Assessment of Leishmania exposure in blood donors from a non‐endemic urban area: A study in São Paulo
Ismael Severino de Lima, Suzete Cleusa Ferreira, Anna Shoko Nishiya, Norival Kesper, Jerenice Esdras Ferreira, Claudia Maria de Castro Gomes, Juliana Derriga, Katia Cristina Dantas, Silvia Petrossi Gallo Polato, Nanci Alves Salles, Jose Angelo Lauletta Lindoso, Tila Fanciani

TL;DR
This study found no active Leishmania infections in blood donors from São Paulo, a non-endemic area, despite some positive antibody tests, suggesting limitations in current screening methods.
Contribution
The study evaluates the reliability of serological screening for Leishmania in non-endemic regions and recommends molecular methods for improved transfusion safety.
Findings
2.82% of blood donors had ELISA-reactive results, but none tested positive by PCR for Leishmania DNA.
Non-specific reactivity in serological tests may be due to inflammatory conditions like pregnancy or abortion.
Molecular methods and universal leukoreduction are recommended to improve transfusion safety in low-endemicity areas.
Abstract
According to the World Health Organization, more than 1 billion people are at risk of leishmaniasis in over 89 countries. Environmental changes such as deforestation, urban expansion and climate change facilitate the spread of sand fly vectors and reservoirs, increasing disease transmission. The introduction of Leishmania into non‐endemic regions may be further driven by globalization and ecotourism. Transfusion transmission, particularly of Leishmania infantum, remains a concern due to the parasite's ability to survive under blood storage conditions and its prolonged latent phase. We aimed to determine the prevalence of Leishmania spp. among blood donors in a non‐endemic region. A prospective, cross‐sectional study was conducted with 5145 blood donor samples collected from January to December 2023. Serological screening was performed using an in‐house immunoglobulin G (IgG) ELISA…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
