Critique of a Radically New Model for Plasma Membrane Bilayer Organization
Yvonne Lange, Theodore L. Steck

TL;DR
This paper critiques a new model of plasma membrane organization that contradicts existing scientific understanding.
Contribution
The paper identifies flaws in a novel model of lipid distribution in cell membranes.
Findings
Incomplete digestion by phospholipase A2 can be explained by product inhibition.
Ultrastructural analysis shows most phosphatidylcholine is in the outer leaflet.
High cholesterol concentration in the outer leaflet is not detectable by probes.
Abstract
A novel model recently proposed by Doktorova et al. challenges current concepts for the molecular organization of the plasma membrane bilayer. It is at odds with previously published research. The model posits that there are far fewer phospholipid molecules in the outer leaflet of the bilayer than in the inner leaflet and that the resulting area deficit is filled by cholesterol. This conclusion is based on the incomplete hydrolysis of the phosphatidylcholine in intact erythrocytes by phospholipase A2, leading to the inference that the undigested fraction is endofacial. But the incomplete digestion can be explained by product inhibition. Furthermore, ultrastructural analysis has shown that almost all of the phosphatidylcholine in the erythrocyte bilayer resides in the outer leaflet. Finally, the high concentration of cholesterol predicted for the outer leaflet of resting human…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
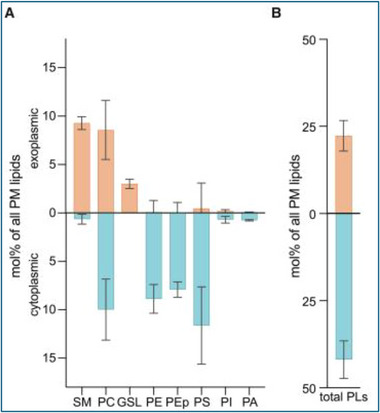 FIGURE 1
FIGURE 1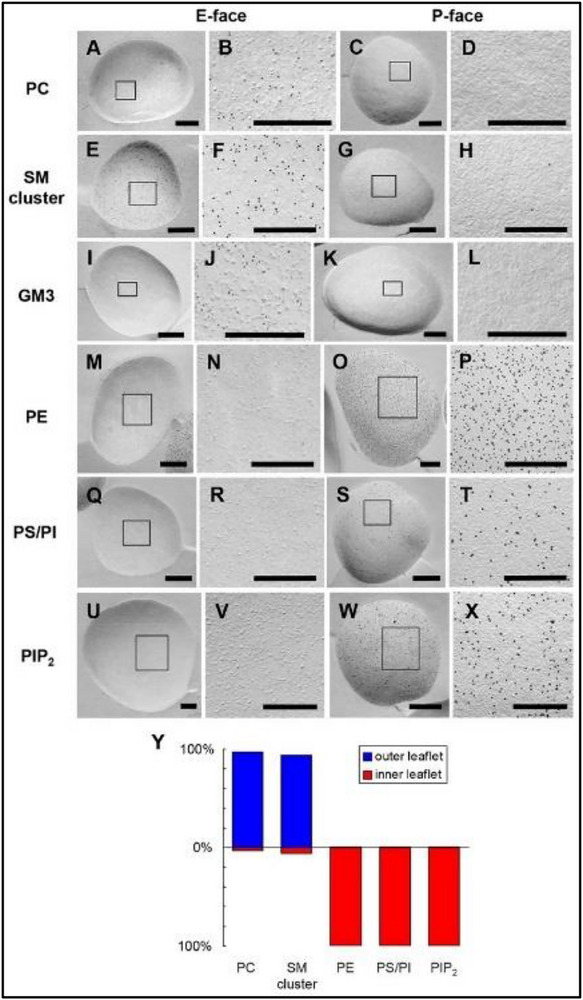 FIGURE 2
FIGURE 2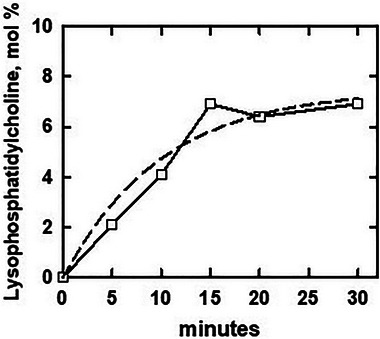 FIGURE 3
FIGURE 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsErythrocyte Function and Pathophysiology · Lipid Membrane Structure and Behavior · Cellular transport and secretion
Introduction
1
It is generally held that eukaryotic plasma membranes are built upon a bilayer of phospholipids with glycolipids, sterols, and proteins integrated therein. The asymmetrical transbilayer disposition of individual proteins and the exofacial location of glycolipids are well established [1, 2, 3]. The transverse asymmetry of human erythrocyte plasma membrane phospholipids has also been characterized [4]. Nearly all of the sphingomyelin is accessible to probes in the intact cell; hence, it is exposed at the exofacial side. Phosphatidylethanolamine, phosphatidylserine, and phosphatidylinositol and its derivatives resist cleavage by phospholipase A2 and have therefore been assigned to the cytoplasmic leaflet [1, 5, 6, 7, 8, 9]. On the other hand, the transbilayer distributions of both phosphatidylcholine and cholesterol are unsettled [1, 10, 11].
