Dual resistance to carbapenems and colistin in Enterobacter: Taiwan surveillance of antimicrobial resistance, 2010–2020
Ying-Chi Huang, Tzu-Wen Huang, Praveen Rahi, Shu-Chen Kuo, Yan-Ru Chen, Chi-Tai Fang

TL;DR
A study in Taiwan found that a significant portion of Enterobacter bacteria are resistant to both carbapenems and colistin, highlighting a growing threat to public health.
Contribution
The study reveals the prevalence and genetic mechanisms of dual carbapenem and colistin resistance in Enterobacter isolates in Taiwan over a decade.
Findings
35 out of 41 carbapenem-resistant Enterobacter isolates exhibited dual resistance to colistin.
Colistin resistance was mainly due to activation of the arnBCADTEF operon, not mcr genes.
A conserved IncHI2 plasmid carrying blaIMP-8 and mcr-9 circulated across Enterobacter species for over 10 years.
Abstract
Extensively drug-resistant gram-negative bacteria harbouring dual resistance to carbapenems and colistin represent a critical global health threat. A total of 929 population-representative Enterobacter isolates were systematically collected from 29 hospitals across four regions of Taiwan between 2010 and 2020. Forty-one isolates (4.4%) were nonsusceptible to carbapenems and underwent whole-genome sequencing, resistance gene profiling, plasmid analysis, and antimicrobial susceptibility testing (AST). Among them, 35 isolates (85.4%) exhibited dual resistance to carbapenems and colistin; however, only half (17/35) were detectable by standard phenotypic AST. Colistin resistance was primarily mediated by activation of the chromosomal arnBCADTEF operon, which was frequently inducible and often undetected by standard testing, rather than by mcr-9 or mcr-10. A conserved IncHI2 plasmid carrying…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
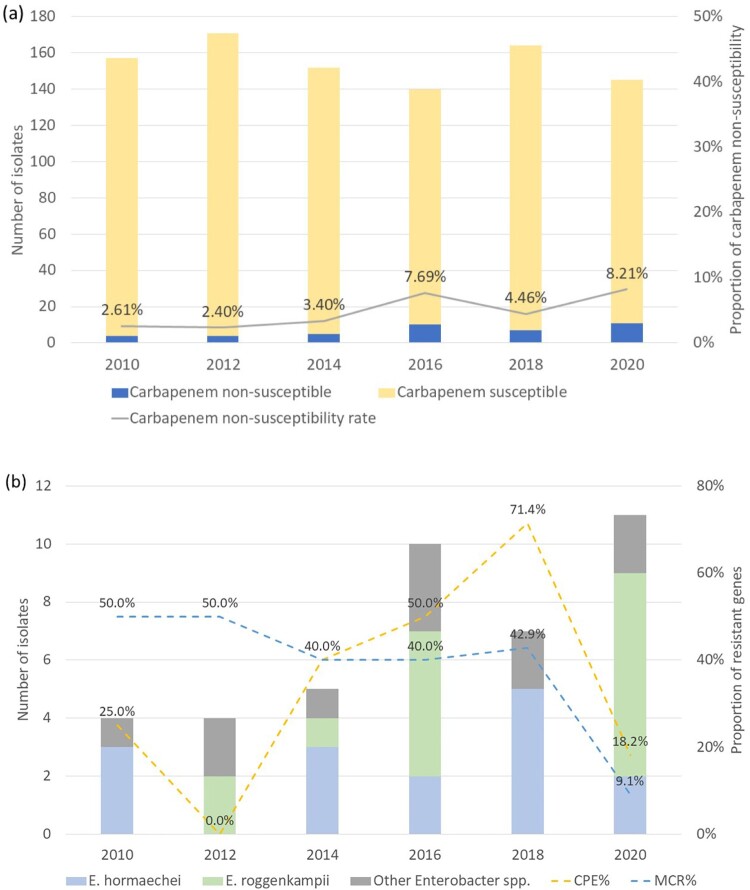 Figure 1
Figure 1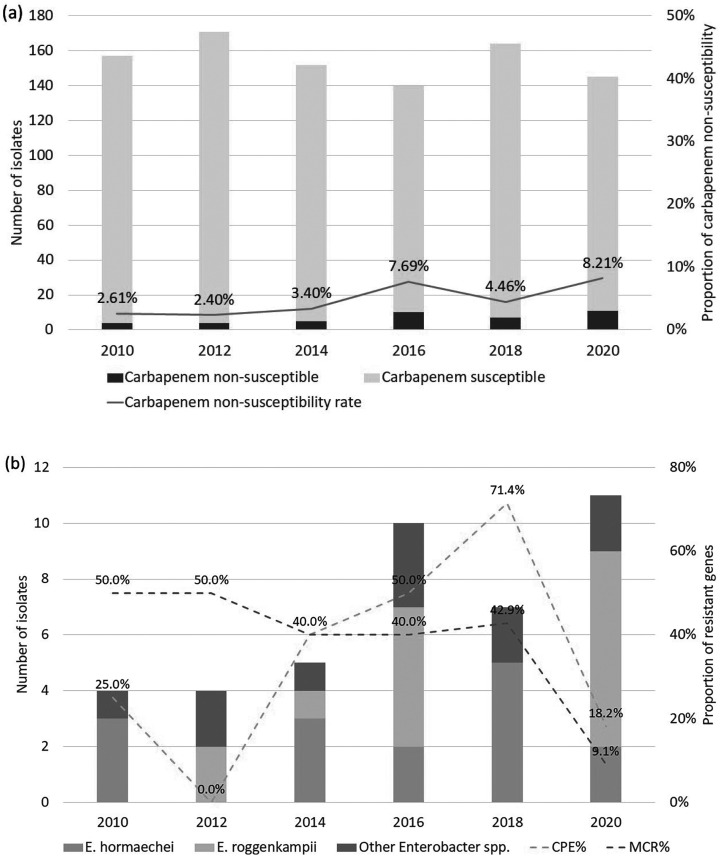 Figure 2
Figure 2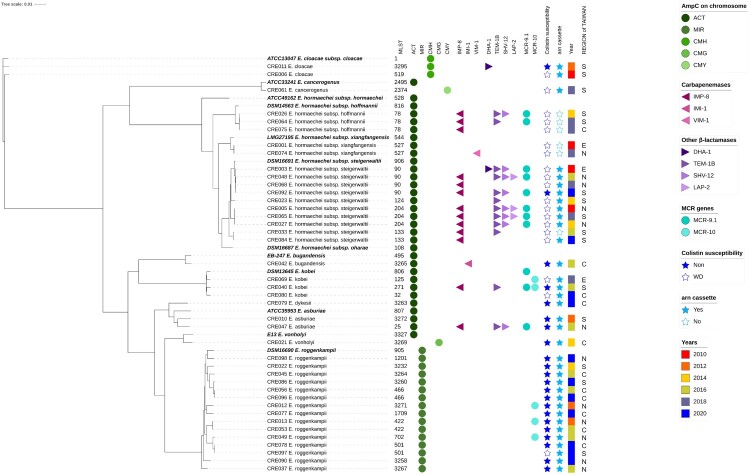 Figure 3
Figure 3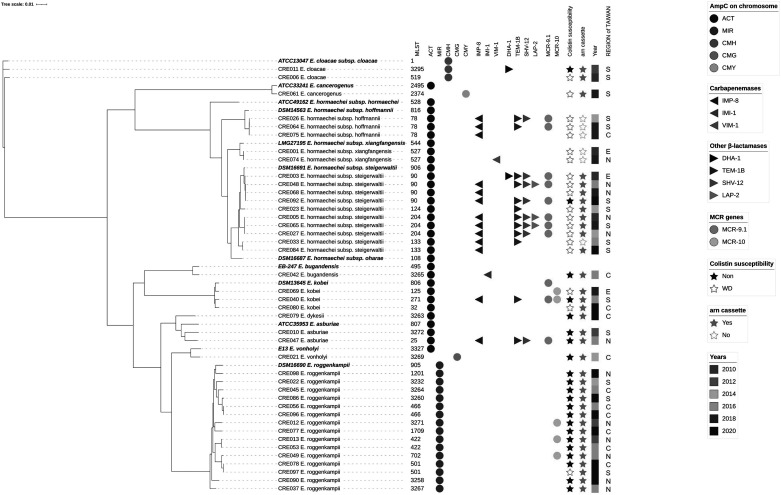 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6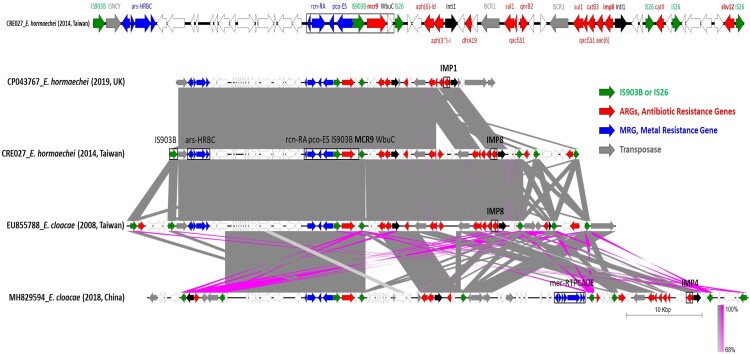 Figure 7
Figure 7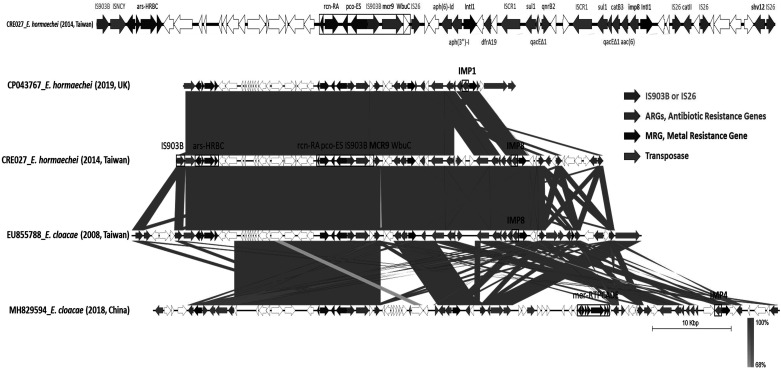 Figure 8