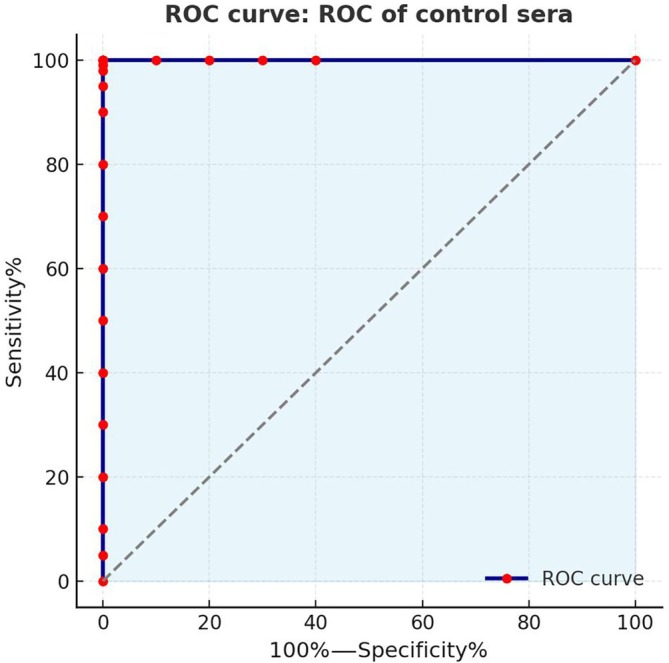 FIGURE 1
FIGURE 1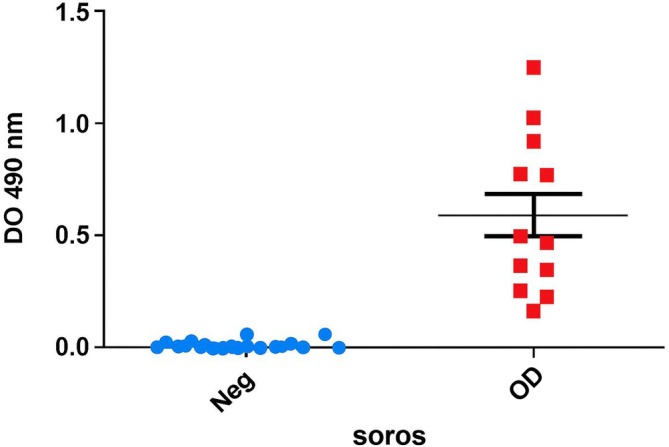 FIGURE 2
FIGURE 2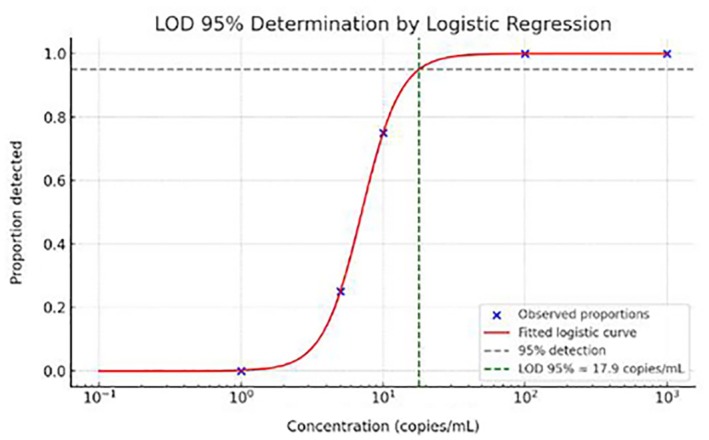 FIGURE 3
FIGURE 3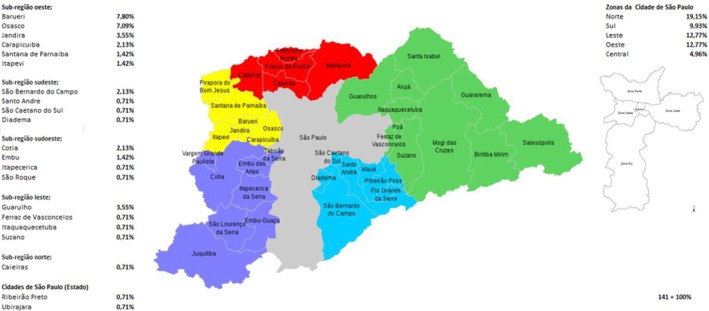 FIGURE 4
FIGURE 4| Variable |
| % |
| % |
|
|---|---|---|---|---|---|
| Age (years) | 0.4757 | ||||
| <38 | 72 | 51.10 | 2432 | 48.60 | |
| 39–58 | 63 | 44.70 | 2232 | 44.60 | |
| >59 | 6 | 4.20 | 340 | 6.80 | |
| Sex | 0.6862 | ||||
| Female | 70 | 49.60 | 2398 | 47.90 | |
| Male | 71 | 50.40 | 2606 | 52.10 | |
| Education level | 0.9783 | ||||
| R1—Never attended school | 0 | 0.00 | 3 | 0.06 | |
| R2A—Incomplete elementary | 4 | 2.80 | 113 | 2.25 | |
| R3—Complete elementary | 7 | 5.00 | 221 | 4.40 | |
| R4—High school | 56 | 39.70 | 1980 | 39.60 | |
| R5—Technical course | 2 | 1.40 | 145 | 2.90 | |
| R6—Higher education | 70 | 49.70 | 2475 | 49.50 | |
| R7—Master's degree | 2 | 1.40 | 62 | 1.20 | |
| R8—Doctorate degree | 0 | 0.00 | 5 | 0.09 | |
| Race/ethnicity | 0.6562 | ||||
| Black | 10 | 7.10 | 388 | 7.70 | |
| Mixed (Parda) | 34 | 24.10 | 1464 | 29.30 | |
| White | 94 | 66.70 | 3029 | 60.50 | |
| Asian (Yellow) | 3 | 2.10 | 114 | 2.30 | |
| Indigenous | 0 | 0.00 | 9 | 0.20 | |
| Donor type | 0.7041 | ||||
| First‐time | 48 | 34.00 | 1556 | 31.10 | |
| Repeat | 50 | 35.50 | 1783 | 35.60 | |
| Sporadic | 43 | 30.50 | 1665 | 33.30 |
| TESA‐blot result |
| % |
|---|---|---|
| Positive | 13 | 23.6% |
| Negative | 36 | 65.4% |
| Doubtful | 6 | 10.9% |
| Total | 55 | 100.0% |
| Concentration (copies/mL) | No. of replicates | No. of positives |
|---|---|---|
| 1000 | 20 | 20 |
| 100 | 20 | 20 |
| 10 | 20 | 15 |
| 5 | 20 | 5 |
| 1 | 20 | 0 |
| 0 | 20 | 0 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsResearch on Leishmaniasis Studies · Trypanosoma species research and implications · Hemophilia Treatment and Research
Highlights
- The seroprevalence of Leishmania among blood donors in a non‐endemic area was 2.82%, but none of the samples were confirmed by PCR.