A recent publication by Doktorova et al. presents a new model for plasma membrane bilayer organization that differs dramatically from current concepts [9]. It derives from their determination that only about half of the phosphatidylcholine in intact red cell membranes is hydrolyzed by phospholipase A2 (Figure 1). Four key inferences were drawn from this finding: (a) The undigested phosphatidylcholine resides in the cytoplasmic leaflet of the bilayer. (b) This result was taken to mean that the number of phospholipid molecules in the endofacial leaflet greatly exceeds that on the exofacial side, possibly by as much as two‐fold. (c) It follows that there is a large difference in the total cross‐sectional area of the phospholipids in the two leaflets. (d) Given that the two leaflets have essentially equal surface areas overall, it was proposed that the gap in the outer leaflet is filled by cholesterol [9]. Simulations predicted that ∼77% of the bilayer sterol is in the exofacial leaflet and ∼23% is in the contralateral leaflet.
Proposed transbilayer distribution of the polar lipids in the human erythrocyte membrane [9]. Intact cells were digested with lipases. The fraction of each lipid that was hydrolyzed was assigned to the exofacial leaflet and the remainder to the contralateral leaflet. Cholesterol is not shown but is included in the total plasma membrane lipids. Note that more than half of the phosphatidylcholine is assigned to the cytoplasmic leaflet. Reproduced from Doktorova et al. with permission. GSL, glycosphingolipid; PA, phosphatidic acid; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PEp, PE plasmalogen; PI, phosphatidylinositol; PL, phospholipid; PM, plasma membrane; PS, phosphatidylserine; SM, sphingomyelin.
Countervailing Literature
2
The model advanced by Doktorova et al. for the molecular organization of the plasma membrane bilayer [9] is at odds with the following three lines of evidence, none of which they addressed.
Essentially All of the Phosphatidylcholine is Exofacial
2.1
The distribution of polar lipids between the two leaflets of the red cell plasma membrane bilayer has been analyzed with specific immuno‐gold tagged protein probes using “sodium dodecyl sulfate‐digested freeze‐fracture replica labeling” or SDS‐F [6, 12]. Telling electron microscope images are reproduced in Figure 2. Concordant with many enzyme digestion studies, this technique placed all of the phosphatidylserine, phosphatidylethanolamine, phosphatidylinositol and phosphatidylinositol 4,5‐bisphosphate in the inner leaflet. In addition, almost all of the membrane sphingomyelin and ganglioside GM3 were found in the outer leaflet. Crucially, nearly all of the phosphatidylcholine was exofacial.