Figure 8- —National Health Research Institutes10.13039/501100004737
- —Taiwan National Science and Technology Council10.13039/100020595
- —Taiwan Ministry of Education
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Escherichia coli research studies · Antibiotics Pharmacokinetics and Efficacy
Introduction
Enterobacter species have emerged as important nosocomial pathogens, ranking third among Enterobacterales after Escherichia coli and Klebsiella pneumoniae [1–3]. Enterobacter bacteraemia is associated with attributable mortality rates approaching 40% [4]. Treatment is challenging because intrinsic AmpC β-lactamase production and widespread fluoroquinolone resistance limit therapeutic options, leaving carbapenems as essential agents [5]. Alarmingly, carbapenem resistance in Enterobacter is increasing worldwide, mediated by carbapenemases or by AmpC/ESBL production combined with porin loss [3,6].
Colistin, often reserved as a last-line agent, is now also facing increasing resistance in Enterobacter spp. [7–9]. Plasmid-mediated mcr genes (mcr-1 to mcr-10) have emerged as transferable determinants of colistin resistance in Enterobacterales, with mcr-9 and mcr-10 especially relevant in Enterobacter spp. [7,10]. Doijad et al. recently demonstrated that colistin resistance in Enterobacter can also be mediated by the chromosomal arnBCADTEF operon (also named pmrHFIJKLM operon), which modifies lipid A through the addition of 4-amino-4-deoxy-L-arabinose (L-Ara4N), in isolates from German hospitals [9]. Expression of the arn operon is regulated mainly by PhoPQ two component system and can be induced upon colistin exposure, resulting in resistant subpopulations escape detection by routine susceptibility testing [8,9]. Both in vivo and clinical treatment failure have been documented in association with this mechanism [11,12]. Notably, a high prevalence of colistin heteroresistance (approximately 28%) has been reported among Enterobacter clinical isolates in Japan, with clustering in specific species and lineages within a single medical centre [13]. Despite detailed mechanistic characterization, the population-level prevalence and clinical impact of arn-mediated colistin resistance remain poorly defined at a global scale.