- The findings suggest that non‐specific ELISA reactivity may be associated with physiological conditions rather than true infection.
- The incorporation of complementary molecular methods and universal leukoreduction is recommended to enhance transfusion safety and avoid unnecessary donor deferrals.
INTRODUCTION
Leishmaniasis is a neglected tropical disease caused by protozoa of the genus Leishmania, transmitted primarily through the bite of infected female phlebotomine sandflies. The disease manifests mainly in two clinical forms: cutaneous and visceral leishmaniasis (VL), the latter being potentially fatal if left untreated. Although vectorial transmission is the most common route, non‐vectorial transmission including congenital, sexual, organ transplantation and blood transfusion has been increasingly recognized, raising significant concerns in transfusion medicine [1].
Leishmania parasites have been shown to persist in the peripheral blood of asymptomatic individuals, posing a silent threat to transfusion recipients, particularly immunocompromised patients. While the transfusional transmission of Leishmania is not well documented, evidence of parasite DNA in blood donors detected through molecular techniques supports the possibility of this transmission route [1, 2, 3].
In endemic areas, studies have reported substantial prevalence rates of asymptomatic infection among blood donors. For instance, in Mato Grosso do Sul, Brazil, Leishmania DNA was detected in 5.4% of healthy donors [1], and 2.8% in Araçatuba, São Paulo, a region with recent VL urbanization [2]. In Teresina, Piauí, another endemic city in Northeastern Brazil, Leishmania DNA was found in 3.6% of donors [3]. Similar findings have been observed internationally, with molecular detection rates of 2.8% in blood donors from Iran [4] and 4.2% seroprevalence in India [5].
Interestingly, several studies have also reported Leishmania infection in blood donors from non‐endemic regions. In Paraná, Southern Brazil, Leishmania DNA was found in 0.4% of donors despite the region being considered non‐endemic [6]. In Madrid, Spain, where VL is not widespread, molecular testing detected Leishmania infantum DNA in 0.88% of asymptomatic blood donors [7, 8, 9, 10, 11, 12]. Such findings highlight the potential role of silent carriers and the risk of transfusion‐transmitted leishmaniasis (TTL) even outside recognized endemic zones.
These findings reinforce the importance of active surveillance and the need for targeted screening strategies in endemic regions, particularly for transfusion recipients at high risk, such as immunocompromised individuals. However, a few studies report the prevalence of Leishmania spp. in non‐endemic areas. Therefore, the objective of our study was to determine the prevalence of Leishmania spp. among blood donors from a non‐endemic region.
MATERIALS AND METHODS
This was a prospective, cross‐sectional study that included 5145 blood donor samples collected from January to December 2023. Serological screening was performed using an in‐house immunoglobulin G (IgG) ELISA based on Leishmania chagasi culture antigen [13]. Samples with positive or inconclusive serological results were further tested by real‐time PCR targeting the internal transcribed spacer (ITS) and kinetoplast DNA (kDNA) regions, following the protocol described by Pirmez et al. as a confirmatory assay [14].
In‐house ELISA
An in‐house ELISA was performed to detect anti‐Leishmania antibodies. Polystyrene microplates (Costar High Binding 3690, Corning, NY, USA) were coated with L. chagasi antigen at a concentration of 0.5 μg/mL (50 μL/well) and blocked with 5% skim milk, as previously described [8, 9, 10]. Serum samples were diluted 1:200 and incubated in duplicate at 37°C for 60 min. After five washes with phosphate‐buffered saline (PBS)‐Tween buffer, 50 μL of anti‐human IgG peroxidase‐conjugated antibody (A0170, Sigma‐Aldrich, St. Louis, USA), diluted 1:30,000, was added. Plates were incubated again at 37°C for 60 min followed by additional washing. A chromogenic substrate solution was then added (50 μL/well) and incubated at 37°C for 30 min. The reaction was stopped by adding 1 N hydrochloric acid, and absorbance was read at 492 nm using a microplate reader [13].