Ultrastructural analysis of the distribution of polar lipids between the two leaflets of the human erythrocyte bilayer. The membranes were cleaved by freeze‐fracture and decorated with the indicated lipid‐specific probes. The E‐face shows the outer leaflet, and the P‐face shows the inner leaflet at both low and high magnification. The relevant finding is that nearly all of the phosphatidylcholine was found in the outer leaflet. Reproduced from Murate et al. with permission [6]. GM3, ganglioside GM3; PC, phosphatidylcholine; PIP2, phosphatidylinositol 4,5‐bisphosphate; SM, sphingomyelin.
The diametrical distribution of each of the lipids between the two leaflets in these studies provides an internal control. Furthermore, the fidelity of the technique was corroborated with liposomes composed of different saturated and unsaturated phosphatidylcholines as well as other phospholipids [6, 12, 13]. In particular, control experiments showed that anionic phospholipids do not interfere with the labeling of phosphatidylcholine in vesicles. This lends assurance that the detection of phosphatidylcholine at the endofacial surface of the red cell membrane would not have been hindered by the negatively charged phospholipids therein [6]. These ultrastructural data contrast strongly with the phospholipase digestion results which were taken to signify that half or more of the phosphatidylcholine is located in the cytoplasmic leaflet of the membrane.
Why Does Plasma Membrane Phosphatidylcholine Resist Digestion?
2.2
Doktorova et al. determined that about half of the phosphatidylcholine in intact red cells was not hydrolyzed by phospholipase A2 and proposed that it must therefore be endofacial (Figure 1) [8, 9]. However, there is a sampling bias in this approach: probing just the outer leaflet of the plasma membrane bilayer risks underestimating any unreacted exofacial phospholipid. Indeed, in an early study, a combination of sphingomyelinase and phospholipase A2 hydrolyzed 48% of the phospholipid in intact red cells, positive evidence that there is no phospholipid deficit in the outer leaflet.
The large variability of the undigested fraction suggests an experimental rather than a biological cause for the observed incomplete hydrolysis of phosphatidylcholine (Figure 1) [9]. Furthermore, factors other than the sidedness of the phosphatidylcholine can limit its digestion [14, 15, 16, 17, 18, 19, 20]. In particular, product inhibition can prematurely curtail the attack of phospholipase A2 on its substrate [21]. This effect has been observed for the honeybee enzyme used by Doktorova et al. [20, 22, 23]. Indeed, the progress curve for digestion underlying the study in question suggests product inhibition [8]. That is, the time course appears to show a more abrupt deceleration than is predicted for Michaelis–Menton kinetics (Figure 3). (The reaction also shows a slight initial acceleration ascribable to early product activation of the enzyme [17, 24].) Both chemical and physical mechanisms have been proposed to explain product inhibition. In the former case, the products of phospholipid hydrolysis are competitive inhibitors of the enzyme [21, 25, 26, 27, 28]. The latter mechanism implicates the increase in membrane surface pressure imposed by the hydrolysis products: compressing the bilayer makes the substrate less vulnerable to further attack [20, 29, 30].
Hydrolysis of the phosphatidylcholine in intact human erythrocytes by phospholipase A2. The data for lysophosphatidylcholine production were replotted from Figure S1E in Lorent et al. [8]. For comparison, we generated the dashed curve assuming Michaelis–Menton kinetics. The question at issue is whether the flattening of the experimental curve at 15 min is due to product (or another type of) inhibition rather than exhaustion of the exofacial substrate as proposed [8, 9].
Our analysis of the primary data in Figure 3 does not settle the question of product inhibition. A more definitive time course is needed. In addition, future work could test for the putative undigested phosphatidylcholine in the outer leaflet of the intact cell. Adding sphingomyelinase to the phospholipase A2 increases the digestion of phosphatidylcholine in intact red cells. In one such study, the phosphatidylcholine was hydrolyzed almost as completely as was the sphingomyelin: 76% versus 82% [31]. Depletion of plasma membrane cholesterol with methyl‐β‐cyclodextrin might also increase the susceptibility of the phosphatidylcholine [19]. In addition, serum albumin can relieve product inhibition by sequestering the inhibitory digestion products [21]. (However, their removal might promote hemolysis.)