In Taiwan, the nationwide Taiwan Surveillance of Antimicrobial Resistance (TSAR), established in 1998, has systematically collected clinical isolates from more than 25 hospitals across four geographic regions [2,3]. Carbapenem resistance in Enterobacter has been linked to blaIMP-8, a metallo-β-lactamase that compromises most novel β-lactam/β-lactamase inhibitor combinations [3]. The convergence of carbapenemase blaIMP-8 with colistin resistance would leave few therapeutic options.
This study aimed to describe this emerging antimicrobial resistance pattern of major global concern – the convergence of carbapenem and colistin resistance in Enterobacter, using the population-based, multicentre TSAR programme. We further characterized the genetic basis of this dual resistance through whole-genome sequencing.
Methods
Isolate collection
Isolates were obtained through the TSAR programme from 29 hospitals across 4 regions of Taiwan between 2010 and 2020 biennially [2,3]. Participating hospitals comprised 12 medical centres and 17 regional hospitals. In each surveillance year, bacterial isolates were collected sequentially from clinical specimens between July and September without species preselection. All isolates were identified by MALDI-TOF mass spectrometry (Bruker Daltonics, USA), and antimicrobial susceptibility testing (AST) was performed following species identification. Only isolates identified as Enterobacter spp. were included in the present study. Demographic and clinical information provided by the hospitals included patient age, specimen type, and ward type (intensive care unit [ICU], non-ICU inpatient ward, or outpatient setting including the emergency department). Patients were categorized into three age groups: children (<18 years), adults (18–64 years), and elderly (≥65 years). The protocol was approved by the Institutional Review Board of the National Health Research Institutes (EC1101105-E).
Antimicrobial susceptibility testing (AST)
Minimal inhibitory concentrations (MIC) to amikacin, aztreonam, cefepime, cefotaxime, ciprofloxacin, colistin, gentamicin, ertapenem, imipenem, meropenem, levofloxacin, piperacillin–tazobactam, tigecycline, and trimethoprim–sulfamethoxazole were determined by broth microdilution (Thermo Fisher Scientific, UK) according to CLSI guidelines [14]. Susceptibility to newer β-lactam/β-lactamase inhibitor combinations (meropenem–vaborbactam, imipenem–relebactam, and ceftazidime–avibactam) was determined by in-house broth microdilution.
The U.S. CDC carbapenem-resistant Enterobacterales (CRE) definitions define CRE as exhibiting non-susceptibility to at least one carbapenem or evidence of carbapenemase production [15]. Accordingly, in the present study, isolates with imipenem MIC ≥ 2 µg/mL were classified as carbapenem non-susceptible [14], because some isolates classified as susceptible to meropenem were not susceptible to imipenem. Colistin MICs ≤ 2 µg/mL were interpreted as wild-type susceptibility [14], and tigecycline non-susceptibility was defined as MIC > 2 µg/mL according to EUCAST breakpoints(https://www.eucast.org/clinical_breakpoints).
Broth macrodilution of colistin susceptibility and inducible resistance testing
Colistin susceptibility testing was performed by the broth macrodilution method in accordance with CLSI guidelines [16]. Sterile PYREX® glass test tubes (12 × 75 mm) fitted with metal closures were used for all dilutions. Bacterial isolates were obtained from overnight cultures on blood agar plates. Three to five well-isolated colonies of identical morphology were selected and suspended in sterile saline. The turbidity of the bacterial suspension was adjusted to a 0.5 McFarland standard. Antimicrobial stock solutions were prepared so that the series of colistin dilutions were at twice the desired final concentrations, to account for the 1:2 dilution resulting from addition of equal volumes of inoculum. The 0.5 McFarland suspension was diluted 1:150 in cation-adjusted Mueller–Hinton broth to yield a suspension of approximately 1 × 10^6^ CFU/mL. One millilitre of this inoculum was then added to each test tube containing 1 mL of the appropriate colistin concentration (and to a control tube containing 1 mL of broth only), resulting in a final inoculum of 5 × 10^5^ CFU/mL. Tubes were incubated at 35 ± 2 °C for 16–20 h, and the MIC was recorded as the lowest concentration of colistin that completely inhibited visible bacterial growth. The skipped-well phenomenon was defined as a non-monotonic growth pattern during broth macrodilution testing, characterized by regrowth at higher colistin concentrations. When this phenomenon was observed, MIC values were assigned as the lowest colistin concentration that inhibited visible bacterial growth without regrowth at higher concentrations.
To further evaluate colistin inducible resistance, LB-colistin agar spot assay was conducted using LB agar supplemented with 2 µg/mL colistin (LA_2 µg/mL) [17]. Overnight LB cultures (pH 7.0) were adjusted to an optical density of OD₆₀₀ = 0.9. A 1:10 dilution was prepared by mixing 100 µL of culture with 900 µL phosphate-buffered saline (PBS). From each of the three sources – (i) overnight culture, (ii) OD₆₀₀ = 0.9 refreshed culture, and (iii) 1:10 dilution – 10 µL aliquots were spotted onto LA_2 µg/mL plates. After air-drying, the plates were incubated at 37 °C for 16–20 hours. The appearance of discrete colonies or hazy growth was interpreted as evidence of colistin-tolerant or heteroresistant subpopulations. All experiments were performed in triplicate to ensure reproducibility.
Sample preparation, DNA extraction, and whole genome sequencing (WGS)
From 2010 to 2020, a total of 929 Enterobacter isolates were collected, of which 41 were imipenem non-susceptible. Single colonies of these 41 isolates were cultured overnight and DNA was extracted using the Promega Wizard® Genomic DNA Purification Kit. WGS was performed on the Pacific Biosciences Sequel II platform (https://www.pacb.com). Genome assembly was conducted using ≥150 Mb of HiFi reads (>5 kb) with the Genome Assembly tool in PacBio SMRTLink version 10.2. The assembled genomic sequences were deposited under BioProject accession number PRJNA874798, with accession numbers for individual isolates provided in Supplementary Table S1. Twelve reference genomes used for comparative analysis are provided in Supplementary Table S2 [18].