TESA‐blot immunoblotting for Trypanosoma cruzi
To exclude Trypanosoma cruzi infection, the trypomastigote excreted‐secreted antigen (TESA)‐blot immunoblotting assay was performed following Umezawa et al. [15]. Nitrocellulose membranes containing TESA were blocked with PBS containing 5% skim milk and incubated for 1 h at room temperature (RT) with constant agitation. The membranes were then incubated with serum samples diluted 1:100 in PBS with 1% skim milk for 2 h at RT, followed by five washes with PBS. Next, the membranes were incubated with peroxidase‐conjugated anti‐human IgG (1:4000, Sigma‐Aldrich, USA) for 2 h at RT. After another series of PBS washes, immune complexes were visualized by adding a chromogenic solution consisting of 6 mg of 4‐chloro‐1‐naphthol, 2 mL methanol, 10 mL PBS and 10 μL H_2_O_2_. The reaction was stopped with distilled water upon the appearance of reactive bands, and the membranes were dried between filter papers. A reactive band at 150–160 kDa was considered indicative of T. cruzi infection [16].
Real‐time PCR for Leishmania
A highly sensitive in‐house real‐time PCR multiplex assay targeting the ITS ribosomal RNA (rRNA) and kDNA minicircle regions of Leishmania spp. was used to quantify parasite load [16, 17, 18]. Total nucleic acids were extracted from 500 μL of whole blood using the MagNA Pure Compact Nucleic Acid Isolation Kit—Large Volume (Roche, Germany), with elution in 100 μL, or alternatively using the QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer's instructions. An internal control was added during extraction using 2 μL of poliovirus vaccine (diluted 1:100 in ultrapure water), which was detected by a specific real‐time PCR assay using the following primers and probe: PV_Forward: 5′‐CCCTCCCCTCACAAAAACAG‐3′, PV_Reverse: 5′‐TCACTTGCATGGAGTCTTGCA‐3′ and PV_Probe: 5′‐VIC‐TGGTACTGTTTCCTTGCC‐NFQ‐3′ [17]. For detection of Leishmania, amplification was performed using primers L.ITS.F (5′‐CAAATACACGCATGCACTCTC‐3′) and L.ITS.R (5′‐TTTAATAATCCTGGTCACAGCC‐3′), and the probe FAM‐5′‐AGCGTCGAAACTCCTCTCTGGTGC‐3′‐TAMRA. Additionally, primers HM1 (5′‐CCGCCCCTATTTTACACCAACCCC‐3′), HM2 (5′‐GGGGAGGGGCGTTCTGCGAA‐3′) and HM3 (5′‐GGCCCACTATATTACACCAACCCC‐3′) were used to amplify a 120 bp fragment of the conserved kDNA minicircle region of Leishmania [16, 18]. Amplification was carried out in a StepOne Plus Real‐Time PCR System (Applied Biosystems, Foster City, CA, USA) with the following cycling conditions: initial denaturation at 95°C for 10 min, followed by 45–50 cycles of 94°C for 15 s and 60°C for 60 s.
Statistical analysis
This was a prospective cross‐sectional study conducted with 5000 blood donor samples to estimate the prevalence of (disease/agent). Categorical variables were analysed using 2 × 2 contingency tables, with associations assessed by the chi‐square test or Fisher's exact test when expected cell counts were <5. Prevalence was calculated with a 95% confidence interval (CI). A p‐value <0.05 was considered statistically significant. All statistical analyses were performed using GraphPad Prism, version 9.5.1 (GraphPad Software, San Diego, CA, USA).
Statistical estimation of the limit of detection with 95% CI
The limit of detection (LOD) and its 95% CI were estimated using a binary logistic regression model, in accordance with the guidelines provided by the Clinical and Laboratory Standards Institute (CLSI) EP17‐A2 (2012) [19]. Serial dilutions of known target concentrations (expressed in copies/mL) were prepared, and 20 replicate reactions were tested at each concentration level. The proportion of positive results at each dilution was used to model the probability of detection as a function of analyte concentration.
The logistic regression equation applied was:
From the fitted model, the LOD—defined as the concentration at which there is a 95% probability of detection—was determined using the inverse of the logistic function:
The 95% confidence interval (CI) for the LOD was calculated using the delta method or profile likelihood estimation, as implemented in standard statistical software (e.g., R or Python's statsmodels), based on the standard errors of the estimated regression coefficients.
This method allows for robust estimation of the detection capability and its uncertainty, and is widely accepted for molecular diagnostic validations.