Another factor limiting the action of phospholipase A2 on the exofacial face of plasma membrane bilayers is the close packing of their saturated phospholipids [8, 14, 32, 33]. If, as we suspect, the exofacial leaflet is replete in saturated phosphatidylcholines that resist digestion, the extent of chain saturation and tight packing in the outer leaflet would be even greater than proposed [8].
In any case, why phospholipase A2 digestion routinely fails to hydrolyze much of the plasma membrane phosphatidylcholine is presently uncertain. This makes the conclusion of Doktorova et al. premature [9].
Evidence for Excess Cholesterol at the Erythrocyte Surface is Lacking
2.3
The cholesterol in monolayers, vesicles, and plasma membranes is almost entirely complexed with the phospholipids [34, 35, 36, 37, 38]. These studies showed that only cholesterol exceeding stoichiometric equivalence is readily accessible to ligands and probes such as cholesterol oxidase, methyl‐β‐cyclodextrin, and bacterial toxins. It can be estimated from the data presented by Doktorova et al. [9] that the cholesterol/phospholipid ratio in the outer leaflet of the red cell bilayer is ∼1.45 mole/mole (see Box 1). This high level of cholesterol should be readily detected at the surface of unperturbed human erythrocytes. However, it is not [11, 36, 39, 40, 41].
Doktorova et al. did a nice experiment to estimate the transverse distribution of sterols [9]. They labeled erythrocytes with a small amount of dehydroergosterol (DHE), a good cholesterol surrogate and reporter that presumably equilibrates across the bilayer. The addition of Di4, a cationic fluorescence quencher that cannot cross the bilayer, reduced the DHE fluorescence in the bilayer by about two‐thirds. This is taken as the fraction of DHE (and, presumably, cholesterol) located in the outer leaflet, supporting their premise that about three‐quarters of the sterol is exofacial.
BOX 1 | Calculation of leaflet cholesterol/phospholipid ratios from Doktorova et al. [9]
- The average mole ratio of inner leaflet (i) to outer leaflet (o) phospholipid (PL) is ∼1.8 (their Figure 1D). The inner leaflet therefore has 64.3%, and the outer leaflet has 35.7% of the total phospholipid (PL_t_). For 100 moles of total phospholipid, PL_i_ = 64.3 and PL_o_ = 35.7 moles.
- Cholesterol (CH) constitutes 40 mol% of the total membrane lipid [i.e.,100 × CH_t_/(CH_t_ + PL_t_)]. This corresponds to CH_t_/PL_t_ = 0.67. That is, the concentration of membrane cholesterol is 67 moles/100 moles phospholipid.
- The molecular dynamics simulation predicts that 77% of the cholesterol is exofacial and 23% is endofacial. Thus, CH_o_ = 51.6 and CH_i_ = 15.4 moles per 100 moles phospholipid.
- It follows that the lipid mole ratio in the outer leaflet is CH_o_/PL_o_ = 51.6/35.7 = 1.45 and the lipid mole ratio in the inner leaflet is CH_i_/PL_i_ = 15.4/64.3 = 0.24.
Other mechanisms might lead to excess exofacial cholesterol [8, 10, 11, 42, 43, 44, 45]. In some studies, the driver of the transbilayer asymmetry of the sterol is taken to be its preferential association with the saturated phospholipids abundant in the outer leaflet rather than the surface area compensation postulated by Doktorova et al. [9]. The association of the cholesterol with the saturated phospholipids in the exofacial leaflet should reduce its chemical activity and therefore its reactivity with probes. This is not predicted by the Doktorova model [9] but is found experimentally [9, 11, 36, 39, 40, 41].