Species identification and phylogenetic analysis
Species assignment was confirmed by in silico DNA–DNA hybridization (isDDH) using the Genome-to-Genome Distance Calculator (GGDC, cutoff 70%) [19] and by Average Nucleotide Identity (ANI) using QIAGEN CLC Genomics Workbench v9.5.3 [20]. Results are provided in Supplementary Table S1. Core-genome alignment was generated using Panaroo v1.5.2, defining core genes as those present in 99–100% of isolates, yielding 2,760 core genes for phylogenetic reconstruction [21]. Maximum-likelihood phylogenetic analysis was performed using IQ-TREE v2.1.4 with automatic model selection (ModelFinder) based on the concatenated core-gene alignment [22]. Branch support was assessed using 1,000 ultrafast bootstrap replicates and 1,000 SH-aLRT tests (-m MFP – bb 1000 – alrt 1000 – nt AUTO). Support values are shown on the final tree, which was visualized using iTOL v6 webserver (https://itol.embl.de/).
Genomic characterization and comparative analysis
Genome annotation including insertion sequence, virulence gene and integrase was performed using the NCBI Prokaryotic Genome Annotation Pipeline (PGAP) [23]. Multilocus sequence typing (MLST) was carried out using MLST 2.0 [24]. Comprehensive Antibiotic Resistance Database (CARD) was used to detect acquired antimicrobial resistance genes [25]. Plasmid replicon types were determined using the PubMLST plasmid MLST database (https://pubmlst.org/organisms/plasmid-mlst; accessed on 2025/05/27). Plasmid comparisons were also analysed using BRIG (BLAST Ring Image Generator) to visualize structural similarities and differences among plasmid sequences [26]. Promoter prediction was predicted using BPROM [27]. Nucleotide sequence alignments were performed with BLAST [28], and genomic context visualizations were generated with EasyFig [29].
Statistical analysis
The Cochran–Armitage trend test was used to evaluate temporal trends in the proportion of isolates classified as imipenem non-susceptible versus imipenem susceptible, with year of surveillance treated as an ordinal variable. Proportions of clinical characteristics, resistance genes, and antimicrobial susceptibility were compared between E. hormaechei and E. roggenkampii isolates using Fisher’s exact test. Two-sided P values <0.05 were considered statistically significant. All statistical analyses were performed using R version 4.3.2 (R Foundation for Statistical Computing, Vienna, Austria).
Results
Species identification and epidemiology
From 2010 to 2020, 929 Enterobacter isolates were collected, of which 41 (4.6%) were imipenem non-susceptible (32 intermediate, MIC = 2 µg/mL; 9 resistant, MIC > 2 µg/mL). The proportion of imipenem non-susceptible isolates increased over the study period, peaking at 8.2% in 2020 (Cochran–Armitage trend test, P = 0.013) (Figure 1a). Figure 1.(A) Carbapenem non-susceptibility among 929 Enterobacter isolates in Taiwan (2010–2020). Proportion of carbapenem-non-susceptible isolates (grey line). (B) Species distribution of carbapenem non-susceptible Enterobacter isolates, mainly E. roggenkampii and E. hormaechei. The yellow dashed line indicates the proportion of carbapenemase producing isolates. The blue dashed line indicates the proportion of mcr gene carriage.
Among the 41 isolates, blood was the most common source (39%), followed by urine (22%), pus (19.5%), and sputum (14.6%). Rare sources included ascites, catheter tips, and unknown origins, with one isolate from each. Nearly half of the isolates were obtained from elderly patients (≥65 years, 48.8%), and most were recovered from inpatients (75.6%).
WGS-based identification revealed equal numbers of E. hormaechei and E. roggenkampii (15 each, 36.6%), with the remainder including three E. kobei (7.3%), two each of E. asburiae and E. cloacae (4.9% each), and one isolate (2.4%) each of E. bugandensis, E. cancerogenus, E. vonholyi, and E. dykesii (Figure 1b). Core-genome phylogenetic analysis demonstrated clear species-level clustering, with all clinical isolates forming well-supported monophyletic clades corresponding to their respective reference strains (Figure 2). Consistently, all pairwise ANI comparisons among clinical isolates formed distinct clusters consistent with species-level cutoffs (≥95–96%), supporting robust species delineation (Supplementary Figure S1). Figure 2.Phylogenetic tree of 41 carbapenem-non-susceptible Enterobacter isolates collected in Taiwan (2010–2020). Reference strains are shown in bold, and isolates from this study are in regular font. WD indicates colistin wild type (MIC ≤ 2 µg/mL). Resistance determinants, arn operon, mcr genes, and year of collection are annotated alongside each isolate.
Within the E. hormaechei complex (n = 15), isolates further segregated into three distinct subspecies clades: subsp. steigerwaltii (n = 10), subsp. hoffmannii (n = 3), and subsp. xiangfangensis (n = 2), each clustering tightly with its corresponding type strain (accession numbers in Supplementary Table S2). These assignments were further supported by high within-subspecies genomic similarity, with ANI values >98.9% and isDDH values >90% (Supplementary Table S1).
Phylogenetic analysis showed that isolates were geographically interspersed across Taiwan, with no evidence of regional clustering (Figure 2). Isolates from the same species, subspecies, and sequence types were distributed across multiple regions, and carbapenemase genes were predominantly detected in E. hormaechei across diverse sequence types (ST78, ST90, ST133, ST204).