Ethics statement
Human subjects research was conducted following approval from the Institutional Review Board (CAPPesq) of Hospital das Clínicas, University of São Paulo (online registration CAAE# 62177822.8.0000.0068). All blood donors provided written informed consent before sample collection. For donors aged 16 or 17, signed consent was obtained from a parent or legal representative.
RESULTS
Reactivity threshold determination
The reactivity threshold (optical density—0.125) of the ELISA was established based on the reactivity of Leishmania alkaline extract (EAL) with 22 samples from healthy individuals (N) and 12 samples from patients with VL, including cases with high, moderate and low serological reactivity as determined by conventional serology. The cutoff was calculated using the receiver‐operating characteristic (ROC) curve method [19] in Prism™ software version 5.0 (GraphPad Software Inc., 1999) for Windows®. This analysis compared the Abs_492nm_ values of samples from healthy individuals with those from VL patients confirmed by parasitological and/or serological tests (Figure 1). The in‐house ELISA for Leishmania spp. demonstrated both 100% sensitivity and specificity (Figures 1 and 2) [19, 20].
Receiver‐operating characteristic (ROC) curve generated from control sera, showing that the in‐house ELISA for Leishmania achieved 100% sensitivity and specificity.
Reactivity results of the panel containing positive and negative controls tested by the in‐house ELISA using Leishmania chagasi alkaline extract antigen.
Among the 5145 donor samples tested, a seroprevalence of 2.74% (141/5, 145) for Leishmania spp. was observed. None of the ELISA‐reactive samples tested positive by real‐time PCR targeting the ITS and kDNA regions.
Sociodemographic data indicated that most reactive individuals were young (under 38 years old), self‐identified as White and had higher education levels; however, none of these variables were statistically significant (p > 0.05). Notably, among the 141 ELISA‐positive samples, 31 donors (22.0%) reported having given birth or having experienced a miscarriage, compared to only 6 (0.12%) among ELISA‐negative samples. This association was statistically significant (p < 0.0000001; Table 1).
We used the same panel of samples employed to determine the cutoff value of the in‐house ELISA for leishmaniasis to assess whether the Chagas Architect test (Abbott) would cross‐react with Leishmania spp. All samples that tested positive for Leishmania also showed reactivity in the Chagas Architect assay.
All samples that initially tested reactive for Chagas were excluded from in‐house Leishmania ELISA screening, as they could potentially yield false‐positive results due to cross‐reactivity. During the study period, 55 samples that tested positive for Chagas by the Architect assay were submitted to confirmatory testing using the TESA‐blot. Among these, 13 (23.6%) were confirmed positive, and 6 (10.9%) showed indeterminate results by TESA‐blot (Table 2).
ELISA positivity using epimastigote alkaline extract compared to TESA‐blot
Among the 36 samples (65.4%) that tested negative for Chagas disease by the TESA‐blot confirmatory assay, we performed the in‐house Leishmania ELISA and real‐time PCR. All results were negative for both tests.
The LOD at 95% CI for the real‐time PCR assay was estimated at approximately 17.9 copies/mL, based on a logistic regression model. The graph (Figure 3) shows the logistic curve fitted to the experimental data, indicating the intersection point with the 95% detection threshold (Table 3).
Logistic regression model used to determine the 95% limit of detection (LOD) for the real‐time PCR assay targeting Leishmania spp. The curve shows the probability of detection as a function of parasite concentration (copies/mL). The dashed line indicates the 95% detection probability threshold, intersecting the curve at approximately 17.9 copies/mL, representing the assay's LOD with 95% confidence interval.
This approach is commonly used to define the analytical sensitivity of molecular assays, as described by Armbruster and Pry [16, 17] and other authors who apply logistic curve modelling to estimate LOD in molecular diagnostics (Table 3 and Figure 3) [16, 17].
When analysing the distribution of seropositive blood donors by residential region, we observed the highest prevalence in the Northern region (19.15%), followed by the Eastern (12.77%), Western (12.77%), Southern (9.93%) and Central (4.96%) regions (Figure 4).
Percentage distribution of the number of cases by region in the State of São Paulo.
DISCUSSION
According to the World Health Organization, more than 1 billion people are at risk of contracting leishmaniasis across over 89 countries [21]. Deforestation, unplanned urbanization and the replacement of natural forests with plantations or human dwellings bring people closer to the natural habitats of sand fly vectors and wild reservoirs, thus favouring disease transmission. Other important factors contributing to the spread of leishmaniasis to non‐endemic areas include globalization, climate change and ecotourism [22].
