on the host chromosomal Xer machinery. Our findings support this, as the movement of the blaNDM-1-encoded large plasmid across different A. baumannii clonal lineages within the same hospital showcases structural fluidity mostly driven by the combined activity of IS elements and Xer-mediated recombination at xrs modules. Further work will be required to validate this hypothesis experimentally. For instance, creating a targeted knockout of the plasmid-encoded xerC gene and assessing its impact on plasmid cointegrate formation and xrs module mobility in a suitable Acinetobacter host would provide direct evidence of its specific function.
In conclusion, this study builds upon our previous work that characterized a large, ca. 170 kb plasmid encoding blaNDM-1, blaOXA-58 and a host of other AMR genes, originally identified in carbapenem-resistant, MDR A. nosocomialis and A. baumannii in HSNZ in 2015 and 2016, respectively [20]. Genome sequencing of A. baumannii isolates from the same hospital in subsequent years revealed that this plasmid continues to be transmitted, as it is now found in distinct, clonally unrelated A. baumannii lineages. Comparative analysis of the complete plasmid sequences demonstrates structural fluidity, primarily mediated by IS elements and xrs modules. In two of these plasmids (pAC1932-1 and pAC2014-1), the blaOXA-58 xrs module and several adjacent xrs modules are absent. However, there were no differences in carbapenem MIC values between these isolates and those harbouring both blaNDM-1 and blaOXA-58, suggesting that blaNDM-1 is the primary contributor to the carbapenem resistance phenotype. The blaOXA-58 gene is likely either silent or expressed at very low levels, possibly due to the absence of an upstream promoter. Transcriptomic analysis would provide clarity on the expression dynamics of these carbapenemase genes, and the absence of such data represents a limitation of this study. Thus, transcriptomic profiling of these A. baumannii strains is recommended as an important direction for future research.
In plasmid pAC1839-1, a novel presumptive 15 kb transposon that harbours a type III CBASS phage defence system was identified, although its function and biological significance remain currently unknown. A. baumannii AC1839 also harbours an additional plasmid, pAC1839-2, which carries the tetA(39)-tetR tetracycline resistance gene pair in a xrs module. This identical module has been found in several Acinetobacter isolates within diverse Rep_3 family plasmids, underscoring its mobility. What we proposed 4 years ago in our initial report [20], that this plasmid is capable of ongoing horizontal transfer, remains valid today. Thus far, the plasmid has only been identified in non-Global Clone (non-GC) A. baumannii lineages. However, in HSNZ and many hospitals globally, the predominant lineage is Global Clone 2 (GC2) [2660]. Should this plasmid be acquired by a GC2 strain, its dissemination potential would likely increase significantly. These findings underscore the urgent need for continuous genomic surveillance, particularly for a pathogen that remains at the top of the World Health Organization’s critical priority pathogen list [3], and also highlight the utility of long-read sequencing in understanding the complex genomic rearrangements that underlie the lateral transfer of AMR genes and other mobile elements.
Supplementary material
10.1099/mgen.0.001630Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Thacharodi A Vithlani A Hassan S Alqahtani A Pugazhendhi A Carbapenem-resistant Acinetobacter baumannii raises global alarm for new antibiotic regimensi Science 20242711136710.1016/j.isci.2024.11136739650735 PMC 11625361 · doi ↗ · pubmed ↗
- 2Zampaloni C Mattei P Bleicher K Winther L Thäte C et al A novel antibiotic class targeting the lipopolysaccharide transporter Nature 202462556657110.1038/s 41586-023-06873-038172634 PMC 10794144 · doi ↗ · pubmed ↗
- 3World Health Organization (WHO) WHO bacterial priority pathogens list, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance 2024 https://www.who.int/publications/i/item/978924009346110.1016/S 1473-3099(25)00118-5PMC 1236759340245910 · doi ↗ · pubmed ↗
- 4Isler B Doi Y Bonomo RA Paterson DL New treatment options against carbapenem-resistant Acinetobacter baumannii infections Antimicrob Agents Chemother 201963 e 01110-1810.1128/AAC.01110-18PMC 632523730323035 · doi ↗ · pubmed ↗
- 5Kinross P Gagliotti C Merk H Plachouras D Monnet DL et al Large increase in bloodstream infections with carbapenem-resistant Acinetobacter species during the first 2 years of the COVID-19 pandemic, EU/EEA, 2020 and 2021 Euro Surveill 2022271710.2807/1560-7917.ES.2022.27.46.2200845 PMC 967323536398574 · doi ↗ · pubmed ↗
- 6Miethke M Pieroni M Weber T Brönstrup M Hammann P et al Towards the sustainable discovery and development of new antibiotics Nat Rev Chem 2021572674910.1038/s 41570-021-00313-1PMC 837442534426795 · doi ↗ · pubmed ↗
- 7Nikibakhsh M Firoozeh F Badmasti F Kabir K Zibaei M Molecular study of metallo-β-lactamases and integrons in Acinetobacter baumannii isolates from burn patients BMC Infect Dis 20212178210.1186/s 12879-021-06513-w 34372787 PMC 8353788 · doi ↗ · pubmed ↗
- 8Tang B Wang C Sun D Lin H Ma J et al In silico characterization of blandm-harboring conjugative plasmids in Acinetobacter species Microbiol Spectr 202210 e 021022210.1128/spectrum.02102-2236301090 PMC 9769834 · doi ↗ · pubmed ↗