Antimicrobial susceptibility, resistant genes, and virulence genes
Clinical characteristics, antimicrobial susceptibility profiles, and resistance determinants of E. hormaechei, E. roggenkampii, and other Enterobacter species are summarized in Table 1. No significant differences in clinical characteristics were observed between E. hormaechei and E. roggenkampii (all P > 0.05). Table 1.Comparison of clinical characteristics, antimicrobial susceptibility, and resistance genes between E. hormaechei, E. roggenkampii, and other Enterobacter spp. E. hormaechei (EH)E. roggenkampii (ER)Other Enterobacter spp.P - value (EH v.s ER)TotalNumber (%)15 (36.6)15 (36.6)11 (26.8) 41 (100)Year 2010–20146 (40)3 (20)4 (36.4)0.21313 (31.7) 2016–20209 (60)12 (80)7 (63.6) 28 (68.3)Age group 0.655 Children2 (13.3)1 (6.7)0 (0) 3 (7.3) Adult6 (40)6 (40)5 (45.5) 17 (41.5) Elderly6 (40)8 (53.3)6 (54.5) 20 (48.8) Unknown1 (6.7)0 (0)0 (0) 1 (2.4)Hospital type 0.5 Medical centre8 (53.3)7 (46.7)4 (26.7) 19 (46.3) Regional hospital7 (46.7)8 (53.3)7 (46.7) 22 (53.7)Region 0.064 Central1 (6.7)6 (40)4 (36.4) 11 (26.8) Eastern2 (13.3)0 (0)1 (9.1) 3 (7.3) Northern5 (33.3)6 (40)1 (9.1) 12 (29.3) Southern7 (46.7)3 (20)5 (45.5) 15 (36.6)Source 0.251 Blood4 (26.7)7 (46.7)5 (45.5) 16 (39) Pus2 (13.3)3 (20)3 (27.2) 8 (19.5) Respiratory3 (20)3 (20)0 (0) 6 (14.6) Urine6 (40)1 (6.7)2 (18.2) 9 (22.0) Other1 (6.7)1 (6.7)1 (9.1) 3 (7.3)Invasive3 (20)8 (53.3)5 (45.5)0.06416 (39)Location 0.648 ICU4 (26.7)2 (13.3)0 (0) 6 (14.6) Non-ICU inpatient9 (60)11 (73.3)11 (100) 31 (75.6) Outpatient (OPD and ER)2 (13.3)2 (13.3)0 (0) 4 (9.8)Antimicrobial non-susceptibility Amikacin3 (20)0 (0)0 (0)0.1123 (7.3) Gentamicin9 (60)0 (0)2 (18.2)<0.00111 (26.8) Aztreonam13 (86.7)4 (26.7)3 (27.3)0.00120 (48.8) Cefotaxime14 (93.3)4 (26.7)6 (54.5)<0.00124 (58.5) Cefepime12 (80)0 (0)1 (9.1)<0.00113 (31.7) Ciprofloxacin12 (80)5 (33.3)2 (18.2)0.01319 (46.3) Trimethoprim/sulfamethoxazole9 (60)0 (0)2 (18.2)<0.00111 (26.8) Piperacillin/ tazobactam14 (93.3)3 (20)1 (9.1)<0.00118 (43.9) Tigecycline10 (66.7)1 (6.7)0 (0)0.00111 (26.8) Colistina0 (0)12 (80)5 (45.4)<0.00117 (41.5) Meropenem/ vaborbactam0 (0)0 (0)0 (0)-0 (0) Imipenem/ relebactam12 (80)0 (0)4 (36.4)<0.00116 (39) Ceftazidime/ avibactam12 (80)0 (0)2 (18.2)<0.00114 (34.1)Antimicrobial resistance gene ESBL or AmpC gene carried on plasmid10 (66.7)0 (0)2 (18.2)<0.00112 (29.3) Carbapenemase geneb12 (80)0 (0)3 (27.3)<0.00115 (36.6) Chromosomal arn gene9 (60)15 (100)11 (100)0.01735 (85.4)mcr genec8 (53.3)3 (20)3 (27.3)0.06414 (34.1)^a^Colistin MIC was done by broth microdilution.^b^Carbapenemase genes included blaIMP-8 (n = 11) and blaVIM-1 (n = 1) in E. hormaechei, blaIMP-8 in E. kobei and E. asburiae, and blaIMI-1 in E. bugandensis. No carbapenemase genes were detected in the two E. cloacae isolates.^c^All three E. roggenkampii isolates harboured the mcr-10 gene, and all eight E. hormaechei isolates carried mcr-9. Among the remaining Enterobacter species, one E. kobei co-harboured mcr-9 and mcr-10, another E. kobei harboured mcr-10 only, and one E. asburiae harboured mcr-9 only.
Compared with E. roggenkampii, E. hormaechei isolates showed significantly higher resistance to aztreonam, cefotaxime, cefepime, ciprofloxacin, trimethoprim–sulfamethoxazole, tigecycline, and piperacillin/tazobactam (all P ≤ 0.025; ORs 8.0–120.0). Notably, 80% of E. hormaechei isolates were resistant to imipenem/relebactam or ceftazidime/avibactam, whereas none of the E. roggenkampii isolates showed resistance to these agents (P < 0.001). In contrast, colistin resistance was markedly more prevalent in E. roggenkampii than in E. hormaechei (80% vs. 0%, P < 0.001).
Genotypically, the chromosomal arn operon was universally present in all Enterobacter isolates except for six E. hormaechei isolates. Plasmid-mediated ESBL/AmpC and carbapenemase genes were confined mostly to E. hormaechei (80%). Although mcr genes were more frequently detected in E. hormaechei (53.3% vs 20.0%), this difference did not reach statistical significance (P = 0.128). The overall mcr carriage rate across Enterobacter isolates declined from 40–50% before 2018 to 9.1% by 2020 (Figure 1b).
Analysis of virulence-associated genes revealed that most isolates carried conserved adhesion – and biofilm-related determinants, including fimH, ompA, ompC, and the curli operon (csgA/csgG/csgD) (Supplementary Table S1).