Transfusion‐transmitted Leishmania infection has become an increasing concern, particularly in regions endemic for VL, due to the potential for a prolonged latent phase and the parasite's intracellular localization. In this context, universal leukoreduction has been suggested as an effective strategy to reduce transfusion transmission risk, since Leishmania primarily resides in leukocytes [23].
In our study, we observed a seroprevalence of 2.74% (141/5145) for Leishmania spp. enzyme immunoassay‐reactivity among blood donors from São Paulo, a region considered non‐endemic for VL. However, none of the ELISA‐reactive samples tested positive by real‐time PCR. A comparable investigation carried out in the non‐endemic city of Salvador, Northeastern Brazil, found an anti‐Leishmania seroprevalence of 5.4% (38/700) by ELISA, yet none of these seropositive samples demonstrated the presence of parasite DNA by qPCR targeting glucose‐6‐phosphate dehydrogenase, ITS or kDNA regions. This similarity underscores the recurrent issue of serological reactivity without molecular confirmation in low‐endemic settings and reinforces the need for confirmatory testing protocols to ensure transfusion safety and prevent unnecessary donor deferrals [22].
Although ELISA is widely used due to its high sensitivity and ease of implementation, false‐positive results are well‐documented in low‐endemicity settings. In our study, 31 (44.3%) of the 70 women's seroreactive samples were from women who were either pregnant or had a recent history of abortion. Both pregnancy and abortion are associated with systemic inflammatory responses and immune modulation, which can lead to polyclonal B‐cell activation and increased production of non‐specific antibodies, potentially resulting in false‐positive serological results [23]. Moreover, Leishmania infection itself has been associated with an increased risk of miscarriage, particularly during active infection, possibly due to parasite‐induced placental inflammation and immunopathology [24].
Screening for Leishmania spp. infection in blood donors presents significant challenges, particularly in low‐endemicity settings. In‐house assays, such as ELISA based on alkaline extract antigen from L. infantum chagasi (EAL), are frequently used in surveillance studies due to their low cost and reasonable sensitivity [25]. However, the lack of standardization and potential cross‐reactivity with other endemic infections or physiological conditions (e.g., pregnancy) limit their clinical and transfusional applicability [24]. Commercial assays employing recombinant antigens (e.g., rK39 and rK28) offer improved specificity and quality control and are more suitable for systematic screening in blood banks [25]. Nevertheless, their sensitivity may vary depending on the circulating Leishmania species, and their higher cost poses a barrier to routine implementation. In the absence of an ideal serological test for blood donor screening, positive results should be interpreted with caution. The use of molecular assays such as PCR as confirmatory tools is recommended [26], along with the adoption of preventive strategies such as universal leukoreduction [27].
The absence of PCR positivity despite ELISA reactivity may reflect low parasite burdens in asymptomatic individuals, residual antibodies from past exposure or non‐specific antibody reactivity in inflammatory conditions. The real‐time PCR assay used in this study had a validated analytical sensitivity of 17.9 copies/mL [26, 27], which is adequate for detecting active infections. Therefore, the lack of detectable Leishmania DNA supports the hypothesis that the ELISA‐reactive results do not reflect current parasitaemia.
Sociodemographic analysis revealed higher seroprevalence among young individuals, self‐declared White and with higher educational attainment. This profile differs from that observed in traditionally endemic areas, where leishmaniasis is more common among socioeconomically vulnerable populations. However, the urbanization and expansion of the disease into higher‐income areas, along with better access to health services and routine screening, may explain this pattern in non‐endemic settings [28].
Interestingly, spatial distribution analysis of seroreactive donors revealed a higher prevalence in the Northern region of the state of São Paulo, followed by the Eastern and Western regions. These geographic differences may reflect a combination of ecological, environmental and urban structural factors that influence vector density and human exposure. The Northern region, in particular, is characterized by residual forest fragments, peri‐urban settlements and a higher frequency of canine leishmaniasis, which could contribute to sustained low‐level transmission. Similar patterns were reported by Silva et al. [29], who documented a progressive geographic spread of VL in both human and canine populations in Northern municipalities of São Paulo state. These findings suggest that despite being considered non‐endemic, certain microregions may harbour enzootic transmission cycles, highlighting the need for targeted surveillance and public health interventions [28, 29].
