Divergent understandings in comparative oncology
Zachary T. Compton, Amy M. Boddy, Lisa M. Abegglen, Daniel Chávez, Joshua D. Schiffman, Marc Tollis, Carlo C. Maley

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
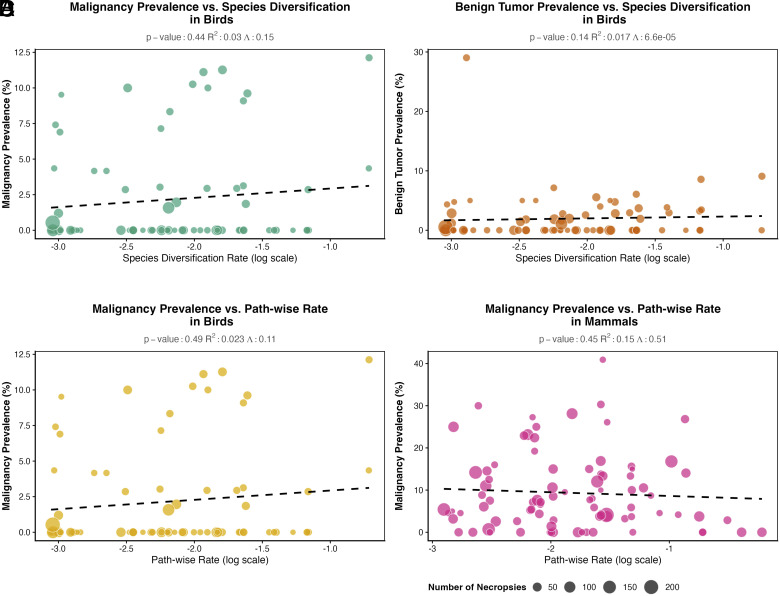 Figure 1
Figure 1- —HHS | NIH | NCI | CCR | Basic Research Laboratory (BRL)100016229
- —HHS | NIH | NCI | CCR | Basic Research Laboratory (BRL)100016229
- —HHS | NIH | Advanced Research Projects Agency for Health (ARPA-H)100023015
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer Genomics and Diagnostics · Advances in Oncology and Radiotherapy · Global Cancer Incidence and Screening
Butler et al.’s recent exploration of the rates of species diversification and cancer is an important addition to the study of comparative oncology (1). Do lineages that tend to diversify more rapidly get more cancer? The authors hypothesize that a susceptibility to genomic rearrangements might lead to higher diversification and cancer rates. However, because deleterious rearrangements are swiftly eliminated, only lineages with robust, stabilized genomes can diversify (2). Diversification is a signature of genomic stability rather than instability. Therefore, species with more stable genomes might diversify more and have less cancer. Unfortunately, the statistical model, previously published in Butler et al.’s analysis of body size and cancer (3), is deeply flawed. Butler et al. treat tumor counts as additive, therefore, any apparent relationship between diversification and cancer prevalence may simply reflect differences in sample size rather than related to evolution. Their approach estimates a proportion of necropies that will have cancer, which implies that all species have the same rate of cancer. The species differences only come through their differences in the other predictors (body size, change in body size, and diversification rate) which add or subtract a fixed number of cancers, regardless of the number of necropsies observed. With only 87 mammal and 76 bird species in their analysis, Butler et al. may lack the statistical power needed to detect interactions between the number of necropsies and their other predictors (they do not report testing for those interactions). An alternative statistical model allows each species to have a different intrinsic rate of cancer which is then modified by the other predictors like body size or diversification rate. When we reanalyzed their data under these assumptions using methods from Compton et al. (4), we found no evidence of an association between diversification rate and either malignancy or benign tumor prevalence in either mammals or birds (Fig. 1). Nor did we find an association between change in body size and malignancy in either birds or mammals (Fig. 1). While a larger sample size may prove such a relationship in the future, it is unlikely to rescue fundamentally flawed methodology.
While Butler et al.’s analysis of the rate of change in body size is intriguing, we predict that the direction of that change is important (5). Their analysis fails to distinguish between lineages that have recently shrunk versus recently grown. We predict these scenarios have opposite implications: shrinking lineages inherit robust cancer suppression from larger ancestors, while growing lineages could remain vulnerable to cancer until selection generates mechanisms appropriate to their new size (6). We appreciate that our colleagues have utilized the cancer prevalence data we published in innovative ways, but we are acutely aware of the need for many more data points to detect subtle effects and interactions between factors.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1G. Butler, J. Baker, S. R. Amend, K. J. Pienta, C. Venditti, Divergent evolutionary dynamics of benign and malignant tumors. Proc. Natl. Acad. Sci. U.S.A. 122, e 2519203122 (2025).41196351 10.1073/pnas.2519203122 PMC 12625976 · doi ↗ · pubmed ↗
- 2D. L. Rabosky , Rates of speciation and morphological evolution are correlated across the largest vertebrate radiation. Nat. Commun. 4, 1958 (2013).23739623 10.1038/ncomms 2958 · doi ↗ · pubmed ↗
- 3G. Butler, J. Baker, S. R. Amend, K. J. Pienta, C. Venditti, No evidence for Peto’s paradox in terrestrial vertebrates. Proc. Natl. Acad. Sci. U.S.A. 122, e 2422861122 (2025).39993196 10.1073/pnas.2422861122 PMC 11892590 · doi ↗ · pubmed ↗
- 4Z. T. Compton , Cancer prevalence across vertebrates. Cancer Discov. 15, 227–244 (2025).39445720 10.1158/2159-8290.CD-24-0573 PMC 11726020 · doi ↗ · pubmed ↗
- 5Z. T. Compton , Paradoxical indeed. Proc. Natl. Acad. Sci. U.S.A. 122, e 2504512122 (2025).40324092 10.1073/pnas.2504512122 PMC 12088420 · doi ↗ · pubmed ↗
- 6E. Y. Erten, M. Tollis, H. Kokko, Shrinking to bird size with dinosaur-level cancer defences: Evolution of cancer suppression over macroevolutionary time. P Lo S Comput. Biol. 21, e 1013432 (2025).40924809 10.1371/journal.pcbi.1013432 PMC 12435733 · doi ↗ · pubmed ↗