Chromosomal mechanisms of colistin resistance
Colistin MICs showed no consistent correlation with mcr-9/mcr-10 carriage but were strongly associated with the chromosomal arnBCADTEF operon (Table 1). Among the 41 isolates, 35 (85.4%) carried the arn operon, while six E. hormaechei lacked it (Supplementary Table S1). All arn-negative strains remained susceptible (MIC ≤ 0.25–1 µg/mL), whereas arn-positive isolates exhibited variable MICs ranging from ≤0.25 to >4 µg/mL. All isolates retained intact mgrB and phoPQ genes without truncations, indicating these were not primary drivers of resistance.
Table 2 and Supplementary Figure S2 summarize colistin susceptibility testing results and inducible resistance phenotypes among representative isolates with different arn operon statuses and genetic backgrounds. Isolates lacking the arnBCADTEF operon, including E. hormaechei subsp. xiangfangensis (ST527) and subsp. hoffmannii (ST78), consistently exhibited low colistin MICs (≤0.125–0.25 µg/mL), showed no skipped-well phenomena, and failed to grow on colistin-containing agar (2 µg/mL), indicating stable susceptibility. In contrast, multiple arn-positive isolates, particularly E. hormaechei subsp. steigerwaltii ST90 and E. roggenkampii ST501, demonstrated discordant susceptibility profiles, characteristic skipped-well phenomena in broth macrodilution assays and growth on colistin-containing agar, consistent with inducible colistin resistance. Table 2.Colistin susceptibility and inducible resistance among Enterobacter spp. (extracted from Supplementary Table S1 and Fig S3).IsolateEnterobacter speciesMLSTColistin MIC range (µg/mL)amcr genesarn operonPromoter of arn operonbSkip wellColistin MHA 2 µg/mLInterpretationCRE097E. roggenkampiiST5010.25∼> 4−+Type 1+Colony growthInducible resistanceCRE078E. roggenkampiiST501> 4−+Type 1−Colony growthPersistent resistanceCRE001E. hormaechei subsp. xiangfangensisST527≤ 0.125−−NA−−No resistanceCRE074E. hormaechei subsp. xiangfangensisST527≤ 0.125∼0.25−−NA−−No resistanceCRE003E. hormaechei subsp. steigerwaltiiST900.125∼1++Type 2−Colony growthInducible resistanceCRE092E. hormaechei subsp. steigerwaltiiST900.125∼4++Type 2+Colony growthInducible resistanceCRE048E. hormaechei subsp. steigerwaltiiST90≤ 0.125∼2++Type 2+Colony growthInducible resistanceCRE068E. hormaechei subsp. steigerwaltiiST90≤ 0.125−+Type 2+Colony growthInducible resistanceCRE026E. hormaechei subsp. hoffmanniiST78≤ 0.125∼0.25+−NA−−No resistanceCRE064E. hormaechei subsp. hoffmanniiST78≤ 0.125+−NA−−No resistanceCRE075E. hormaechei subsp. hoffmanniiST78≤ 0.125−−NA−−No resistanceCRE033E. hormaechei subsp. steigerwaltiiST133≤ 0.125−−NA−−No resistanceCRE084E. hormaechei subsp. steigerwaltiiST133≤ 0.125−+Other−−No resistance+, present; −, absent; NA, not applicable.^a^The MIC range was determined in triplicate using the broth macrodilution method.^b^Type 1 promoter: attggtttaaatgtttccatttcaaaatgttgcggaagatcacatc; Type 2 promoter: tttattggaaatggtcggggatttttttatttgttga; Other promoter details are described in Supplementary Table S1.
Promoter type distribution differed by species. Type 1 promoters, predominant in E. roggenkampii, were associated with constitutive or inducible resistance, whereas type 2 promoters, common among E. hormaechei ST90 isolates, were linked specifically to inducible resistance. One exceptional E. hormaechei isolate (CRE084) harboured a divergent promoter variant, likely explaining the absence of inducible resistance despite an intact arn operon. Overall, colistin resistance in these Enterobacter isolates was primarily determined by chromosomal arn operon regulation with species differences.
Frequent co-carriage of blaIMP-8 and mcr-9 on IncHI2 plasmids
Co-carriage of blaIMP-8 and mcr-9 on IncHI2 plasmids persisted for 10 years in this study, representing a long-term stable multidrug resistance platform (Table 3). Overall, 36.6% (15/41) of Enterobacter isolates produced carbapenemases, the vast majority carrying blaIMP-8 (n = 13), primarily in E. hormaechei (n = 11), and in one isolate each of E. kobei and E. asburiae. Other carbapenemases included blaIMI-1, identified in a single isolate of E. bugandensis, and blaVIM-1, detected in one isolate of E. hormaechei. These IncHI2 plasmids were frequently co-localized with mcr-9.1 (7/8, 87.5%) and additional β-lactamases such as blaSHV-12 and blaTEM-1 (Table 3). E. hormaechei ST204 isolates exemplified this archetypal superplasmid, consistently carrying blaIMP-8, mcr-9.1, and additional β-lactamases on the same IncHI2 plasmid. BRIG analysis confirmed a highly conserved IncHI2 ST1 backbone across isolates from different years and regions (2010–2020) (Supplementary Figure S3A). Despite the frequent presence of mcr genes, most blaIMP-8–positive isolates remained colistin-susceptible. Table 3.Characteristics of blaIMP-8-carrying Enterobacter spp.SpeciesIsolateYearRegionMLSTSizes of plasmids or inserts (bp)Location of blaIMP-8 genesCo-harbouring ß-lactamasemcr genearn cassetteE. hormaechei subsp. steigerwaltiiCRE0052010NST204326857IncHI2 ST1SHV-12, TEM-1mcr-9.1+E. hormaechei subsp. steigerwaltiiCRE0272014NST204318009IncHI2 ST1SHV-12, TEM-1mcr-9.1+E. hormaechei subsp. steigerwaltiiCRE0652018SST204313832IncHI2 ST1SHV-12, TEM-1mcr-9.1+E. hormaechei subsp. steigerwaltiiCRE0332016SST133161895FIIYTEM-1NA−E. hormaechei subsp. steigerwaltiiCRE0842020SST133143296FIIYNANA+E. hormaechei subsp. steigerwaltiiCRE0482016NST9080734IncHI2 ST1NAmcr-9.1+E. hormaechei subsp. steigerwaltiiCRE0682018NST90154455FIIYNANA+E. hormaechei subsp. steigerwaltiiCRE0922020SST90187175FIIYaTEM-1mcr-9.1a+E. hormaechei subsp. hofmanniiCRE0262014SST78283759IncHI2 ST1SHV-12, TEM-1mcr-9.1−E. hormaechei subsp. hofmanniiCRE0642018SST78258855IncHI2 ST1TEM-1mcr-9.1−E. hormaechei subsp. hofmanniiCRE0752018CST7880458No matchNANA−E. asburiaeCRE0472016NST271278171IncHI2 ST1NAmcr-9.1+E. kobeiCRE0402016SST25299622IncHI2 ST1TEM-1mcr-9.1, mcr-10 b++, present; –, absent; NA, not applicable.^a^In this isolate, mcr-9.1 and blaIMP-8 were located on different plasmids: mcr-9.1 on IncHI2 and blaIMP-8 on IncFIIY plasmid.^b^In this isolate, blaIMP-8 and mcr-9.1 co-localized on the same IncHI2 plasmid, whereas mcr-10 was carried on a separate plasmid.