These findings underscore the importance of confirmatory testing, especially in blood donor screening programs in non‐endemic regions. The integration of molecular methods as complementary tools could help reduce unnecessary donor deferrals and enhance transfusion safety when Leishmania spp. screen is applied.
Finally, given the potential for subclinical infections, the influence of inflammatory physiological states such as pregnancy and abortion on serological testing, and the limitations of serology in low‐prevalence populations, universal leukoreduction emerges as a feasible and effective measure to reduce the risk of Leishmania transmission through transfusion. In parallel, continuous epidemiological surveillance is essential to monitor the potential introduction or re‐emergence of Leishmania in urban, non‐endemic areas, especially in light of migration, environmental changes and vector expansion.
In conclusion, in this study, a seroprevalence of 2.74% for Leishmania spp. was observed among blood donors from the metropolitan region of São Paulo, an area considered non‐endemic for the disease. However, the absence of PCR positivity in ELISA‐reactive samples suggests that these results do not reflect active infection but possibly residual antibodies or non‐specific reactivity, particularly among pregnant women and those with a history of abortion. These findings highlight the limitations of serological tests in low‐prevalence populations and reinforce the need for the use of confirmatory molecular methods in transfusion screening protocols.
Furthermore, the adoption of complementary strategies, such as universal leukoreduction, is essential to mitigate the risk of transfusion‐transmitted infection, especially in contexts where direct parasite detection is limited. Continuous epidemiological surveillance remains crucial to monitor emerging cases and guide public health policies in response to environmental changes, population mobility and vector expansion into urban areas. These data contribute to the improvement of transfusion safety programs in low‐endemicity regions and emphasize the importance of more specific diagnostic strategies for Leishmania spp. in blood banks.
CONFLICT OF INTEREST STATEMENT
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Silva PL , Montenegro S , Werkauser R , Sales KGS , Soares FCS , Costa VMA , et al. Asymptomatic Leishmania infection in blood donors from a major blood bank in Northeastern Brazil: a cross‐sectional study. Rev Inst Med Trop Sao Paulo. 2020;62:e 92.33263698 10.1590/S 1678-9946202062092 PMC 7694538 · doi ↗ · pubmed ↗
- 2Mohammed R , Melkamu R , Pareyn M , Abdellati S , Bogale T , Engidaw A , et al. Detection of asymptomatic Leishmania infection in blood donors at two blood banks in Ethiopia. P Lo S Negl Trop Dis. 2023;17:e 0011142.36893183 10.1371/journal.pntd.0011142 PMC 10030029 · doi ↗ · pubmed ↗
- 3Lima LVR , Pereira SRF , dos Santos SO , Silva ES , Oliveira‐Silva F , Moreira C , et al. Detection of Leishmania DNA in blood donors from Teresina, Brazil. Rev Inst Med Trop Sao Paulo. 2011;53:1–5.
- 4Shahbazi F , Bandehpour M , Mohebali M , Eslami G , Kazemi B , Haghighi A , et al. Detection of Leishmania infantum DNA by PCR in healthy blood donors in Iran. Transfus Apher Sci. 2008;38:153–157.
- 5Dantas‐Torres F , Miró G , Baneth G , Bourdeau P , Breitschwerdt E , Capelli G , et al. Canine leishmaniasis control in the context of one health. Emerg Infect Dis. 2019;25:1–4.10.3201/eid 2512.190164 PMC 687427731742505 · doi ↗ · pubmed ↗
- 6Junior MTS , Matsumoto T , Pimenta MFS , Shimozako HJ , Luz ZM . Investigação de Leishmania spp. em doadores de sangue de um hemocentro regional do Estado do Paraná, Brasil. Rev Soc Bras Med Trop. 2010;43:131–136.
- 7Aliaga L , Ceballos J , Antonio Sampedro A , Cobo F , López‐Nevot MA , Merino‐Espinosa G , et al. Asymptomatic Leishmania infection in blood donors from the southern of Spain. Infection. 2019;47:739–747.30888587 10.1007/s 15010-019-01297-3 · doi ↗ · pubmed ↗
- 8Druzian AF , Gama MEA , Nunes WS , Souza GS , Ciaravolo RM , Lima LV , et al. Detection of asymptomatic infection with Leishmania infantum by PCR in blood donors from Araçatuba, São Paulo, Brazil. Rev Soc Bras Med Trop. 2010;43:116–117.20464137