Conclusions
3
Three lines of evidence counter the proposal by Doktorova et al. that a large fraction of the phosphatidylcholine in the erythrocyte plasma membrane resides in its endofacial leaflet (Figure 1) and that the imbalance in phospholipid area across the bilayer is compensated for by an excess of exofacial cholesterol. First, an ultrastructural analysis of the binding of phospholipid‐specific ligands showed that almost all of the phosphatidylcholine in this membrane is in its outer leaflet (Figure 2). Second, both the phospholipase A2 literature and the time course of digestion reported by Lorent et al. (Figure 3) suggest that the limited hydrolysis of phosphatidylcholine they observed might not signify its sequestration in the cytoplasmic leaflet but rather its incomplete digestion at the exofacial surface. Third, the high level of uncomplexed cholesterol they proposed for the outer leaflet of the resting human erythrocyte should be readily detectable by various probes, but it is not.
One more consideration: in sheep erythrocytes, the phosphatidylcholine is almost completely replaced by sphingomyelin [46, 47, 48, 49]. This lipid constitutes about half of the total phospholipid complement in these cells. If, as reviewed above, sphingomyelin is confined to the outer leaflet of the erythrocyte bilayer, sheep red cells would have no exofacial area deficit and, therefore, no sterol excess.
Doktorova et al. did not address these issues. Rather, they supported their model with a variety of ancillary experiments and conceptual tests [9]. Those data and insights are valuable. However, they do not rule out other models for the distribution of lipids across the plasma membrane bilayer and are not sufficient to establish theirs.
Author Contributions
The authors contributed equally to the conception and writing of the manuscript.
Funding
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1M. Murate and T. Kobayashi , “Revisiting Transbilayer Distribution of Lipids in the Plasma Membrane,” Chemistry and Physics of Lipids 194 (2016): 58–71.26319805 10.1016/j.chemphyslip.2015.08.009 · doi ↗ · pubmed ↗
- 2P. Mattjus , “Specificity of the Mammalian Glycolipid Transfer Proteins,” Chemistry and Physics of Lipids 194 (2016): 72–78.26234207 10.1016/j.chemphyslip.2015.07.018 · doi ↗ · pubmed ↗
- 3G. Pabst and S. Keller , “Exploring Membrane Asymmetry and Its Effects on Membrane Proteins,” Trends in Biochemical Sciences 49, no. 4 (2024): 333–345.38355393 10.1016/j.tibs.2024.01.007 · doi ↗ · pubmed ↗
- 4J. Op den Kamp , “Lipid Asymmetry in Membranes,” Annual Review of Biochemistry 48 (1979): 47–71.10.1146/annurev.bi.48.070179.000403382989 · doi ↗ · pubmed ↗
- 5A. Zachowski , “Phospholipids in Animal Eukaryotic Membranes—Transverse Asymmetry and Movement,” Biochemical Journal 294 (1993): 1–14.8363559 10.1042/bj 2940001 PMC 1134557 · doi ↗ · pubmed ↗
- 6M. Murate , M. Abe , K. Kasahara , K. Iwabuchi , M. Umeda , and T. Kobayashi , “Transbilayer Distribution of Lipids at Nano Scale,” Journal of Cell Science 128, no. 8 (2015): 1627–1638.25673880 10.1242/jcs.163105 · doi ↗ · pubmed ↗
- 7R. Clarke , K. Hossain , and K. Cao , “Physiological Roles of Transverse Lipid Asymmetry of Animal Membranes,” Biochimica et Biophysica Acta ‐ Biomembranes 1862, no. 10 (2020): 183382.32511979 10.1016/j.bbamem.2020.183382 · doi ↗ · pubmed ↗
- 8J. H. Lorent , K. R. Levental , L. Ganesan , et al., “Plasma Membranes Are Asymmetric in Lipid Unsaturation, Packing and Protein Shape,” Nature Chemical Biology 16, no. 6 (2020): 644–652.32367017 10.1038/s 41589-020-0529-6PMC 7246138 · doi ↗ · pubmed ↗