In contrast, IncFIIY plasmids (4/13) carrying blaIMP-8 rarely co-localized with mcr genes, forming a distinct genomic profile. These IncFIIY plasmids showed greater variability in resistance loci and mobile genetic elements (Supplementary Figure S3B).
Structural conservation of the blaIMP–mcr-9 superplasmid
Comparative genomic analysis revealed that all blaIMP-8 was consistently embedded within a highly conserved class 1 integron (intI1–blaIMP-8–aac(6”)-Ib–catB3–qacEΔ1–sul1± ISCR1), irrespective of whether the gene resided on IncHI2 or IncFIIY plasmids (Figure 3). This integron was characterized by the presence of the intI1 integrase at the 5’ conserved segment (5’ CS), followed by a resistance gene cassette comprising blaIMP-8–aac(6’)-Ib–catB3, and ending with a 3’ conserved segment (3’CS) containing qacEΔ1–sul1. In most cases, the cassette was followed by an Insertion Sequence Common Region 1 (ISCR1) element, forming a typical complex class 1 integron frequently associated with multidrug resistance. This integron therefore represents the fundamental transmissible unit, while plasmid backbones provided distinct dissemination platforms. Figure 3.Comparative genomic analysis of the genetic environments of mcr-9 and blaIMP-8 among Taiwan Enterobacter isolates, 2010–2020. Each horizontal block represents a genomic region containing either the mcr or blaIMP gene. Arrows indicate open reading frames coloured by functional annotation. Grey shading between sequences indicates regions of shared homology among different plasmids ranging from 80% to 100%. Red: mcr-9 gene and surrounding genes; Yellow: blaIMP-8 gene and associated integron elements.
Global dissemination of blaIMP–mcr-9 superplasmids
To place the Taiwan findings in a broader context, we compared these IncHI2 superplasmids with international isolates, demonstrating a globally conserved blaIMP–mcr-9 resistance module (Figure 4 and Supplementary Table S3). Across Taiwan, the UK (CP043767), and China (MH829594, MH399264), the mcr-9 locus was consistently embedded within the IS903B–mcr-9–wbuC–pco–rcn cluster, linking mcr-9 gene with copper and arsenic resistance determinants and suggesting co-selection by environmental pressures. This mcr-9 module was frequently linked to a conserved class 1 integron carrying blaIMP and together they formed a composite multidrug resistance region of ∼44–60 kb bracketed by IS26 elements. Figure 4.Comparative genomic analysis of global IncHI2 plasmids co-harbouring mcr-9 and carbapenemase genes in Enterobacter spp. Linear representations of plasmid regions are shown with open reading frames (ORFs) depicted as arrows, colour-coded by predicted function: red indicates antibiotic resistance genes (ARGs), purple represents heavy metal resistance genes (MRGs), blue denotes insertion sequences (IS26), green for IS903B elements, black/grey for transposases and hypothetical proteins, and uncoloured/white for other coding sequences. Homologous regions with ≥90% nucleotide identity was shaded in grey, with pink connectors indicating shared regions across plasmids.
Detection of mcr-10 and Associated Plasmid Structures
Five isolates (12.2%) carried mcr-10, including E. roggenkampii (n = 3) and E. kobei (n = 2). As with mcr-9, mcr-10 carriage did not associate with elevated colistin MICs in our collection. All mcr-10 genes were located on IncF plasmids, which exhibited heterogeneous architectures with no closely related international counterparts in our comparisons (data not shown).
Discussion
This nationwide surveillance study revealed that, colistin resistance among carbapenem-non-susceptible Enterobacter isolates in Taiwan was primarily mediated by the chromosomal arn operon, which was present in 85.4% of isolates. In contrast, carriage of mcr genes had little effect on the resistance phenotype. A particularly concerning finding was inducible colistin resistance mediated by the arn operon, a phenotype easily missed by routine antimicrobial susceptibility testing. Together, these findings challenge the long-standing assumption that Enterobacter species are uniformly susceptible to colistin and underscore the need to re-evaluate the role of colistin in the treatment of infections caused by carbapenem-non-susceptible Enterobacter.
From a clinical microbiology perspective, the predominance of chromosomally mediated, inducible colistin resistance has profound implications for routine antimicrobial susceptibility testing. In Enterobacter, exposure to colistin can readily activate the chromosomal arn operon, resulting in heterogeneous resistance phenotypes that are difficult to detect and that substantially increase the risk of inappropriate therapy. Although CLSI currently categorizes Enterobacter species as intrinsically susceptible to colistin [14], resistance rates of 4–20% have been reported in multiple countries [30], suggesting systematic underestimation of true resistance. Taken together, these findings indicate that Enterobacter species may be more appropriately regarded as functionally intrinsically resistant to colistin in clinical practice. Accordingly, colistin may be suboptimal for the treatment of Enterobacter infections and should be used with caution even when in vitro susceptibility is reported, as exposure may rapidly select for resistance and increase the risk of therapeutic failure [11,12]. Awareness of chromosomal resistance mechanisms and genomic context is therefore essential to guide appropriate antimicrobial decision-making.
Our species-level analysis demonstrated marked heterogeneity in colistin resistance among Enterobacter species, consistent with previous hsp60-based studies [9,31]. The arn operon was universally present in non–E. hormaechei species but absent in E. hormaechei hsp60 clusters III (subsp. hoffmannii) and VI (subsp. xiangfangensis) [9]. In contrast, arn carriage within E. hormaechei cluster VIII (subsp. steigerwaltii) was heterogeneous, occurring even among isolates sharing the same sequence type (ST133) in the present study. This finding contrasts with a German study reporting uniform arn absence in ST133 and suggests regional variation in the genetic composition of these strains [9].
The long-term circulation of a conserved IncHI2 “superplasmid” carrying blaIMP-8 and mcr-9 from 2010 to 2020 in Taiwan indicates its stability and dissemination, both clonally within E. hormaechei ST204 and ST78 and across other Enterobacter species, including E. asburiae and E. kobei. Similar IncHI2 plasmids co-harbouring mcr-9 and carbapenemases like blaIMP or blaNDM have been reported globally [32–34]. The IncHI2 superplasmid has also been identified in other Enterobacterales, such as K. pneumoniae, E. coli, and Salmonella [35,36], and is known to be transferable via conjugation [35,37]. Importantly, our findings suggest that complex integrons, particularly those mediated by ISCR1, rather than the plasmid backbones themselves, constitute the fundamental transferable units sustaining the dissemination of blaIMP-8 [38]. Although blaNDM was not detected among Enterobacter isolates during the study period, NDM-producing Enterobacter have been reported in India and China [34,37]. Notably, a recent outbreak reported in 2025 documented the emergence of blaNDM-harbouring Enterobacter in a southern Taiwan hospital, indicating that blaNDM has now entered Taiwan and underscoring the need for continued genomic surveillance [39].
This study marks the detection of mcr-10 in Enterobacter species in Taiwan. Unlike mcr-9, mcr-10 was located on IncFIB/FII plasmids and not associated with carbapenemase genes. This finding is consistent with a report from China [40]. Although mcr-10 did not confer phenotypic resistance on its own, its presence in various animal and environmental reservoirs highlights the importance of a One Health framework that integrates human, animal, and environmental surveillance [41,42]. This illustrates how novel resistance genes can persist and gain significance under selective pressures, even without an immediate clinical impact.
Despite being the most comprehensive genomic dataset of carbapenem-non-susceptible Enterobacter in Taiwan to date, this study has limitations. Only 41 isolates were sequenced, which likely underrepresents the true circulating diversity. The lack of functional validation experiments prevents definitive genotype-phenotype confirmation, and clinical outcome data were unavailable. Furthermore, while population analysis profiling (PAP) is the gold standard for heteroresistance, our assays served only as a screening tool and may have underestimated resistant subpopulations.
In conclusion, the convergence of carbapenem non-susceptibility and colistin resistance renders Enterobacter a high-risk multidrug-resistant organism. This threat is further compounded by inducible colistin resistance – often undetectable by standard antimicrobial susceptibility testing – and by the long-term persistence of multidrug-resistant plasmids. These findings underscore the urgent need to re-evaluate current therapeutic strategies and to strengthen measures against the spread of these difficult-to-treat resistance mechanisms.
Supplementary Material
0113 Supplement.xlsx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Davin-Regli A, Lavigne JP, Pages JM. Enterobacter spp.: update on taxonomy, clinical aspects, and emerging antimicrobial resistance. Clin Microbiol Rev. 2019;32:e 00002–e 00019. doi: 10.1128/CMR.00002-1931315895 PMC 6750132 · doi ↗ · pubmed ↗
- 2Huang YC, Kuo SC, Fang CT, et al. Changing epidemiology and antimicrobial resistance of bacteria causing bacteremia in Taiwan: 2002-2020. Microbiol Spectr. 2024;12:e 0060824. doi: 10.1128/spectrum.00608-2438916365 PMC 11301998 · doi ↗ · pubmed ↗
- 3Wang JT, Wu UI, Lauderdale TL, et al. Carbapenem-nonsusceptible Enterobacteriaceae in Taiwan. P Lo S One. 2015 Mar 20;10:e 0121668. doi: 10.1371/journal.pone.012166825794144 PMC 4368706 · doi ↗ · pubmed ↗
- 4Alvarez-Marin R, Navarro-Amuedo D, Gasch-Blasi O, et al. A prospective, multicenter case control study of risk factors for acquisition and mortality in Enterobacter species bacteremia. J Infect. 2020;80:174–181. doi: 10.1016/j.jinf.2019.09.01731585192 · doi ↗ · pubmed ↗
- 5Annavajhala MK, Gomez-Simmonds A, Uhlemann AC. Multidrug-Resistant Enterobacter cloacae complex emerging as a global, diversifying threat. Front Microbiol. 2019;10:44. doi: 10.3389/fmicb.2019.0004430766518 PMC 6365427 · doi ↗ · pubmed ↗
- 6Doumith M, Ellington MJ, Livermore DM, et al. Molecular mechanisms disrupting porin expression in ertapenem-resistant Klebsiella and Enterobacter spp. clinical isolates from the UK. J Antimicrob Chemother. 2009;63:659–667. doi: 10.1093/jac/dkp 02919233898 · doi ↗ · pubmed ↗
- 7Yeh TK, Lin HJ, Liu PY, et al. Antibiotic resistance in Enterobacter hormaechei. Int J Antimicrob Agents. 2022;60:106650. doi: 10.1016/j.ijantimicag.2022.10665035934231 · doi ↗ · pubmed ↗
- 8Hong YK, Ko KS. Pmr AB and Pho PQ variants in colistin-resistant enterobacter spp. isolates in Korea. Curr Microbiol. 2019;76:644–649. doi: 10.1007/s 00284-019-01672-130891622 · doi ↗ · pubmed ↗